抚仙湖北部沉水植被演变规律及其生态指示意义(1987-2020年)*
2022-11-09朱可欣
朱可欣,王 荣*
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,南京 210008) (2:中国科学院大学,北京 100049)
湖泊是地球系统重要组成部分,在涵养水源、净化环境、调节气候、维持生物多样性等方面都发挥着重要作用. 20世纪,湖泊生态系统受气候变化和人类活动的影响,生态环境急剧恶化[1]. 全球气候变暖、极端气候事件增加以及城市化进程加快、工农业发展等已经导致许多湖泊富营养化不断加剧,造成浮游藻类暴发、沉水植物消亡,伴随出现生物多样性下降和生态系统功能丧失等生态环境问题[2].
沉水植被是湖泊生态系统的重要组成部分,为其它水生生物提供栖息地、维持生物多样性、抑制浮游植物生物量[3]. 人类活动和气候变化通过光照、营养盐、透明度、温度等[4]影响湖泊沉水植物的生长,从而影响整个湖泊生态系统功能的发挥. 沉水植物的逐步消亡会打破湖泊水体的生态平衡,使得水体由水生植物占优势(草型湖)转变为浮游植物占优势的状态(藻型湖)[5],湖泊生态系统发生稳态转换,生态系统功能明显丧失. 认识沉水植物的演变规律是了解湖泊生态系统现状和演变趋势预测的关键.
我国湖泊监测时段较短,普遍在湖泊生态环境风险出现后才逐渐开展. 传统野外调查覆盖范围较小,费时费力. 遥感技术因其探测范围大、时间连续、精度高等特点已经成为湖泊生态环境调查主要辅助手段之一[6-7]. 基于遥感技术的植被提取方法也已逐步完善,研究成果已经广泛应用于湖泊生态系统对全球变化响应的研究中. 李静泰等[8]利用长时间序列Landsat 8影像分析了太湖水生植被时空变化特征并进行驱动机制研究,认为风向对水生植被的迁移有明显影响; 梁莉莉等[9]通过遥感数据进行植被识别,分析了东平湖菹草时空格局变化. 遥感影像本身存在“异物同谱”、“同物异谱”等问题,仅依靠原始光谱难以消除其造成的误差,目前的研究发现除利用原始遥感数据直接提取沉水植被信息外,还能借助指数计算、图像变换等手段构建多个变量以增强地物光谱信息,利用多变量决策树方法进行水生植物的遥感信息提取. 张莹莹等[10]基于决策树分类方法,集成遥感影像光谱特征、植被指数、水体指数、纹理特征等对洪湖水生植物遥感信息提取进行精度验证. 王琪等[11]构建太湖水生植物分类决策树模型识别沉水植物优势种. 常规决策树一般依据人为目视解译和经验构建,受人为主观影响较大,而分类回归树(classification and regression trees,CART)方法能通过算法自动选择地物分类特征,是决策树模型的典型代表[12].
抚仙湖是我国第二大深水湖泊,湖容积约210亿m3,蕴含丰富的淡水资源[13]. 抚仙湖流域属滇中高原,湖泊被群山环抱,流域生态系统封闭程度高. 近年来不断增强的工农业活动和区域气候变化,使得抚仙湖流域内出现山地生态系统退化、水污染加剧、水质下降和水生态结构受损等问题[14]. 围绕抚仙湖及流域已开展了湖泊水质变化规律[15-16]、农业污染来源判识[17]、植被群落结构演变[18]、生态修复[14]等工作,还缺乏关于沉水植被长期变化的相关研究.
抚仙湖主要污染源位于流域北部澄江县,以居民生活污水、工业污染、农业面源污染为主[17]. 中国科学院抚仙湖高原深水湖泊研究站调查报告表明,2019年5月抚仙湖北部水域出现狐尾藻大量增殖以及附着藻类暴发的情况,局部水质严重恶化. 对该区域沉水植物长期演化过程以及驱动机制的探讨将有利于认清抚仙湖生态系统演化特征以及为全湖管理提供科学建议. 本文利用Landsat遥感数据,基于CART算法构建分类决策树提取了抚仙湖北部湖区沉水植物面积,分析该区域沉水植被面积动态变化,结合人类活动、水体营养、气候水文数据探究驱动沉水植被演替的机制. 本文旨在通过对抚仙湖北部区域沉水植物的变化来了解抚仙湖水生生态系统对于环境变化的响应特征,为抚仙湖水质条件维持和生态环境改善提供科学依据.
1 材料与方法
1.1 研究区概况
抚仙湖(24°21′~24°38′N,102°49′~102°57′E,图1)地处云南省中部,北临澄江市,与星云湖通过隔河相连,是我国典型断陷深水湖泊,海拔1723.4 m,面积约216 km2,南北长31.5 km,东西最宽11.5 km,最大水深158 m,平均水深89 m,湖容积量约210亿m3,平均换水周期250年,流域面积较小,约为675 km2. 抚仙湖地处亚热带半湿润季风气候区,干湿季分明,光照充足,年平均气温15.5℃,全年无霜期近300 d,降水集中于5-10月[19]. 湖泊存在季节性分层,温跃层一般出现在5-11月,深度都在40 m以上[20]. 近年来,由于人口增加,工农业活动加剧,旅游业快速发展等导致抚仙湖整体水质呈现下降趋势[16]. 中国科学院抚仙湖高原深水湖泊研究站监测的2020年抚仙湖年均总氮(TN)、总磷(TP)浓度为0.318、0.032 mg/L(标准差为0.081、0.008 mg/L),湖泊综合营养指数具有增加趋势[21]. 抚仙湖沉水植物现处于高大冠层型植物占优势、外来物种快速发展和丝状附着藻增殖的发展阶段,群落结构不稳定[22]. 2019年5月抚仙湖北岸狐尾藻大量增殖的同时附着藻类暴发,已经严重影响了抚仙湖局部水质和生态环境. 依据监测采样点的分布,选定了抚仙湖北部76.4 km2的局部湖区作为研究范围(图1).
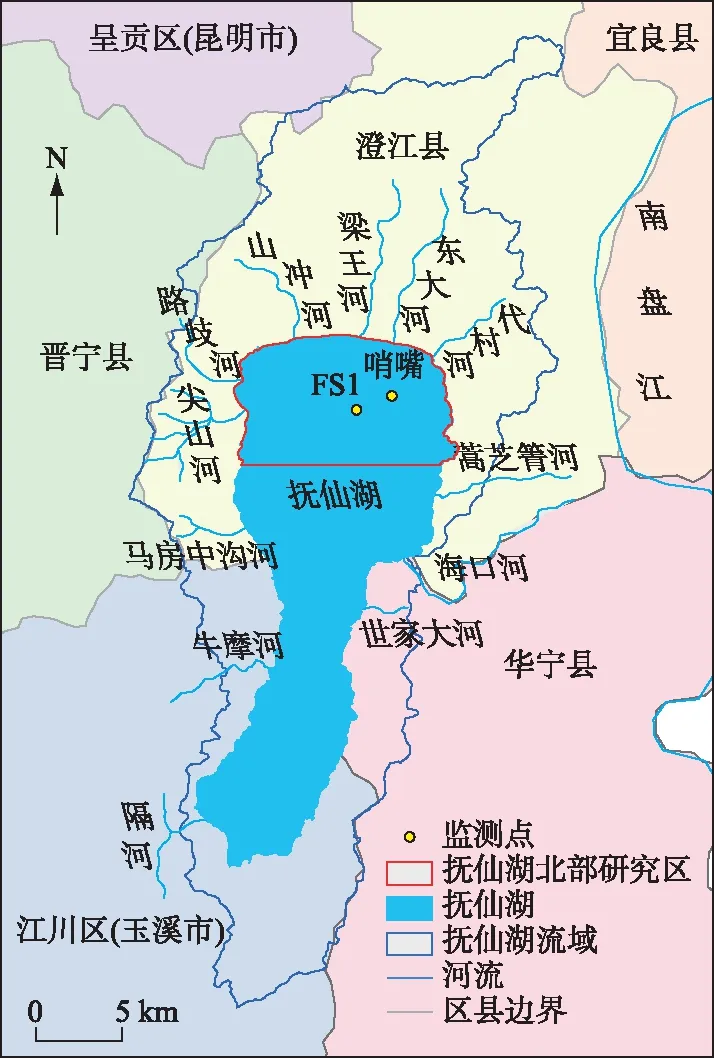
图1 抚仙湖研究区范围Fig.1 Selection range of Lake Fuxian research area
1.2 数据来源
遥感影像从中国区域Landsat地表反射率产品集获取[23],下载1987-2020年遥感反射率数据,其中2012、2013年由于卫星传感器更新和云层覆盖问题未获取到有用影像. 气温、降水、风速数据来源于国家气象科学数据中心(http://data.cma.cn). 以昆明、红塔、宜良三站的平均数据作为研究区气象资料. 湖泊水位、透明度、TP、TN浓度数据来源于文献[15-16,24-25],以哨嘴监测点数据指示北部湖区水体营养(图1),2006-2010年的营养数据存在缺失,2019、2020年水体营养数据由中国科学院抚仙湖高原深水湖泊研究站提供(监测点FS1,图1).
1.3 研究方法
1.3.1 植被面积提取 利用ENVI软件对影像进行波段合成、主成分变换、缨帽变换、波段计算等处理. 其中缨帽变换后的亮度指数与绿度指数的差值(TC1-TC2)能较好地识别沉水植被[26]. 主成分信息、缨帽变换和波段计算的结果作为特征变量合成基础影像,对每幅影像随机抽取地物分类训练样本,基于ENVI软件CART算法程序对训练样本进行分析,并自动生成分类决策树文件,以识别沉水植物分布范围.
1.3.2 结构方程模型 结构方程模型(structure equation modeling, SEM)是一种建立、估计和检验因果关系模型的方法,是验证性因子模型和因果模型的结合[27],不仅可以考察变量间的直接影响,还能揭示变量间的间接影响[28]. 本文利用R 4.05软件piecewise SEM软件包进行SEM分析,揭示年际抚仙湖北部湖区沉水植被面积、年均气温、年降水量、年均风速、年均TP浓度、年均TN浓度数据、年均水位、透明度等因子之间的关系. 因数据的缺失问题,将数据拆分为合适的统计数据组合分别进行SEM分析.
2 结果
2.1 气候变化
1987-2020年间,抚仙湖区域年降水量总体变化趋势不明显,略有减少(图2a),1999年降水量最多,为1385.2 mm,2009-2013年降水量整体偏低,仅有654.3、870.1、638.5、748.9和880.2 mm. 年均风速呈显著增加的趋势(图2b),1988-2002年平均风速不超过2.00 m/s, 2003年后,风速显著增加, 最大年平均风速2.47 m/s 出现在2020年. 年平均气温呈明显增加趋势(图2c),1987年抚仙湖区域平均气温为15.95℃,2020年增加到17.2℃,增温超过1.2℃,增速为0.38℃/10 a,最高值出现在2019年,年均气温达到17.6℃.
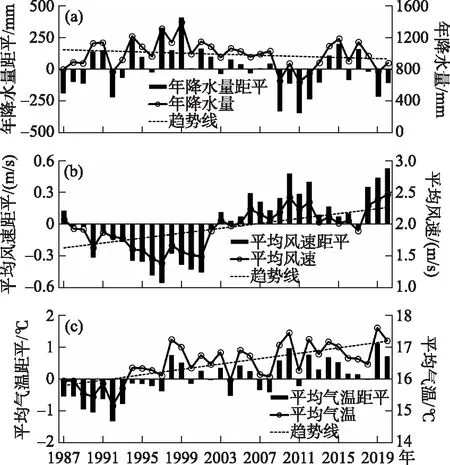
图2 抚仙湖区域气候年变化: (a)降水量;(b)风速;(c)气温Fig.2 Annual climate variation in the Lake Fuxian region: (a) precipitation, (b) mean wind speed, (c) air temperature
2.2 水体营养、水位、透明度变化
1987-2005年间,抚仙湖年平均TP浓度多数年份低于国家地表水I类水标准(图3a),1998年TP浓度0.017 mg/L为1987-2005年间的最高值,2011-2018年TP浓度保持在0.010 mg/L左右,2019、2020年TP浓度显著上升,分别为0.022和0.031 mg/L. 年平均TN浓度在1995、1996年明显偏高,分别达到0.51和0.37 mg/L(图3b). 2012年开始,TN浓度呈逐步上升趋势,2020年为近十年的最高值. 抚仙湖水位变化分为两个时期(图3c),2008年(2007年12月,星云湖抚仙湖出流改道工程建成通水[29])前,多年平均水位为1722.34 m,2008年后(出流改道后)多年平均水位为1721.62 m,水位下降0.73 m,2014年水位最低为1720.79 m (图3c). 抚仙湖水体透明度的变化存在先下降后略有回升的变化趋势,1987-1995年平均透明度为6.64 m,1996-2004年为5.07 m,2005-2020年为5.95 m(图3d).
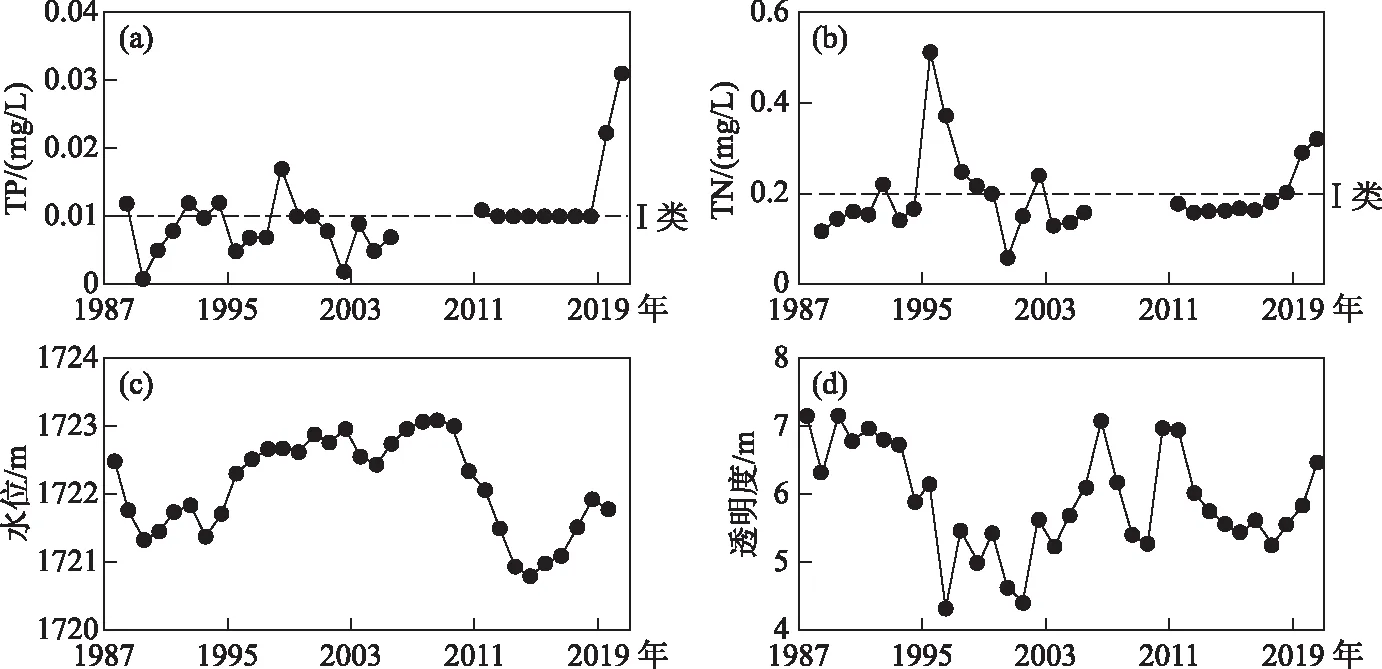
图3 抚仙湖区域水质水文年平均变化: (a)TP浓度;(b)TN浓度;(c)年际水位;(d)水体透明度Fig.3 Annual hydrological changes in water quality in Lake Fuxian region: (a) TP concentration, (b) TN concentration, (c) water level, (d) water clarity
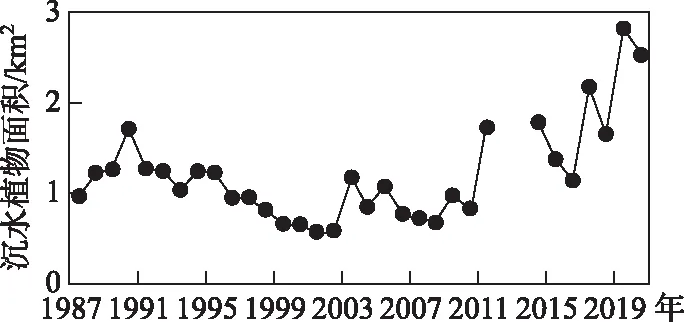
图4 抚仙湖北部湖区沉水植物面积变化 Fig.4 Changes in submerged vegetation area in the northern lake area of Lake Fuxian
2.3 抚仙湖北部湖区沉水植被面积变化
2005年对抚仙湖的实地调查表明,全湖沉水植被分布面积为3.19 km2[30],2016年沉水植物面积为5.14 km2[22],基于2次调查结果进行遥感解译沉水植被的精度验证. 遥感解译方法得到2005年全湖沉水植物的分布面积为3.34 km2,2016年为5.36 km2,误差均小于5%,证明本研究方法提取精度较高. 1987-2020年,抚仙湖北部湖区沉水植物分布面积存在先逐步减少后迅速增加的变化趋势(图4),呈现3个变化阶段. 第一阶段1987-1995年,沉水植物平均面积为1.25 km2(标准差:0.19 km2),第二阶段为1996-2010年,该阶段沉水植物面积最小,平均值为0.83 km2(标准差:0.17 km2),比第一阶段减少0.42 km2. 第三阶段2011-2020年,该阶段沉水植物分布面积明显增加,平均值达到1.91 km2(标准差:0.53 km2),比第二阶段增加1.08 km2,增幅为130.09%. 在研究时间段内,2001年沉水植物分布面积最小,只有0.58 km2,2019年分布面积最大,为2.83 km2,是2001年的4.88倍.
2.4 SEM路径分析
结构方程模型(SEM)分析结果显示水位(相关系数=-0.64)和总磷浓度(相关系数=0.59)是影响抚仙湖北部湖区沉水植被变化的主要直接因子,水位与沉水植被面积呈现负相关关系;总磷浓度与沉水植物面积有正相关关系. 总氮、透明度与沉水植物面积之间的关系没有通过显著性检验(图5). 气候要素中没有直接作用于沉水植物变化的因子,但气温通过影响总磷浓度间接影响沉水植物面积的变化,气温与总磷浓度呈正相关关系,而总磷与沉水植物呈正相关,因此气温的增加对沉水植物的面积扩张具有积极影响. 风速对水体透明度有积极影响,气温与水体透明度具有负相关关系,其他因子间关系没有通过显著性检验. 总氮浓度与总磷浓度间的关系错综复杂,难以用简单的路径指示,因此未列入结构方程分析结果中.
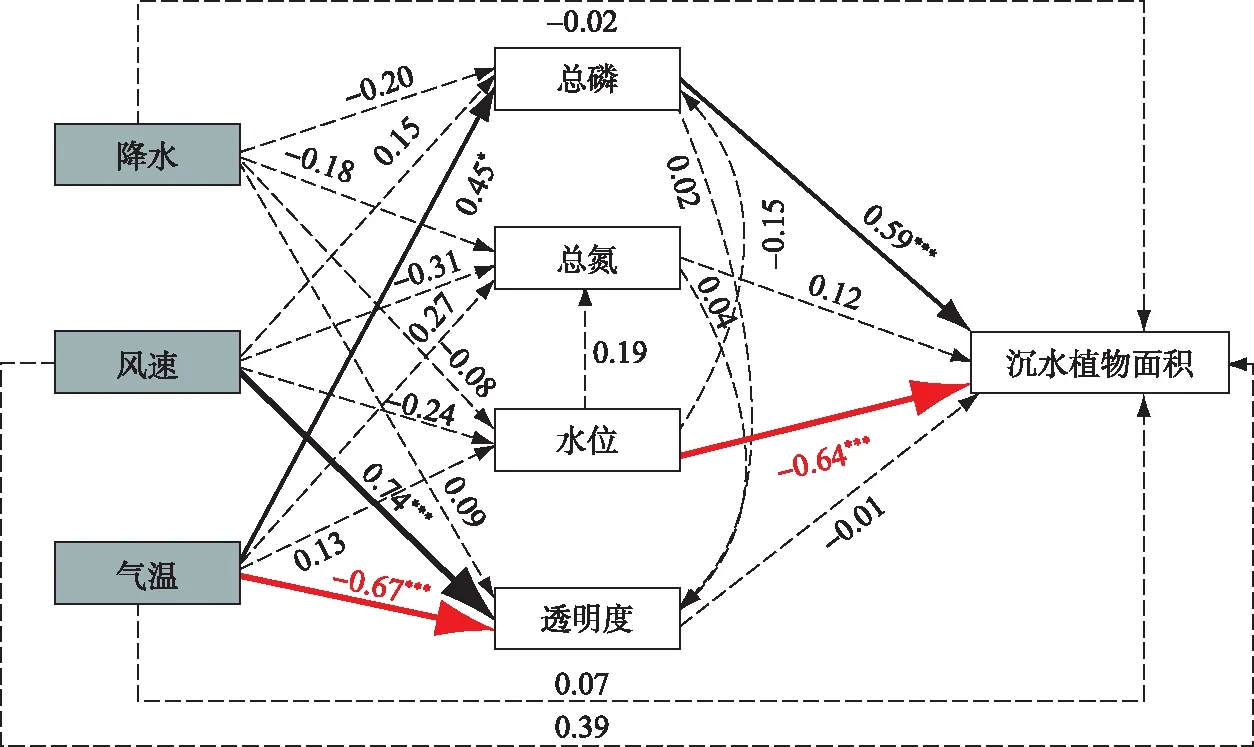
图5 影响因子与沉水植物面积变化关系(黑线代表通过显著性检验的正相关关系,红线代表通过显著性检验的负相关关系,虚线则代表未通过显著性检验;*:P<0.05;**:P<0.01;***:P<0.001)Fig.5 Relationship between impact factors and changes in submerged plant area(Black lines represent positive correlations that passed the significance test. Red lines represent negative correlations that passed the significance test. Dashed lines represent those that did not pass the significance test. *: P<0.05; **: P<0.01; ***: P<0.001)
3 讨论
3.1 沉水植被面积变化主要影响因素
前人的研究表明水深是影响沉水植被生长、分布的主要因素[31]. 水深变化可导致水体温度、光照、底质、溶解氧、风浪等众多环境因子的相关变化,同时可引起植物株高、分株数、生物量、化学计量学特征形状改变[32]. 抚仙湖为深水湖泊,沉水植物分布范围受到水深的限制. 受“星云湖抚仙湖出流改道工程”的影响,抚仙湖水位在2008年后明显下降. 在本研究的SEM路径分析中,气候变化对于水位的影响没有通过显著性检验,但在长时间尺度上,抚仙湖区域以气温增高,降水减少,风速增加为主要气候特征,会综合影响抚仙湖水位[33]. 人类活动和气候变化相关的多要素使得抚仙湖水位明显降低,使得原本底质适宜但光照不足的深水区域被沉水植被占据,从而增加了植被覆盖面积. 早期抚仙湖为贫营养湖泊,营养浓度低,沉水植物生长差[30]. 2011年后营养盐浓度的增加,特别是总磷浓度的增加,也是抚仙湖沉水植物增多的重要因素. 抚仙湖沉水植物变化的第一阶段(1987-1995年),抚仙湖中水位低营养,北部湖区沉水植被覆盖率为1.64%;第二阶段(1996-2010年)抚仙湖高水位低营养,沉水植物面积明显减少,水位上升是此阶段沉水植物面积变化的主要原因;第三阶段(2011-2020年)抚仙湖低水位高营养,水位降低,营养增加,共同作用于沉水植物,导致此阶段沉水植物的分布面积急剧增加.
3.2 抚仙湖沉水植物群落结构变化
高戈明等总结了抚仙湖1980-2016年沉水植物群落结构的变化[22],发现1980-1983年间抚仙湖沉水植被以苦草为主要优势种;2005-2015年,轮藻类优势度增加,金鱼藻、黑藻也在增多;2016年黑藻、金鱼藻成为主要优势种,外来物种伊乐藻逐渐增多,轮藻类大量减少. 中国科学院抚仙湖高原深水湖泊研究站也曾报告,2019年5月以来抚仙湖北部沿岸水域附着藻类和耐污染狐尾藻的生物量明显增加. 同时,有研究发现低水位不利于苦草的生长,水位降低,冠层型狐尾藻会成为优势种类[34],水位的降低可能同步影响抚仙湖沉水植被由苦草向狐尾藻的转变. 近十年来抚仙湖水体营养浓度升高,水位下降、气温增加等因素耦合,引起沉水植被覆盖度快速增加的同时也伴随着植被群落结构的转变.
3.3 抚仙湖面临的生态威胁
抚仙湖现阶段处于初级生产力增加,生物群落结构快速变化的关键时期. 早期抚仙湖氮磷浓度较低,沉水植被与藻类均受到低营养抑制. 2011年后,水位降低、外源性氮磷输入,特别是磷的增加促进了沉水植被面积扩张. 水生植物能通过吸收水体营养、去氮固磷[35]、分泌化感物质[36]等过程抑制藻类的生长,光照和营养充足的条件下沉水植物还能促进浮游藻类沉降[37],从而保持湖泊的清水稳态. 当营养负荷增加到一定程度时,沉水植被的生长将受到限制,例如苦草在富营养过程中会被限制生长[38],藻类浓度会迅速增加. 附着藻类通过抑制沉水植物光合作用阻碍其生长. 营养盐浓度增高会显著增加附着藻类生物量,对沉水植物光合作用和生产力的影响增大[39],限制沉水植物的生长,使其减少乃至消失[40]. 抚仙湖沉水植被主要组成由苦草逐渐向狐尾藻转变,以及附着藻增多等现象说明抚仙湖北部水体营养已经过度富集,藻类可能会迅速增长,抚仙湖面临藻类暴发的生态系统突变风险.
依据生态系统稳态转换理论预测[41],一旦生态系统发生突变将难以修复,存在显著滞后效应. 抚仙湖具有补给系数小,换水周期长、沉水植被分布面积小等特点,入湖营养极易在沉积物中累积[42]. 目前的研究表明,抚仙湖生态系统正处于富营养化状态. 除了本研究中揭示的沉水植物变化,还体现在浮游植物[43]、营养[16]等指标上. 在长期的外力驱动下,湖泊沉积物可能从污染物的汇转变为污染源,抚仙湖水生生物、水体营养以及沉积物营养释放之间的正反馈机制可能不断增强,从而加速水生态环境的改变,发生突变. 然而,目前相关研究较少. 为了避免生态系统突变的发生,抚仙湖现阶段应严格控制入湖营养盐排放,进行高分辨率水质、水位、沉水植物、浮游植物、附生植物等变化的监测,优化管控水量,依据污染状况和环境压力分区进行保护管理[14],抑制抚仙湖富营养化趋势.
本研究指出抚仙湖北部湖区由于水位降低,营养增加以及气候因素的影响,共同导致抚仙湖北部湖区沉水植被面积增加以及群落结构的改变. 城市化、人口增加、工业发展等人为干扰改变了湖泊理化性质,同样对于沉水植物的变化有强烈影响,然而,受制于湖泊监测时长和范围的限制以及人类活动数据分辨率较低的问题,人类活动的影响作用难以准确指示,使得深入提取驱动湖泊状态变化的内部机制和关键过程受限,生态环境因子之间的关系也难以定量精确评估,只能通过相关关系来说明环境变化过程中各因子之间的联系. 在未来的研究中,应该在更丰富数据的基础上,通过模型模拟抚仙湖生态系统对气候变化和人类活动的响应过程,充分揭示湖泊状态变化机制和关键过程,更精准地为抚仙湖生态环境保护提供科学支撑.
4 结论
1)1987-2020年,抚仙湖流域风速、气温明显升高,降水有减少趋势;水体营养浓度从2011年后持续增加;水位在2008年后显著降低. 遥感解译的抚仙湖北部湖区沉水植被面积在2011年后明显增大.
2)水位、营养和气温共同主导了抚仙湖北部湖区沉水植被覆盖度的变化. 近十年,水位降低,营养增加已经促使了抚仙湖沉水植物显著增加.
3)抚仙湖沉水植被覆盖度增长、藻类生物量增多,沉水植物群落演替等,均表明湖泊正逐步富营养化并具有藻类暴发的风险. 相关部门应高度重视对于抚仙湖的管理和监测,建立水量、水质、水生态一体化监测体系,为模型模拟抚仙湖生态系统演变提供数据基础.
致谢:感谢吴庆龙老师的写作指导,感谢赵钰和徐敏在数据分析方面提供的帮助.
