美拉德反应产物对动物免疫机制的影响
2022-11-14何颖
何颖
摘 要:美拉德反应产物会影响动物的生理过程,早期和晚期糖基化终产物都会增强动物机体的氧化应激以及炎症反应,日粮晚期糖基化终产物(advanced glycation end-products,AGEs)具有多种致敏性和免疫原性。动物摄入过量的AGEs或者在炎症状态下AGEs在动物体内大量表达将上调晚期糖基化终产物受體的表达和活化,而晚期糖基化终产物受体被激活后会产生活性氧,并引发炎性应答反应。如果持续刺激,则可能会引发慢性炎症疾病。
关键词:美拉德反应;晚期糖基化终产物;受体;免疫系统
中图分类号:S8816.9 文献标志码:A 文章编号:1001-0769(2022)05-0007-04
1 晚期糖基化终产物对免疫系统的影响
一般而言,晚期糖基化终产物(advanced glycation end-products,AGEs)会在三个层面上影响动物的生理过程,首先是改变信号转导途径,这发生在AGEs与受体互作之后;其次是通过改变信号途径,AGEs诱导或抑制某些细胞因子、激素和自由基的产生;最后由于AGEs的影响和促氧化活性的提高,靶组织中的蛋白发生变性,导致功能性失调。
大量研究表明美拉德反应产物(Maillard reaction products,MRPs)与免疫刺激作用和免疫系统有关。这种互作始于模式识别受体(pattern recognition receptors,PRRs)对MRPs构象的抗原表位识别,随后经下游信号通路传递到细胞核,主要是k基因结合核因子(nuclear factor-k-gene binding,NF-kB)的激活及随之而来的细胞应答。多种PRRs对MRPs具有潜在的识别能力和结合亲和力,但是它们之间的互作可能会引发各种应答反应。早期和晚期糖基化终产物都与氧化应激以及炎症反应的增强有关。AGEs和晚期糖基化终产物受体(receptor for advanced glycation end-products,RAGEs)互作会提高促炎性细胞因子如肠上皮细胞和单核细胞中的肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素-1β(Interleukin-1β,IL-1β)和IL-6的分泌量。自由基的增多和氧化应激的增强会加剧(自动放大)炎症状态,最终可能会影响胶原蛋白和弹性蛋白等长寿蛋白的性能。
被吸收入肠道并抵达黏膜组织的MRPs和AGEs与局部黏膜免疫系统进行对抗反应。这种接触会诱发免疫激活作用,并会影响局部的肠道组织,包括引起组织损伤和炎症反应,激活免疫反应,刺激T淋巴细胞和抗体(IgA)的产生。除了肠道免疫系统的局部活化,活化的免疫细胞还能够到达肠系膜淋巴结,并通过门静脉进入肝脏和全身循环,引发全身性反应。全身性激活免疫反应的一个例子就是过敏。就日粮过敏而言,日粮AGEs具有多种致敏性和免疫原性。启动和推进这些反应的机制仍存在争议。然而,有研究显示该机制取决于糖基化蛋白质的结构和类型,这些分子可能会促进或减弱过敏反应。据报道,随着IgE反应性的增强,烤花生的过敏原性增强。此外,Hilmenyuk等证实,成熟的树突状细胞(dendritic cells,DCs)会加载AGEs修饰态蛋白,增强T辅助细胞2(T helper 2,Th2)的应答反应,促进过敏反应。加热导致蛋白质之间发生交互联结,使它们能够抵抗蛋白质水解作用,同时也可能会增加这些产物的过敏原性。与天然形式的β-乳球蛋白相比,糖基化β-乳球蛋白的致敏性提高就证实了这一点,这可能是由于产生了新的过敏原。相反,加热也可能会使蛋白质变性,进而破坏或掩盖蛋白质构象的抗原表位,导致过敏原性降低。
2 AGEs的细胞受体
在抗原递呈细胞(antigen presenting cells,APCs)上表达的一系列细胞表面受体具有与AGEs结合和互作的亲和力。根据AGEs的糖蛋白结构,这些受体大多含有碳水化合物识别域(carbohydrate recognition domains,CRDs),或者与多肽互作的结构域。具有这些特征的细胞受体种类繁多,但并不 是所有的受体都能结合AGEs。此外,AGEs受体不以类似的亲和力与修饰态蛋白结合,这可能会引发不同的应答反应。综合与AGEs相关的研究,有6种与膜相关的受体被认为非常有价值。
3 RAGEs
RAGEs——晚期糖基化终产物受体,是一种多配体受体,属于免疫球蛋白超家族成员之一。该受体在不同类型的细胞上表达,包括单核细胞/巨噬细胞、内皮细胞和树突状细胞。它们能够识别大量分子,例如AGEs和淀粉样β多肽,并参与不同细胞的活化、迁移和成熟。摄入过量的AGEs或者在炎症状态下AGEs大量表达将上调RAGEs的表达和活化。RAGEs激活后会产生活性氧(reactive oxygen species,ROS),并引发炎性应答反应;如果持续刺激,则可能会引发慢性炎症疾病。RAGEs蛋白有3种变异体,分别为完整的RAGEs、N-端被删除的RAGEs和可溶性晚期糖基化终产物(soluble receptor for advanced glycation end products,sRAGEs),它们都拥有相同的核心功能域。sRAGEs的分子量为46~50 kDa,分泌至细胞外,含有细胞外配体结合域,通过负反馈机制调控RAGEs的水平。
4 半乳糖凝集素-3
AGE-受体3(AGE-receptor 3,AGE-R3)/半乳糖凝集素-3是外源凝集素家族的成员之一,与其他两个分子(AGE/R1/OST-48和AGEs-R/80K-H)一起形成AGEs-R复合物。该复合物主要位于细胞外分子量约32 kDa的半乳糖凝集素-3亚基,对巨噬细胞上的AGE-牛血清白蛋白(bovine serum albumin,BSA)复合物具有高度的亲和力。这种多功能受体通过自身的碳水化合物识别域和N末端结构域与糖蛋白互作,是急性和慢性炎症等生理过程的主要调控因子。鉴于半乳糖凝集素-3和糖蛋白(如AGEs)互作的能力,其在动物机体的免疫应答中也发挥重要作用。
5 Ⅰ型A类清道夫受体
(单体)Ⅰ型A类清道夫受体(scavenger receptor class AⅠ,SR-AⅠ)的分子量约为77 kDa,属于巨噬细胞清道夫受体(macrophage scavenger receptors,MSR)家族成员,由6个不同的結构域组成。同该家族的其他受体一样,SR-AⅠ主要参与调控微生物的吞噬作用。这些膜结合模式识别受体通常在单核细胞、巨噬细胞和树突状细胞上表达,具有多种功能和广泛的配体。除了结合微生物配体外,它们还能结合糖基化蛋白(例如AGEs)等修饰态蛋白,且具有高度的亲和力,并能促进它们的内吞作用。它们是先天免疫反应中的一个重要的功能性分子,参与巨噬细胞极化和动脉粥样硬化等疾病的发生。
6 分化抗原簇36(cluster of differentiation,CD36)
CD36[SR-B簇Ⅲ型(SR-BⅢ)]是一种糖蛋白,属于清道夫受体家族,在巨噬细胞上表达。与该家族的其他成员相似,分子量为88 kDa的CD36拥有大量的配体,并与微生物配体以及修饰过的分子结合。这种多功能受体参与吞噬、抗原提呈和凋亡细胞清除等生理活动,有助于炎性应答。多项研究表明,作为一种AGEs结合受体,CD36能够促进细胞对糖基化分子的摄取。
7 树突状细胞C型凝集素
树突状细胞C型凝集素[dendritic cell(DC)-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing non-integrin,DC-SIGN;CD209]是Ⅱ型跨膜受体家族成员之一,在树突状细胞膜上表达,介导树突状细胞与T淋巴细胞的黏附过程。DC-SIGN在DCs上大量表达,作为C型凝集素家族成员之一,其含有一个C末端凝集素结构域。DC-SIGN与多种配体结合,并基于它们产生各种反应。例如,有研究表明,在DC-SIGN与含有甘露糖的分子互作之后,白细胞介素10(Interleukin 10,IL-10)、IL-12和IL-6的生成量增加,但是DC-SIGN与含有海藻糖的分子结合之后,只有IL-10的生成量上调。DC-SIGN与碳水化合物结合的潜在能力使其成为结合修饰态糖蛋白和特异性AGEs的候选分子。
8 I型跨膜甘露糖受体
I型跨膜甘露糖受体(type I transmembrane mannose receptor,MMR;CD206)是C型凝集素家族中的另一个成员,拥有8个碳水化合物识别域。MMR不能与半乳糖结合,优先结合甘露糖和海藻糖,与葡萄糖的亲和力较低。有研究表明MMR作为模式识别受体,通过树突状细胞促进对抗原的摄取和呈递。这种与碳水化合物结合的能力有助于树突状细胞进行抗原内在化,使甘露糖受体成为结合AGEs的一个候选分子。
9 抗原递呈细胞
作为获得性免疫系统的效应细胞,T淋巴细胞不能识别游离抗原。树突状细胞和B淋巴细胞能够内吞抗原,并通过其主要组织相容性复合物Ⅱ类分子(major histocompatibility complex class II,MHC Ⅱ)向T淋巴细胞呈递抗原多肽。这些细胞被称为专业抗原递呈细胞(antigen-presenting cells,APCs),因为展示内源性获得的肽是其功能的主要组成部分之一。树突状细胞和巨噬细胞(也称为单核血细胞)是在先天性免疫系统和获得性免疫系统之间建立联系的主要免疫细胞。抗原递呈细胞通常进行内吞,并处理复杂的抗原,随后通过不同途径在MHC结合凹槽上表达多肽。此后,根据抗原的类型和性质,T淋巴细胞开始增殖和分化,并产生各种形式的免疫应答。树突状细胞的主要作用是将抗原引入初始型T淋巴细胞(naive T cells),其中巨噬细胞和B淋巴细胞分别通过细胞介导和体液反应激活T淋巴细胞。抗原识别、呈递和活化的整个过程是免疫系统对不同干扰做出正确应答所必需的,而APCs在其中起着关键作用。
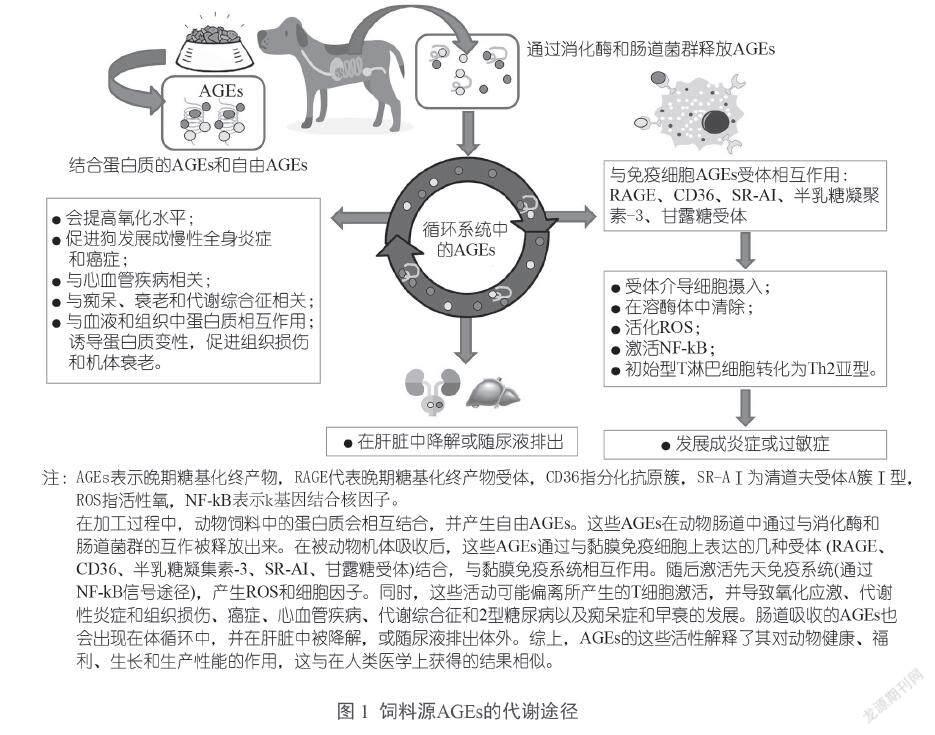
当AGEs与APCs上表达的受体互作时,这些AGEs会被APCs内吞,并通过MHC Ⅱ复合物分子呈递给特异性T淋巴细胞。在小鼠模型中,与未经过修饰的卵清蛋白相比,通过在髓样树突状细胞中表达的Ⅰ型和Ⅱ型A类清道夫受体(SR-AI/Ⅱ),AGEs修饰态卵清蛋白能够被更有效地吞噬。抗原呈递的增强导致卵清蛋白特异性CD4+辅助T淋巴细胞的活性增加。有研究发现,人类的树突状细胞增强了对用异硫氰酸荧光素标记的AGEs修饰态卵清蛋白的内吞作用,该过程是由AGEs结合甘露糖受体、清除受体以及这些细胞的大胞饮作用介导的。结果,由此产生的T细胞激活导致Th2细胞因子(IL-5、IL-4和IL-6)的水平显著提高;相比之下,非糖基化卵清蛋白负载的DCs导致Th1细胞因子(IFN-γ)或调节性T细胞细胞因子(IL-10)的水平提高。图1描述了饲料源AGEs影响新陈代谢的几个因素。
原题名:Immunomodulation by processed animal feed: the role of Maillard reaction products and advanced glycation end-products (AGEs)(英文)
原作者:Malgorzata Teodorowicz、Wouter H Hendriks、Harry J Wichers和Huub F J Savelkoul
