烯效唑浸种对干旱胁迫下燕麦种子萌发及幼苗抗旱生理特性的影响
2022-11-09贺立恒郝杰史敏莉贾小云宋喜娥
贺立恒,郝杰,史敏莉,贾小云,宋喜娥*
烯效唑浸种对干旱胁迫下燕麦种子萌发及幼苗抗旱生理特性的影响
贺立恒1,郝杰2,史敏莉1,贾小云3,宋喜娥1*
(1.山西农业大学 农学院,山西 太谷 030801;2.山西农业大学 草业学院,山西 太谷 030801;3.山西农业大学 生命科学学院,山西 太谷 030801)
【目的】探究不同质量浓度烯效唑浸种对2个燕麦品种种子萌发、幼苗生长和抗旱生理特性的影响。【方法】采用二因素随机试验,2个燕麦品种分别为0117-119(V1)和YY11-18(V2),烯效唑设4个质量浓度,分别为0 mg/L(U1)、15 mg/L(U2)、30 mg/L(U3)和45 mg/L(U4)。【结果】V1U1处理的发芽势为52.00%,显著高于其他处理(<0.05);V1U2处理的发芽率为72.67%,显著高于其他处理(<0.05)。V2U4处理的发芽势和发芽率均最低,分别为18%和36%。苗高、最大根长及冠层干质量均表现为V1U1处理最高,V2U4处理最低。V1U2处理的侧根数、根干质量及根冠比均最高,分别为6.30个、4.33 mg和0.34,均显著高于其他各处理(<0.05)。V1U2处理的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性、根系活力和脯氨酸量均最高,分别达到150.22 U/g、34.83 U/g、387.35 U/g、239.43 µg/(g·h)和298.88 µg/g,均显著高于其他各处理(<0.05),但是,V1U2处理的丙二醛量则显著低于其他处理(<0.05),为7.28 nmol/g。V1U4处理的叶绿素a、叶绿素b和总叶绿素量最高,分别为1.51、0.58 mg/g和2.12 mg/g,均显著高于其他处理(<0.05)。因此,适宜质量浓度烯效唑处理可促进燕麦根系的生长发育,并通过调控燕麦植物体内保护酶的活性,有效清除活性氧来降低干旱对植株引起的伤害,促进有机物质的合成,提高燕麦的抗旱能力。【结论】综上可知,燕麦品种0117-119的抗旱能力优于品种YY11-18,15 mg/L烯效唑质量浓度浸种最好,品种0117-119和15 mg/L烯效唑的处理组合较优。
燕麦;烯效唑;干旱胁迫;种子萌发;抗旱生理
0 引 言
【研究意义】燕麦是世界上第五大禾本科作物,在我国北方高寒地区有着久远的种植历史。目前,我国栽培的燕麦大多为裸燕麦,在黄土高原地区又叫莜麦。燕麦既是粮食作物,也是饲料作物,其营养、保健价值和饲用价值较高。燕麦种子中含有氨基酸、粗脂肪、膳食纤维等多种营养元素,具有预防冠心病和降低血清胆固醇的功效[1]。燕麦种子麸皮中的可溶性纤维和β-糊精具有降低血脂、保护肠胃、降低血糖的特殊功效,在功能食品的开发利用上具有很高的应用价值[2]。近年来,随着种植业结构调整与农业供给侧结构性改革,燕麦种植面积逐年提升,但是受自然和气候条件的制约,燕麦产量普遍较低而且不稳定,成为制约燕麦产业发展的重要因素之一。在我国燕麦主产区,水资源严重短缺,干旱灾害发生频繁,尤其是春季,这个时期的干旱直接影响燕麦种子的萌发及幼苗的生长[3]。
【研究进展】化学调控技术与常规栽培措施配套使用是抵御干旱的有效途径之一。烯效唑作为一种三唑类植物生长调节剂,在调节植物的逆境生理代谢方面起着非常重要的作用,现已在作物生产中得以广泛使用[4-5]。研究[6-7]表明,烯效唑可促进植物淀粉和干物质积累、提高叶片中光合色素含量和植物光合速率、调节植物内源激素水平。目前,烯效唑在农业生产上的应用主要有浸种和叶面喷施2种方法,其使用方法与效果因不同作物而异。
烯效唑浸种处理后,许多作物在逆境条件下都表现出良好的生存能力,如:幼苗长势旺盛,增产效果明显。闫艳红等[8]研究表明,在不同水分胁迫下,烯效唑浸种能够提高大豆出苗率、茎粗和根冠比,增加叶片中的脯氨酸与可溶性糖量,降低丙二醛量,降低脂质过氧化程度,提高保护性酶的活性。刘丽琴等[9]研究表明,适宜质量浓度的烯效唑浸种可显著增强干旱胁迫环境下红小豆幼苗的光合作用、抗氧化和渗透调节能力,有效促进红小豆植株的生长,提高植株的抗旱性和产量。石艳华等[10]研究发现,在不同水分条件下,经烯效唑浸种处理的苦荞作物的株穗数、株粒数、百粒质量均有不同程度的提高,其抗旱能力也有明显的提升。【切入点】燕麦种子萌发和苗期生长阶段对干旱胁迫较为敏感。烯效唑浸种对植物抗逆性影响的研究,目前主要集中在小麦、玉米、大豆等作物上,对燕麦生长影响方面的研究还较少。【拟解决的关键问题】为此,采用15%的PEG-6000模拟干旱环境,研究不同质量浓度烯效唑浸种对0117-119和YY11-18燕麦种子萌发及幼苗抗旱生理特性的影响,为我国北方干旱区燕麦的抗旱生理调控和生产提供理论依据和技术指导。
1 材料与方法
1.1 植物材料
供试燕麦品种0117-119(V1)和YY11-18(V2)的种子由山西农业大学高寒区作物研究所提供。
1.2 试验设计
试验在山西农业大学种子实验室及旱作栽培重点实验室进行。采用二因素随机区组的试验方法进行,试验因素为燕麦品种和烯效唑质量浓度梯度,其中4个烯效唑浸种质量浓度设置为:0 mg/L(U1)、15 mg/L(U2)、30 mg/L(U3)和45 mg/L(U4)。
用质量分数为15%的PEG6000模拟干旱胁迫,通过种子标准发芽试验检测发芽特性。挑选颗粒饱满、无病虫害、种胚完整、大小一致的燕麦种子,采用质量分数为10%的双氧水消毒30 min后,用蒸馏水冲洗5次,随后用滤纸吸干表面水分。将已消毒的种子置于不同质量浓度的烯效唑溶液中浸种24 h,浸种完成后,将种子均匀整齐摆放在铺有2层滤纸、大小为20 cm×15 cm的发芽盒内。每个发芽盒内摆放100粒,加入20 mL质量分数为15%的PEG6000溶液,并将其放入培养箱内培养(培养条件设为光照8 h,暗培养16 h,光强1 000 lx,恒温20 ℃),每个处理3次重复。每3天更换1次滤纸及PEG溶液。5 d后测定发芽势,10 d后测定发芽率、苗高、最大根长、侧根数、冠层干质量、根干质量、根冠比和幼苗丙二醛量、叶绿素量、根系活力、脯氨酸以及SOD、POD和CAT量。
1.3 测定指标及方法
发芽势与发芽率:
发芽势(%)=发芽试验初期(5 d)正常幼苗数/供试种子总数×100
发芽率(%)=发芽试验末期(10 d)正常幼苗数/供试种子总数×100
冠层干质量、根干质量和根冠比:每个处理选取10株幼苗,测量每株幼苗的苗高及根长,然后经杀青、烘干后分别称量冠层干质量和根干质量,计算根冠比。
根冠比=根干质量/冠层干质量
根系活力的测定采用氯化三苯基四氮唑(TTC)法[11],叶绿素量的测定采用分光光度法[12],脯氨酸量的测定采用酸性茚三酮比色法[13],丙二醛(MDA)量的测定用硫代巴比妥酸(TBA)法[14],超氧化物歧化酶(SOD)活性测定采用氮蓝四唑法[15],过氧化物酶(POD)活性测定采用愈创木酚溶液比色法[15],过氧化氢酶(CAT)活性测定参照Graham[16]等的方法。
1.4 数据分析方法
采用SPSS 23进行方差分析,并用LSD法进行多重比较。采用Origin 2021(v9.8.0.200)作图。
2 结果与分析
2.1 不同处理对燕麦种子萌发的影响
图1为不同质量浓度烯效唑处理下燕麦种子的发芽势和发芽率,图中不同小写字母表示处理组合间差异显著(<0.05)。由图1可知,2个品种燕麦种子发芽势和发芽率随烯效唑质量浓度的增加变化趋势基本一致,发芽势的表现趋势均为U1处理最高,随着烯效唑质量浓度升高,发芽势显著降低(<0.05),V1处理降幅为63.46%,V2处理降幅为62.24%;

图1 各处理燕麦种子发芽势和发芽率
发芽率表现为U2处理最高,2个燕麦品种种子的发芽率分别比U1处理提高了15.55%和5.37%;随着烯效唑质量浓度增加,发芽率显著下降(<0.05),但不同品种间表现略有不同,其中V1U2处理和V1U3处理间无显著差异,V2U1处理和V2U2处理的发芽率无显著差异。U4质量浓度下2个品种的发芽率均最低,且显著低于其他各处理。V1U2处理的发芽率最高,为69.33%,其次是V2U2处理,发芽率为65%,V2U4处理的发芽势和发芽率均最低,分别为18%和36%。
2.2 不同处理对燕麦幼苗形态指标的影响
表1为各处理燕麦幼苗形态指标。由表1可知,2个燕麦品种的苗高、最大根长及冠层干质量均为U1处理最高;随烯效唑质量浓度提高,苗高、最大根长及冠层干质量均逐渐降低,其中苗高降幅为74.22%,最大根长降幅为50%,冠层干质量降幅为55.78%;同时2个燕麦品种的侧根数、根干质量及根冠比均显著增加,其中侧根数的增幅为46.51%,根干质量增幅为85.84%和根冠比增幅为161.54%。由表1可知,V1U1处理的苗高、最大根长及冠层干质量均最高,其次为V2U1处理;V2U4处理的苗高、最大根长及冠层干质量均最低。但是,侧根数、根干质量及根冠比均表现为V1U2处理最高,分别达到6.30个、4.33 mg和0.34,均显著高于其他处理(<0.05),其次为V2U2处理。
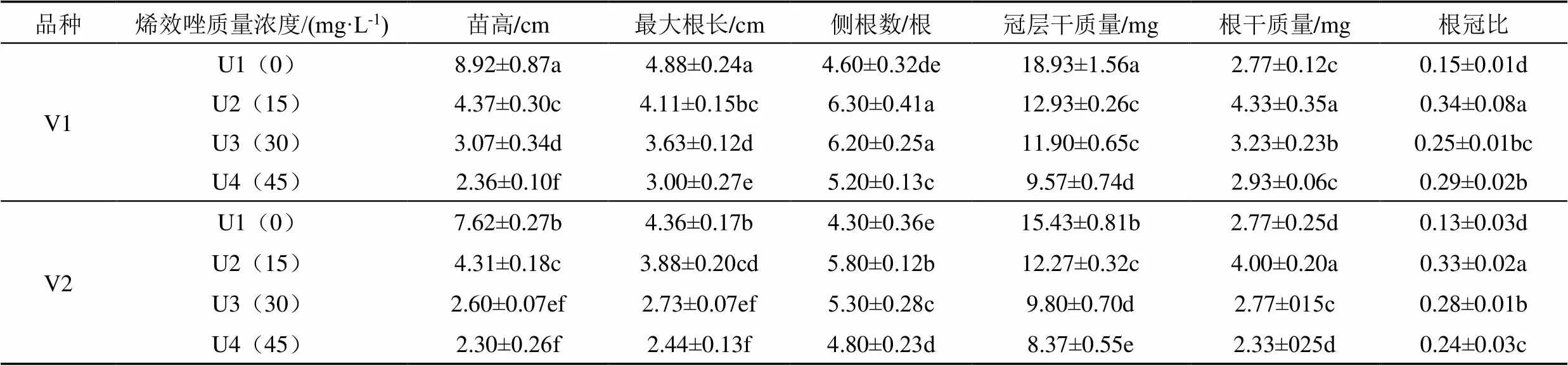
表1 各处理燕麦幼苗形态指标
注 表中纵向数字后面的不同小写字母表示差异显著(<0.05,下同)。
2.3 不同处理对燕麦幼苗抗氧化酶活性的影响
干旱胁迫下,不同质量浓度烯效唑浸种对2个品种燕麦幼苗抗氧化酶活性的影响见图2。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性均为V1U2处理最高,分别达到150.22、34.83、387.35 U/g,除与V2U2处理的SOD活性无显著差异外,与其他处理间差异均显著(<0.05)。当烯效唑质量浓度增加到U3处理和U4处理时,2个燕麦品种的SOD、POD和CAT活性均呈下降趋势,但是,除V1U4处理外,仍均高于V1U1处理和V2U1处理。V2U1处理丙二醛量最高为23.07 nmol/g,V2U4处理次高。然后,随烯效唑质量浓度升高2个燕麦品种的丙二醛量均呈先下降后增加的趋势,但是,仍均分别低于V1U1处理和V2U1处理。V1U2处理的丙二醛量最低为7.28 nmol/g。
2.4 不同处理对燕麦幼苗叶绿素量的影响
由图3可知,除V2U4处理外(V2U3处理与V2U4处理间差异不显著),2个燕麦品种幼苗的叶绿素a、叶绿素b和总叶绿素量均随烯效唑质量浓度增加而显著增加(<0.05)(图3)。不同质量浓度烯效唑浸种处理下,V1处理的叶绿素a、叶绿素b和总叶绿素量均分别高于相应烯效唑质量浓度下的V2处理。V2U1处理的叶绿素a、叶绿素b和总叶绿素量均最低,而V1U4处理的叶绿素a、叶绿素b和总叶绿素量最高,均显著高于其他各处理(<0.05),分别达到1.51、0.58、2.12 mg/g,增幅分别为21.20%、58.86%和29.38%。

图2 不同处理对干旱胁迫下燕麦幼苗抗氧化酶活性的影响

图3 不同处理对干旱胁迫下燕麦幼苗叶绿素的影响
2.5 不同处理对根系活力及脯氨酸量的影响
干旱胁迫下,不同质量浓度烯效唑浸种对2个燕麦品种根系活力及脯氨酸量的影响结果见图4。
2个燕麦品种幼苗的根系活力和脯氨酸量(图4)均随烯效唑质量浓度的增加而呈先升高后降低的趋势,但仍均显著高于U1处理(<0.05)。V2U1处理的根系活力和脯氨酸量均最低,而V1U2处理的根系活力和脯氨酸量均显著高于其他处理(<0.05),分别为239.43 µg/(g·h)和298.88 µg/g,其次是V2U2处理。与V2U1处理相比,增幅分别为53.03%和91.21%。

图4 各处理燕麦根系活力和脯氨酸量
3 讨 论
干旱胁迫会引起种子萌发和成苗困难,是影响农业生产的主要因子。李文娆等[17]研究发现,水分胁迫影响了发芽率及幼苗的生长。燕麦因具有较高的营养和保健功能而受到人们的青睐,其需求量也正在不断增加[1]。中国燕麦主要种植在北方干旱和半干旱地区,降水较少,季节性干旱频繁发生,特别是播种期降水量尤为稀少,而燕麦种子萌发和苗期生长阶段又对水分胁迫较为敏感,所以干旱已成为限制我国燕麦高产稳产的重要原因之一[18]。
目前,烯效唑已被广泛用于调控作物的生长发育和提高作物的抗逆性。李波[20]发现烯效唑处理能够显著提高水分胁迫下大豆的壮苗率及抗旱能力,从而显著提高其产量。本研究也发现15 mg/L质量浓度的烯效唑浸种显著提高了干旱胁迫下燕麦种子的发芽率,但是降低了种子的发芽势(图1),这更加有利于燕麦种子避开发芽前期短暂的阶段性干旱。
关华等[21]在研究烯效唑对小麦苗期生长的调控效应中发现,烯效唑浸种能够增加干旱胁迫下小麦的最大根长。张永清等[22]发现,烯效唑浸种处理对谷子苗期的最大根长有抑制作用。本试验中,烯效唑浸种处理显著降低了干旱胁迫下燕麦幼苗的最大根长,这可能与作物种类和生长特性有关。本试验还发现,随着烯效唑处理质量浓度的提高,燕麦幼苗的苗高和冠层干质量都显著降低,同时根冠比显著增加,这与烯效唑在高粱、玉米上的研究结果一致[23-24]。根冠比的增加有利于幼苗通过吸收利用有限的水分来维持其自身的成活和生长。试验中,当烯效唑质量浓度为15 mg/L时,干旱胁迫下燕麦幼苗的侧根数、根干质量、根冠比均达到最大值(表1),表明适宜质量浓度的烯效唑对燕麦根系及地上部器官的生长有良好的调控作用,通过抑制苗的上部生长,将光合产物优先运往根部来促进下部根系生长,这样利于在干旱环境下有效吸收土壤水分和无机盐等营养成分,提高燕麦的耐旱性。
根系作为植物吸收土壤水分和营养的主要器官,其活力直接影响植物对营养物质的吸收,进而影响其产量和品质[29]。脯氨酸是细胞内的渗透调节物质,在遭受环境胁迫时,可以清除自由基,保护细胞免受伤害[30]。本试验中,烯效唑浸种处理均提高了2个燕麦品种干旱胁迫下的根系活力和脯氨酸量(图4),这与李秋等[31]研究结果基本一致。
干旱会引起植物细胞渗透压的变化,从而导致植物细胞脱水和植物体内活性氧的增加[25-27]。许多研究表明抗氧化保护酶活性的提高可以清除植物体内的活性氧,提高植物的抗旱性。丙二醛是植物细胞膜脂过氧化产物,丙二醛量的增加抑制了抗氧化酶的活性,所以常也被用来作为衡量植物抗旱性的指标之一[28]。本试验发现,不同质量浓度烯效唑浸种处理均可显著提高干旱胁迫条件下2个品种燕麦幼苗SOD、POD和CAT保护酶的活性,降低丙二醛量。当烯效唑质量浓度为15 mg/L时,3种保护酶的活性最高(图2),这与刘丽琴等[9]研究一致。可见,适宜质量浓度的烯效唑能够调控燕麦体内保护酶活性,有效清除活性氧来降低干旱对植物体引起的伤害,从而提高燕麦的抗旱能力。
4 结 论
燕麦品种0117-119的抗旱能力优于品种YY11-18,15 mg/L烯效唑质量浓度浸种最好,品种0117-119和15 mg/L烯效唑的处理组合较优。在干旱地区燕麦作物生产过程中,烯效唑浸种质量浓度以15 mg/L为宜。
[1] BERG Aloys, KÖNIG Daniel, DEIBERT Peter, et al. Effect of an oat bran enriched diet on the atherogenic lipid profile in patients with an increased coronary heart disease risk[J]. Annals of Nutrition and Metabolism, 2003, 47(6): 306-311.
[2] 李春花, 梅春光. 燕麦的营养成分及营养保健价值的研究进展[J]. 现代农业, 2010, 2(5): 134-135.
LI Chunhua, MEI Chunguang. Advances in research on nutrient composition and nutritional health value of oats[J]. Modern Agriculture, 2010, 2(5): 134-135.
[3] 王文义. 山西气候与干旱变化及其影响因子[D]. 南京: 南京信息工程大学, 2010.
WANG Wenyi. Climate and drought change in Shanxi and its impact factors[D]. Nanjing: Nanjing University of Information Science & Technology, 2010.
[4] 白朴, 黄安波, 许立奎, 等. 烯效唑浸种对晚杂控长促蘖效应研究[J]. 杂交水稻, 1995(3): 25-27.
BAI Pu, HUANG Anbo, XU Likui, et al. Effects of uniconazole soaking seeds on growth control and tilling promoting in late hybrid rice[J]. Hybrid Rice, 1995(3): 25-27.
[5] LIU Yang, FANG Yang, HUANG Mengjun, et al. Uniconazole-induced starch accumulation in the bioenergy crop duckweed (Landoltia punctata) I: transcriptome analysis of the effects of uniconazole on chlorophyll and endogenous hormone biosynthesis[J]. Biotechnology for Biofuels, 2015, 8(1): 57.
[6] ZHOU W, YE Q. Physiological and yield effects of uniconazole on winter rape (L.)[J]. Journal of Plant Growth Regulation, 1996, 15(2): 69-73.
[7] GONG Wanzhuo, ZHANG Zhengyi, YANG Wenyu, et al. Effect of uniconazloe for dry seed treatment on morphological characteristics and yield of soybean[J]. Soybean Science, 2007, 26(3): 372-376.
[8] 闫艳红, 李波, 杨文钰. 烯效唑浸种对大豆苗期抗旱性的影响[J]. 中国油料作物学报, 2009, 31(4): 480-485.
YAN Yanhong, LI Bo, YANG Wenyu. Effects of uniconazole soaking on drought tolerance of soybean seedling[J]. Chinese Journal of Oil Crop Sciences, 2009, 31(4): 480-485.
[9] 刘丽琴, 张永清, 李鑫, 等. 烯效唑浸种对干旱胁迫下红小豆生长及其根系生理特性的影响[J]. 西北植物学报, 2017, 37(1): 144-153.
LIU Liqin, ZHANG Yongqing, LI Xin, et al. Influence of seed soaking with uniconazole on growth and root physiological characteristics of adzuki bean under drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(1): 144-153.
[10] 石艳华, 张永清. 烯效唑浸种对不同水分条件下苦荞生长的影响[J]. 湖北农业科学, 2015, 54(6): 1 289-1 295.
SHI Yanhua, ZHANG Yongqing. Effects of soaking seed with uniconazole on the growth of tartary buckwheat under different water conditions[J]. Hubei Agricultural Sciences, 2015, 54(6): 1 289-1 295.
[11] 白宝璋, 金锦子. 玉米根系活力TTC测定法的改良[J]. 玉米科学, 1994, 2(4): 44-47.
BAI Baozhang, JIN Jinzhi. Improvement of TTC method determining root activity in corn[J]. Journal of Maize Sciences, 1994, 2(4): 44-47.
[12] 张宪政. 植物叶绿素含量测定—丙酮乙醇混合液法[J]. 辽宁农业科学, 1986(3): 26-28.
ZHANG Xianzheng. Determination of chlorophyll content in plants-Acetone ethanol mixture method[J]. Liaoning Agricultural Sciences, 1986(3): 26-28.
[13] 职明星, 李秀菊. 脯氨酸测定方法的改进[J]. 河南科技学院学报, 2005, 41(4): 355-357.
ZHI Mingxing, LI Xiuju. Improvement of the method for measuring proline content[J]. Journal of Henan Institute of Science and Technology, 2005, 41(4): 355-357.
[14] 孔祥生, 易现峰. 植物生理学实验技术[M]. 北京: 中国农业出版社, 2008: 32-35.
KONG Xiangsheng, YI Xianfeng. Plant physiology experiment technology[M]. Beijing: China Agricultural Press, 2008: 32-35.
[15] 张志良, 瞿伟菁, 李小方. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2009: 48-50.
ZHANG Zhiliang, QU Weijing, LI Xiaofang. Plant physiology experiment guide[M]. Beijing: Higher Education Press, 2009: 48-50.
[16] PATTERSON BRIAN D, PAYNE Linda A, CHEN Yizhu, et al. An inhibitor of catalase induced by cold in chilling-sensitive plants[J]. Plant Physiology, 1984, 76(4): 1 014-1 018.
[17] 李文娆, 张岁岐, 山仑. 水分胁迫下紫花苜蓿和高粱种子萌发特性及幼苗耐旱性[J]. 生态学报, 2009, 29(6): 3 066-3 074.
LI Wenrao, ZHANG Suiqi, SHAN Lun. Seeds germination characteristics and drought-tolerance of alfalfa and sorghum seedling under water stress[J]. Acta Ecologica Sinica, 2009, 29(6): 3 066-3 074.
[18] 张宇君, 赵丽丽, 王普昶, 等. 盐旱交互对燕麦种子萌发及幼苗生理特性的影响[J]. 草业学报, 2018, 27(5): 141-152.
ZHANG Yujun, ZHAO Lili, WANG Puchang, et al. Effects of interaction between Ca2+salt and drought stress on seed germination and seedling physiology of oats[J]. Acta Prataculturae Sinica, 2018, 27(5): 141-152.
[19] SHEYKHBAGLOU Roghayyeh, RAHIMZADEH Saeede, ANSARI Omid, et al. The effect of salicylic acid and gibberellin on seed reserve utilization, germination and enzyme activity of sorghum (L.) seeds under drought stress[J]. Journal of Stress Physiology & Biochemistry, 2014, 10(1): 234-236.
[20] 李波. 不同水分胁迫下烯效唑浸种对大豆苗期生长及产量的影响[D]. 雅安: 四川农业大学, 2006.
LI Bo. Effects of uniconazole applied to seed on seedling growth and yeild of soybean under different water stress[D]. Ya’an: Sichuan Agricultural University, 2006.
[21] 关华, 杨文钰. 烯效唑对小麦苗期生长的调控效应[J]. 种子, 2002, 18(3): 38-41.
GUAN Hua, YANG Wenyu. Regulatory effect of uniconazole on wheat seedling growth[J]. Seed, 2002, 18(3): 38-41.
[22] 张永清, 裴红宾, 刘良全, 等. 烯效唑浸种对谷子植株生长发育的效应[J].作物学报, 2009, 35(11): 2 127-2 132.
ZHANG Yongqing, PEI Hongbin, LIU Liangquan, et al. Effect of seed soaking with uniconazole on growth and development of foxtail millet plant[J]. Acta Agronomica Sinica, .2009, 35(11): 2 127-2 132.
[23] 张治安, 陈展宇, 李大勇, 等. 烯效唑浸种对高粱幼苗生理指标的影响[J].吉林农业大学学报, 2004, 26(6): 599-602.
ZHANG Zhian, CHEN Zhanyu, LI Dayong, et al. Effects of soaking seeds in uniconazole solution on physiological indexes of sorghum seedlings[J]. Journal of Jilin Agricultural University, 2004, 26(6): 599-602.
[24] 李青苗, 杨文钰. 烯效唑浸种对玉米壮苗的生理效应[J]. 玉米科学, 2003, 11(3): 66-69.
LI Qingmiao, YANG Wenyu. Physiological effects of soaking seed with uniconazole on high quality seedling of maize[J]. Journal of Maize Sciences, 2003, 11(3): 66-69.
[25] BARTELS Dorothea, SUNKAR Ramanjulu. Drought and salt tolerance in plants[J]. Critical Reviews in Plant Sciences, 2005, 24(1): 23-58.
[26] FOYER Christine H, NOCTOR Graham. Oxidant and antioxidant signaling in plants: A re-evaluation of the concept of oxidative stress in a physiological context[J]. Plant Cell & Environment, 2005, 28(8): 1 056-1 071.
[27] TÜRKAN İsmall, BOR Melike, ÖZDEMIR Filiz, et al. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerantGray and drought-sensitiveL. subjected to polyethylene glycol mediated water stress[J]. Plant Science, 2005, 168(1): 223-231.
[28] 戴高兴, 彭克勤, 萧浪涛, 等. 聚乙二醇模拟干旱对耐低钾水稻幼苗丙二醛,脯氨酸含量和超氧化物歧化酶活性的影响[J]. 中国水稻科学, 2006, 20(5): 557-559.
DAI Gaoxing, PENG Keqin, XIAO Langtao, et al. Effect of drought stress simulated by PEG on malonaldehyde, proline contents and superoxide dismutase activity in low potassium tolerant rice seedlings[J]. Chinese Journal of Rice Science, 2006, 20(5): 557-559.
[29] 王秀波, 上官周平. 干旱胁迫下氮素对不同基因型小麦根系活力和生长的调控[J]. 麦类作物学报, 2017, 37(6): 820-827.
WANG Xiubo, SHANGGUAN Zhouping. Effect of nitrogen on root vigor and growth in different genotypes of wheat under drought stress[J]. Journal of Triticeae Crops, 2017, 37(6): 820-827.
[30] YAMADA Mika, MORISHITA Hiromasa, URANO Kaoru, et al. Effects of free proline accumulation in petunias under drought stress[J]. Journal of Experimental Botany, 2005, 56(417): 1 975-1 981.
[31] 李秋, 李立芹. 烯效唑对小麦幼苗生长的影响[J]. 安徽农业科学, 2011, 39(10): 5 715-5 716, 5 719.
LI Qiu, LI Liqin. Effects of uniconazole on wheat seedlings[J]. Journal of Anhui Agricultural Sciences, 2011, 39(10): 5 715-5 716, 5 719.
Soaking Oat Seeds with Uniconazole to Improve Their Germination and Resistance of Seedlings to Drought Stress
HE Liheng1, HAO Jie2, SHI Minli1, JIA Xiaoyun3, SONG Xi’e1*
(1. College of Agriculture, Shanxi Agricultural University, Taigu 030801, China; 2. College of Grassland Sciences, Shanxi Agricultural University, Taigu 030801, China;3. College of Life Sciences, Shanxi Agricultural University, Taigu 030801, China)
【Objective】Promoting technologies are often used to improve seed germination. In this paper, we study the efficacy of soaking oat seeds with uniconazole in improving their germination and resistance of the seedlings against drought stress. 【Method】We compare two oat varieties, 0117-119 (V1) and YY11-18 (V2). Seeds of each variety are soaked by uniconazole at concentrations 0 mg/L (U1), 15 mg/L(U2), 30 mg/L(U3) and 45 mg/L(U4), respectively. The germination pattern and the seedling traits in each treatment is measured. 【Result】The highest germination potential and germination rate of V1 are 52.0% and 72.67% (<0.05), achieved under U1 and U2, respectively. The least germination potential and germination rate are V2 under U4, being 18% and 36%, respectively. V1 under U1 achieves the highest seedling shoot length, maximum root length and crown root dry weight, while V2 under U4 gives the least. The number of lateral roots, root dry weight and root-crown ratio of V1 under U2 are significantly higher than those in other treatments (<0.05), being 6.30, 4.33 mg and 0.34, respectively. V1 under U2 gives the best SOD, POD, CAT, root vigor and proline contents, being 150.22 U/g, 34.83 U/g, 387.35 U/g, 239.43 µg/(g·h)and 298.88 µg/g, respectively, but the lowest malondialdehyde content at 7.28 nmol/g. The contents of chlorophyll a and b, as well as the total chlorophyll in V1 under U4 are significantly higher than those in other treatments (<0.05), being 1.51 mg/g, 0.58 mg/gand 2.12 mg/g, respectively. V1 is more drought resistant than V2. Soaking the seeds by uniconazole at concentration of 15 mg/L is optimal for both varieties, and V1 under U2 works the best. 【Conclusion】Soaking oat seeds with uniconazole can effectively alleviate the damage caused by drought, thereby improving their germination rate. Our results can help design oat seed drill in semi-arid areas that are facing drought stress in northern China.
Oat; uniconazole; drought stress; seed germination; drought-resistant physiology
贺立恒, 郝杰, 史敏莉, 等. 烯效唑浸种对干旱胁迫下燕麦种子萌发及幼苗抗旱生理特性的影响[J]. 灌溉排水学报, 2022, 41(10): 19-25.
HE Liheng, HAO Jie, SHI Minli, et al.Soaking Oat Seeds with Uniconazole to Improve Their Germination and Resistance of Seedlings to Drought Stress[J]. Journal of Irrigation and Drainage, 2022, 41(10): 19-25.
1672 - 3317(2022)10 - 0019 - 07
S519
A
10.13522/j.cnki.ggps.2022182
2022-04-07
国家自然科学基金项目(31571611);山西省重点研发计划项目(201803D221008-6);山西农谷建设科研专项项目(SXNGJSKYZX201703);山西省“1331工程”项目
贺立恒(1974-),男。副教授,博士,主要从事作物生理生态及生长发育调控研究。E-mail: sxndhlh@163.com
宋喜娥(1974-),女。副教授,博士,主要从事为作物化学调控与化学除草研究。E-mail: sxndsxe@163.com
责任编辑:白芳芳
