抗禽流感病毒(H5N1)入胞单分子抗体过表达慢病毒载体的构建及其稳转细胞株的建立
2022-11-11岳玉环吴广谋张国利蔄弘扬李泽鸿
王 瑜,孙 赫,岳玉环,吴广谋,张国利,邱 月,蔄弘扬,田 园*,李泽鸿
(1.吉林农业大学 生命科学学院,吉林 长春 130117;2.中国农业科学院 长春兽医研究所 生物技术应用研究室,吉林 长春 130122)
自1997年香港首次报道人感染H5N1亚型禽流感病毒(avian influenza virus,AIV)以来,高致病性H5N1 AIV在禽类之间持续流行,并不断发生人类感染事件,从而赋予了禽流感全新的公共卫生意义,即高致病性AIV对人类健康的巨大威胁不仅在于造成当前严重的个体感染甚至死亡,更在于病毒持续变异,可能获得人际传播能力,导致新一轮人类流感的大流行[1-2]。
H5N1 AIV最主要的中和抗体来自病毒表面糖蛋白血凝素(HA),所以HA成为以往主要研究靶标。然而H5N1 AIV具有高度变异性,根据分子进化树分析预测结果显示H5N1 AIV HA基因已演变成至少10种不同抗原性特征的变异分支,不同分支之间的免疫交叉反应较弱[3-4]。鉴于病毒变异的影响,基于保守抗原组分的抗体能够对不同亚型AIV产生抗病毒作用。流感病毒基质蛋白M1是AIV主要的结构蛋白,位于病毒囊膜内侧,通过与宿主细胞靶蛋白结合而参与和调控病毒的复制、转录、释放等过程。M1蛋白序列保守,因此针对M1的抗体可以通过与M1蛋白结合,抑制其活性,干扰各亚型AIV的复制、转录及释放,从而起到广谱抗病毒的作用[5-6]。本研究前期利用H5N1的M1蛋白作为抗原从人源噬菌体库抗体中筛选获得了特异性抗M1的ScFv。本研究将艾滋病病毒TAT蛋白核心转导域(PTD)与ScFv及突变的人源Fc融合表达,构建并表达可入胞且稳定性较好的TAT-ScFv-mFc融合抗体。目前关于入胞单分子抗体(ScFv-mFc)在真核细胞表达系统的研究尚未见报道,本研究的目的在于构建稳定表达抗H5N1入胞单分子抗体HEK293T细胞系[7],实现单分子抗体在哺乳动物细胞的表达,为研制抗体药物奠定基础。
1 材料与方法
1.1 主要材料HEK293T细胞系、抗H5N1 AIV M1蛋白的单链抗体3F、ScFv-mFc及TAT-ScFv-mFc原核表达载体由中国农业科学院长春兽医研究所生物工程应用与基因工程药物实验室构建并保存;psPAX2、pMD2.0G及大肠杆菌DH5α感受态细胞购自艾柏森生物科技有限公司;嘌呤霉素(Puro)、BSTBⅠ、MluⅠ、BCA Protein Assay Kit、West Dura Extended Duration Subatrate以及DMEM购自美国Thermo Fisher;pLVX-mCMV-ZsGreen1-Puro(pLVX)质粒及重组酶mix购自通用生物系统(安徽)有限公司;限制性核酸内切酶及T4DNA连接酶购自安诺伦(北京)生物科技有限公司;胎牛血清购自以色列BI公司。
1.2 重组慢病毒载体的构建及重组质粒酶切鉴定以人工合成3F(H5N1)-FC单分子抗体基因序列为模板,PCR扩增目的基因,上游引物5′-agatctcgagctcaagcttcgaa-3′,下游引物5′-tgcgcataaacgccgg-3′,上、下游引物中分别引入酶切位点BstBⅠ、MluⅠ。扩增体系为50 μL:2×KOD PCR Master mix 25 μL,上、下游引物各1 μL,重组质粒1 μL,双蒸水22 μL。反应条件:94℃ 2 min;98℃ 10 s,58℃ 10 s,68℃ 1 min,共30个循环;68℃ 5 min。PCR扩增后的目的片段经1%琼脂糖凝胶电泳,并利用DNA纯化试剂盒进行胶回收。
将扩增并回收的入胞单分子抗体基因序列与pLVX质粒载体采用限制性内切酶BstBⅠ、MluⅠ进行双酶切。酶切体系为Fast Digest BstBⅠ1 μL、Fast Digest MluⅠ 1 μL、10×buffer 2 μL、载体 1 μg、加双蒸水至20 μL,16℃过夜。将酶切产物经1%琼脂糖凝胶电泳后回收。将切胶回收的目的片段与载体使用T4DNA连接酶连接,50℃反应15~20 min。
1.3 重组载体验证将连接体系转化DH5α感受态细胞。从-80℃冰箱中取1管感受态细胞,立即置于冰上;加入连接体系(体积不超过10 μL),轻轻混匀,冰上放置30 min;42℃水浴中准确热击90 s,热击后迅速置于冰上冷却3~5 min;向管中加入1 mL LB液体培养基(不含抗生素),混匀后于37℃、200 r/min摇床振荡培养1 h,使细菌恢复正常生长状态;将上述菌液离心,去除800 μL上清;剩余培养基吸打混匀,取150 μL菌液均匀涂布于含卡那霉素的培养皿中,37℃培养16~24 h;转化后的平板挑菌,将克隆菌液送测序公司进行测序。
1.4 慢病毒包装将细胞状态较好的HEK293T细胞接种到10 cm细胞培养皿中用于转染。准备所需质粒并计算使用量,其中病毒载体∶psPAX2∶pMD2.0G=3∶2∶1(质粒总量20 μg)。将DMEM、质粒、转染试剂配制成质粒-转染试剂复合物(DMEM∶质粒∶PEI = 100 μL∶1 μg∶2 μg),吹打混匀后室温静置15 min,然后将其滴加到待转染的细胞中,转染6 h后换成新鲜的含10% FBS的DMEM培养基。在细胞培养48 h后将含病毒的细胞培养液收集到15 mL离心管中,然后再向细胞培养皿中加入新鲜的含10% FBS DMEM培养基,继续培养24 h后可进行第2次病毒收集。收集的病毒分装后,-80℃保存备用。
1.5 慢病毒侵染HEK293T细胞将细胞状态良好的HEK293T细胞接种到6孔板中,使其在第2天侵染时的密度能达到60%;弃去6孔板中的培养基,加入1.5 mL完全培养基和0.5 mL收集的病毒上清。侵染24 h后换成新的培养基继续培养,并在荧光显微镜下拍照观察;第2天弃去培养基,加入1.5 mL完全培养基和0.5 mL收集的病毒上清继续第2次侵染。侵染24 h后换成新的培养基继续培养,细胞长满时可进行正常的传代培养,并在荧光显微镜下拍照观察侵染后效果;准备已经侵染过的HEK293T细胞以及未侵染的正常HEK293T细胞,待密度达到60%时,在培养基中加入Puro(4 mg/L)培养细胞,待细胞长满时进行细胞传代。培养过程中全程使用Puro培养液以去除转染失败的HEK293T细胞。
1.6 稳定表达的HEK293T细胞系的单克隆筛选将HEK293T细胞稳转细胞株计数,之后梯度稀释成20 cell/mL的细胞悬液,以100 μL/孔均匀加入96孔板;待细胞贴壁后观察每个孔的细胞数量,发现每孔细胞数量在0~4个之间,对只有1个细胞的孔进行标记,待培养基开始变黄时更换新鲜培养基。期间对单克隆细胞在相同曝光强度下进行拍照观察,挑选荧光较强的单克隆细胞扩大培养,并依次转移至24孔板、6 cm细胞培养皿中培养,全程使用含Puro(2 mg/L)的培养基维持培养。
1.7 Western blot检测转染后目的蛋白的表达使用PBS将转染后的293T细胞洗涤2次,离心并收集细胞,加入冰预冷的裂解液并置冰上裂解30 min,使细胞充分裂解并于4℃、12 000 r/min离心5 min,收集上清。将上清煮沸变性5 min后,加等量蛋白上样。蛋白质在10% SDS-PAGE凝胶中分离并转膜。5% BSA-PBS室温封闭2 h后,加入带有HRP标记的抗Fc单克隆抗体4℃过夜。TBST洗膜5×5 min,ECL法检测。
1.8 TAT-ScFv-mFc的纯化制备大量的293T细胞,取细胞上清,4℃、10 000 r/min离心15 min后,用0.22 μm滤膜过滤。使用ProteinG 柱亲和层析柱进行蛋白纯化:用5倍柱床体积的去离子水洗涤,去除20%乙醇;5倍柱体积Buffer A平衡柱子(Buffer A:PBS,pH 7.4);将样品以3 mL/min的速度上ProteinG柱;用Buffer A洗流川;用0.1 mol/L甘氨酸(pH 2.7)洗脱目的蛋白,2 mol/L Tris buffer中和至中性。收集0.1 mol/L甘氨酸洗脱样品,于4℃透析过夜,透析外液:PBS(pH 7.4).取透析后的样品,用超滤法(超滤管)浓缩,SDS-PAGE检测目的蛋白。
1.9 ELISA检测方法的建立使用M1蛋白(0.8 g/L)包被酶标板(100 μL/孔),37℃静置1 h;PBST洗板后加入5%的脱脂奶粉封闭1 h;PBST洗板后加入TAT-ScFv-mFc(0.39 g/L)于反应板中(100 μL/孔),37℃静置1 h;PBS洗板;加入HRP标记的Protein A(0.5 mg/L)作为二抗(100 μL/孔),37℃静置1 h;洗涤后加入TMB显色15 min,最后以50 μL/孔加入终止液(H2SO4:0.5 mol/L),酶标仪读取D450值。
2 结果
2.1 pLVX-TAT-ScFv-mFc-ZsGreen1-Puro表达载体的构建将目的片段与pLVX质粒采用限制性内切酶BstBⅠ、MluⅠ进行双酶切,经1%琼脂糖凝胶电泳后回收。连接转化后(图1),随机选取1条表达载体进行PCR扩增验证目的片段(图2)。结果表明,目的基因片段在1 500 bp条带清晰,说明pLVX-TAT-ScFv-mFc-ZsGreen1-Puro表达载体构建成功。

图1 pLVX-TAT-ScFv-mFc-ZsGreen1-Puro载体图谱
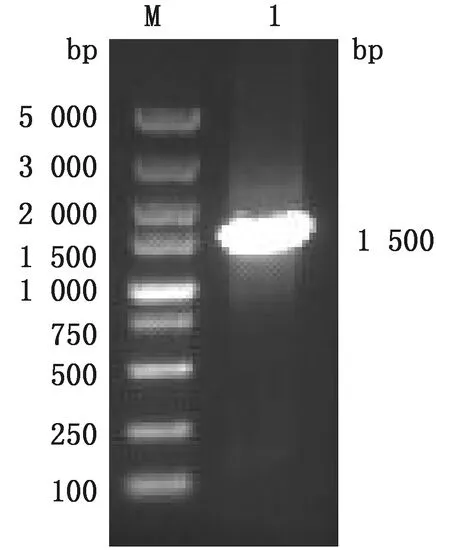
M.DL5000 DNA Marker;1.pLVX-TAT-ScFv-mFc-ZsGreen1-Puro
2.2 稳转细胞株的构建重组质粒瞬时转染状态良好的293T细胞(图3),荧光显微镜观察绿色荧光蛋白,结果显示,在48 h后慢病毒侵染HEK293T细胞成功。继续使用Puro培养侵染后的细胞,发现正常HEK293T细胞在Puro处理后全部死亡,而被病毒侵染过的HEK293T细胞状态良好。通过单克隆筛选出荧光较强的H5N1-HEK293T细胞,分别命名为H5N1-HEK293T-C3和H5N1-HEK293T-D4(图4)。

A.慢病毒侵染前HEK293T细胞;B.慢病毒第1次侵染24 h-HEK293T;C.慢病毒第2次侵染48 h-HEK293T
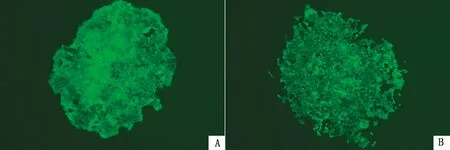
A.H5N1-HEK293T-C3;B.H5N1-HEK293T-D4
2.3 转染后目的蛋白表达检测结果Western blot检测转染后目的蛋白的表达,证实H5N1-HEK293T-C3和H5N1-HEK293T-D4均在55 kDa有明显条带(图5),与预期相符,表明稳转细胞株构建成功,扩大培养后冻存。

1.HEK293T;2.H5N1-HEK293T;3.H5N1-HEK293T-C3;4.H5N1-HEK293T-D4
2.4 TAT-ScFv-mFc的纯化结果细胞裂解液经ProteinG 柱亲和层析柱纯化,以截留分子量为10 kDa 超滤管浓缩至1 mL,采用BCA方法测定纯化蛋白质量浓度为0.39 g/L,初步满足试验目的要求。SDS-PAGE电泳结果显示55 kDa处有目的条带(图6),符合试验预期结果。
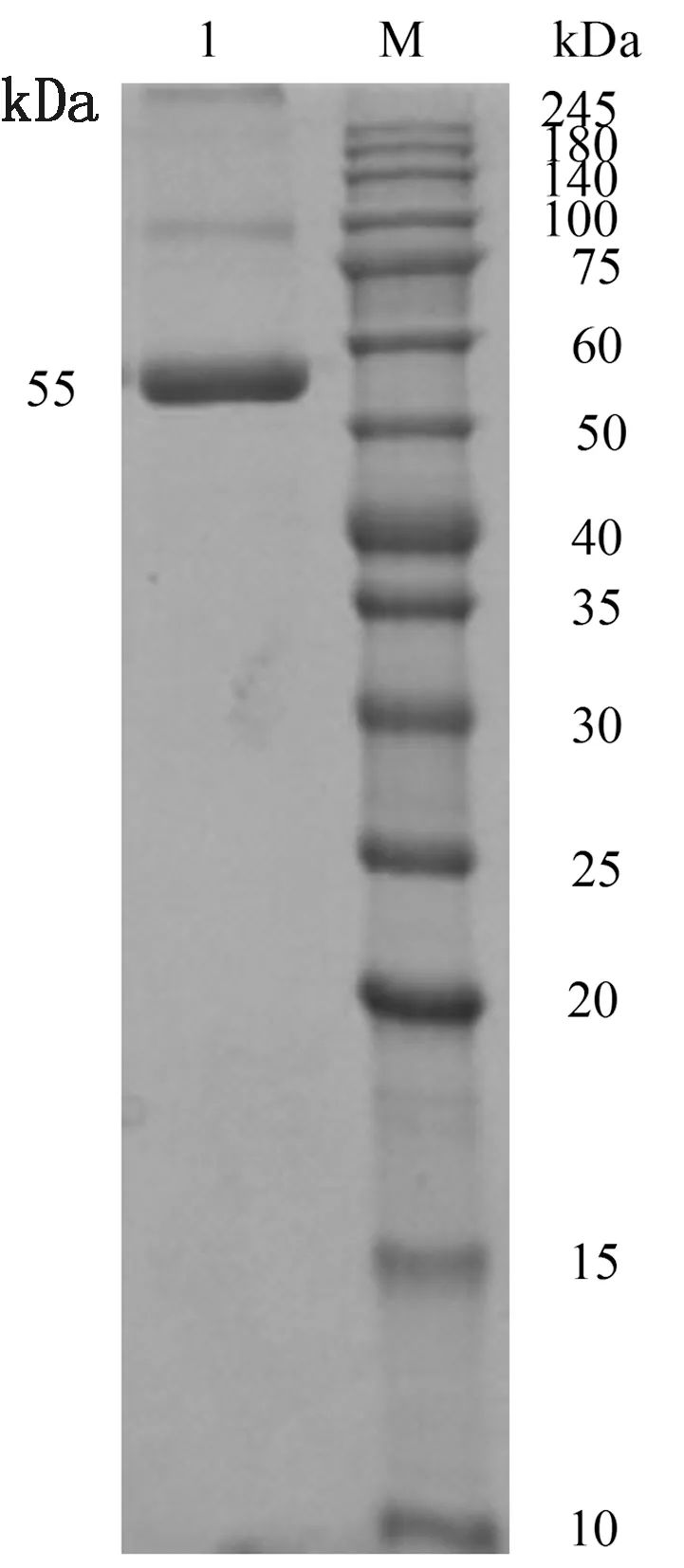
图6 TAT-ScFv-mFc的SDS-PAGE结果
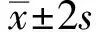
3 讨论
野生鸟类是AIV的天然宿主,病毒可以从野鸟传播给家禽或包括人类在内的哺乳动物。AIV在鸟类中的高流行率、形成新的高致病性重配体的可能性以及这些病毒传播给人类的能力使AIV成为全球高度关注的问题[8]。
针对人禽流感的治疗目前尚无确切的治疗手段,以使用抗病毒药物为主,当前批准的抗病毒药物包括2种离子通道抑制剂和2种神经氨酸酶抑制剂,但在流感病毒不断产生耐药的情况下,制备新型抗流感病毒药物非常必要[3]。抗体对于严重流感的治疗非常有效,但其免疫原性强,临床应用易引发人体变态反应。随着基因工程抗体技术的发展,单分子抗体以其特异性高、相对分子质量小,结构简单,较亲本抗体免疫原性低,在临床应用中能够最大减轻异种蛋白引起的变态反应的独特优势,吸引了众多科研工作者的目光,其制备技术已趋于成熟,特别是噬菌体展示技术更提高了抗体及抗体基因的筛选效率。单分子抗体对病毒感染性疾病的治疗将发挥重要的作用[9]。
本研究选择人高致病性AIV H5N1保守序列M1蛋白为靶标抗原,利用噬菌体抗体库筛选出人源抗M1蛋白的高亲和力单链抗体;为实现抗体分子进入细胞,将TAT蛋白PTD与抗M1蛋白单链抗体基因连接,构建并表达TAT-ScFv。为增加其体内稳定性,将TAT-ScFv与铰链区突变的人IgG的Fc片段融合表达,其中铰链区突变Fc为CPPCP中半胱氨酸突变为APPGP,意在防止分子内和分子间形成二硫键,从而制备出抗人AIV的全人源单分子入胞抗体[10]。
现有的基于M1蛋白的H5N1单分子抗体均为原核表达,DONG-DIN-ON等[11]已成功构建了基于原核表达抗H5N1 M1蛋白的单分子抗体。虽然大肠杆菌E.coli表达系统是常用、有效和方便的表达系统,可以进行许多异源蛋白的高效表达,但原核细胞系统表达蛋白时,存在多种缺陷:真核基因通常含有内含子原核细胞不能对其进行正确的剪切和拼接;缺乏真核生物的蛋白翻译后修饰和加工;表达产物多以包涵体形式存在,复性困难且效率很低;背景杂蛋白多、不易于纯化等,因此不可避免地对利用原核细胞表达基因工程抗体造成一定影响[12]。PIPATTANABOON等[13]将单链可变片段(ScFv)与Fc区结合,产生了在哺乳动物HEK293T细胞中表达的基于血凝素(H5透明质酸)ScFv-Fc单克隆抗体,产率为12.87和33.56 mg/L。其优点有以下几个方面:真核细胞在外源蛋白表达上具有原核细菌系统不可比拟的优势,因其在表达时对蛋白的修饰加工、有效分泌等特性而备受临床科研重视,真核细胞表达系统势必为今后基因工程抗体的高效表达和商业化生产开辟新的空间[14-15]。
本研究在体外构建了抗H5N1 M1蛋白入胞单分子抗体基因的过表达慢病毒载体,并成功插入到HEK293T细胞中。通过Puro和单克隆筛选成功建立了一个稳定表达TAT-ScFv-mFc的HEK293T细胞克隆。将该细胞克隆扩增培养后Western blot检测到TAT-ScFv-mFc在HEK293T细胞上清中的高效表达,且成功获得2 mL(0.39 g/L)的M1蛋白抗体,为进一步研究TAT-ScFv-mFc的功能及在真核表达系统的制备奠定了基础。
