A Late Pliocene Hipparion houfenense fauna from Yegou,Nihewan Basin and its biostratigraphic significance
2022-11-05LIUJinYiZHANGYingQiCHIZhenQingWANGYongYANGJinSongZHENGShaoHua
LIU Jin-Yi ZHANG Ying-Qi* CHI Zhen-Qing WANG Yong YANG Jin-Song ZHENG Shao-Hua
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044* Corresponding author: zhangyingqi@ivpp.ac.cn)
(2 CAS Center for Excellence in Life and Paleoenvironment Beijing 100044 )
(3 Institute of Geology, Chinese Academy of Geological Sciences Beijing 100037)
(4 Institute of Hydrogeology and Environmental Geology, Chinese Academy of Geological Sciences Shijiazhuang 050061)
Abstract Currently, there are still different views regarding the chronology of the Late Cenozoic deposits in the Nihewan Basin, which results from the contradiction between biostratigraphic correlations based on mammalian fossils and magnetostratigraphic dating results. Biostratigraphic correlations indicate that the aeolian red clay exposed in the Sanggan River canyon, the fluviolacustrine red clay with sands and gravels, and the sandy clay of swamp facies on both sides of the lower reaches of the Huliu River belong to the Upper Pliocene, whereas the magnetostratigraphic dating usually correlates them to the Lower Pleistocene. In October 2011, a collection of mammalian fossils was unearthed from a block of collapsed deposits at Yegou in the Nihewan Basin, which is about 300 m north of the Laowogou section that is well known for the Pliocene mammalian fossils from its lower part. The Yegou fossils are identified herein as 10 species in 9 genera: Nyctereutes tingi, N. sinensis, Pachycrocuta pyrenaica, Homotherium sp., Hipparion(Plesiohipparion) houfenense, Dicerorhinus sp., Muntiacus sp., Axis shansius, Gazella blacki, and Paracamelus sp. The fauna is quite different from the classic Early Pleistocene Nihewan Fauna in composition and provides new evidence for the existence of the Upper Pliocene in the Nihewan Basin. Based on a systematic description of the fauna, its composition and geological age are discussed, and the compositional features of large mammals of the Late Pliocene and the Early Pleistocene mammalian faunas in the Nihewan Basin are summarized.
Key words Nihewan Basin, Upper Pliocene, large mammals, Hipparion (Plesiohipparion)houfenense, biostratigraphy, magnetostratigraphy
1 Introduction
In 2011, CHI Zhen-Qing of Institute of Geology, Chinese Academy of Geological Sciences invited ZHENG Shao-Hua of Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences to carry out a joint investigation in the Nihewan Basin. On October 4th of that year, the team discovered abundant fossils of large mammals at Yegou, north of Daodi Village, which are identified herein as 10 species in 9 genera:Nyctereutes tingi,N. sinensis,Pachycrocuta pyrenaica,Homotheriumsp.,Hipparion(Plesiohipparion)houfenense,Dicerorhinussp.,Muntiacussp.,Axis shansius,Gazella blacki,andParacamelussp. The Yegou locality (N40°9′9.9″, E114°39′19.4″) is located about 300 m north of the Laowogou section, the stratotype of the “Daodi Formation” (Du et al., 1988). The fossils were unearthed from a big block of collapsed deposits, the lithology of which is dark brownish grey silty clay. However, the stratigraphic section is not exposed. Based on the fossils,lithology and altitude, this horizon may be roughly correlated to Layer 9 of the Laowogou section. The latter has yieldedHipparionsp., ?Chilotheriumsp.,Axis shansius,Cervussp.,Antilospirasp., etc. (as listed in Cai et al., 2004 and Cai et al., 2013). If taken as one unified fauna, they would represent aHipparion houfenensefauna with considerable diversity.
On the one hand, the extensive existence of Pliocene strata on both sides of the Huliu River has been indicated by large mammal fossils from Luanshigedagou, Dannangou, and Huabaogou (Huang et al., 1974; Tang, 1980b; Wang, 1982) and small mammal fossils from Yuanzigou, Qijiazhuang, Jiangjungou, Laowogou, Hongyanangou, and Huabaogou (Zhang et al., 2003; Cai et al., 2004, 2013; Li et al., 2008). On the other hand, we have also noted that the magnetostratigraphic dating results published in recent years have placed the fossil bearing layer of Luanshigedagou, Hongya Village in between the “Gauss-Matuyama” boundary and the lower boundary of “Olduvai” (2.58–1.95 Ma); the upper fossil layer of Huabaogou, HBG-I,right above the upper boundary of “Olduvai” (<1.77 Ma); the lower fossil layer of Huabaogou,HBG-II, within “Olduvai” (1.95–1.77 Ma); the fossil layers of DD-1~7 of Daodi or Laowogou between the “Gauss-Matuyama” boundary and the upper boundary of “Olduvai” (2.58–1.77 Ma) (Zhu et al., 2007; Deng et al., 2008; Deng, 2011; Ao et al., 2013). Consequently,apparent contradiction occurs between these results and biostratigraphic correlations based on mammalian fossils. The features of the YegouHipparion houfenensefauna that is made up of only large mammals are also indicative of Pliocene age. Its establishment will accordingly provide additional evidence for the existence of the Pliocene in the Nihewan Basin and for the necessity of cautious reinterpretations of these magnetostratigraphic dating results.
Abbreviations F:AM, the Frick Collection of American Museum of Natural History;IVPP V/V, catalogue number of vertebrates of Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP, CAS); IVPP RV/RV, revised catalogue number of vertebrates of IVPP, CAS; JNTZ, catalogue number of Nanjing Museum;NNNM, catalogue number of Nihewan National Nature Reserve; NWUV, catalogue number of vertebrates of Northwest Universituy; THP, catalogue number of Tianjin Natural History Museum; ZKD, Zhoukoudian; L, length; W, width; Tri, trigonid; Min, minimum; Max,maximum.
2 Systematic descriptions

Material Fragmentary right ramus with all teeth but the i2 and m3 (IVPP V 18833.1,Fig. 1A); anterior portion of the fragmentary left ramus with c–p2 (V 18833.2); right m1 (V 18833.3, Fig. 1B); right p4 (V 18833.4); fragment of the right ramus with the trigonid of the m1 (V 18833.5).
Measurements See Table 1.
Description The portion posterior to the m2 is not preserved on IVPP V 18833.1, so the development of the subangular lobe is unclear. The horizontal ramus, especially the anterior portion, is relatively robust. The ventral profile of the horizontal ramus is almost straight, but the ramus gradually deepens posteriorly. The depth is about 16.0 mm anterior to p3, and about 17.5 mm anterior to the m1. The symphysis is oval-shaped in lingual view, and its posterior margin slightly exceeds the posterior end of the p2 (Fig. 1A2). There are two mental foramina,with the anterior one being bigger and located below the diastema between the p1 and p2, and the posterior one being smaller and right below the p3 (Fig. 1A3).
The lower incisors are, well preserved with the exception of the i2, set obliquely on the ramus with the tip protruding dorsoanteriorly. The i1 is spatula-like with a truncated top and no cuspid lingually. The i2 is lost, leaving an oblong alveolus that is larger than that of the i1. The i3 is basically a triangular pyramid and distingctly larger than the i1 that is characterized by afeeble cuspid developed distally. The lower canine is conical and buccolingually compressed.The p1 is unicuspid and single-rooted. The p2, p3, and p4 are basically similar in both size and morphology. They are buccolingually compressed to some extent and possess a developed principal cusp. No accessory cusps are developed on either the p2 or p3, but there is a remarkable posterior accessory cusp on the p4. The protoconid is the dominant cusp on the m1,whereas the paraconid is slightly smaller and the metaconid the smallest. The talonid of the m1 is simple and bears only the hypoconid and entoconid. The former is higher than the latter. A transverse cristid between them divides the talonid into two parts, with the anterior part bigger than the posterior part (see Fig. 1B1). No accessory cusps or tubercles could be discerned on the talonid of the m1. The m2 is oval-shaped in occlusal view. The anterior half is broader than the posterior half. The metaconid is slightly taller than the protoconid, but they are similar in size. The talonid of the m2 bears a distinct hypoconid and a weak crest-like entoconid. No accessory cuspids are developed. A cingulid is slightly developed anterobuccally on the tooth.
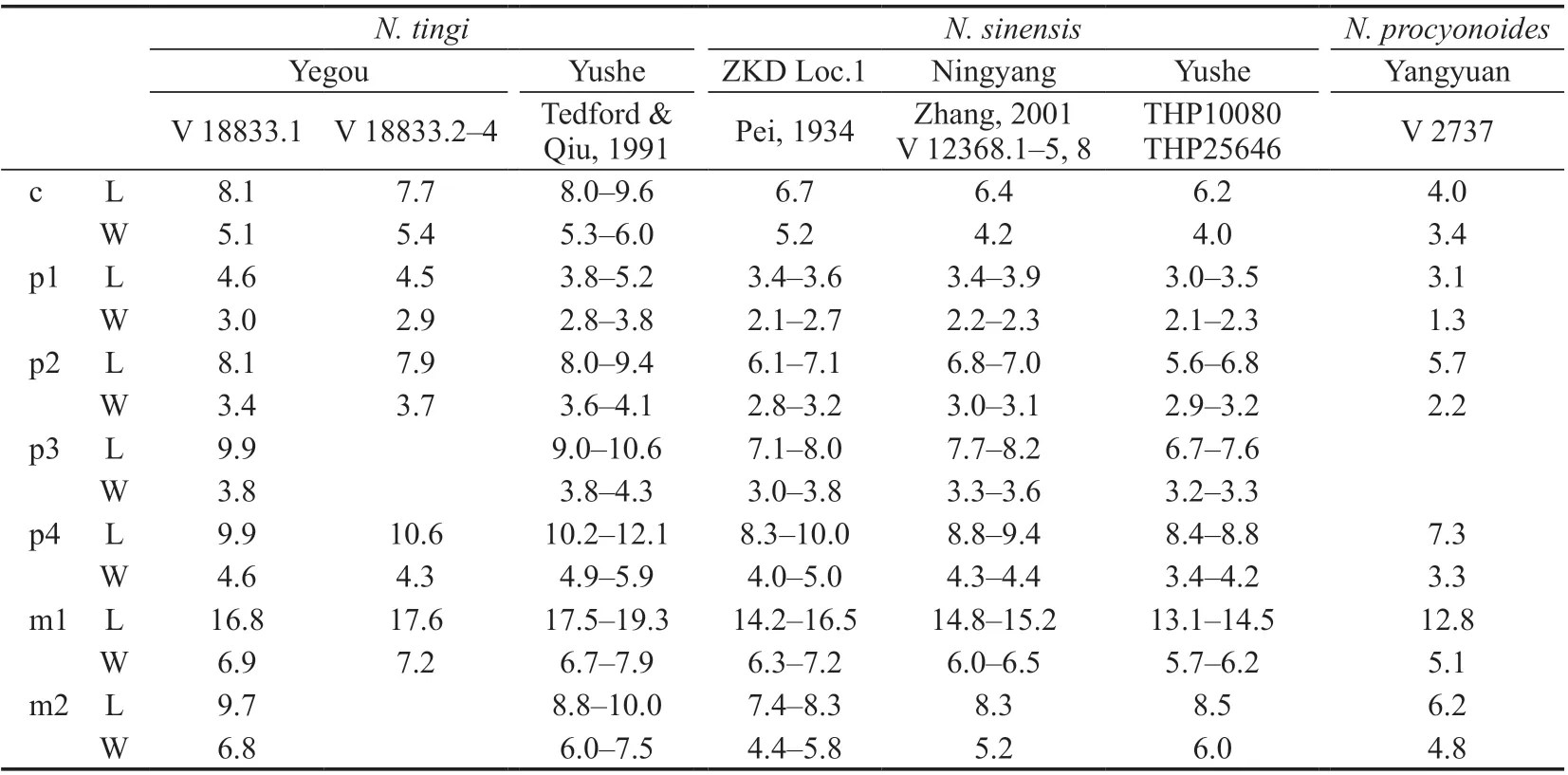
Table 1 Measurements of lower teeth of selected species in Nyctereutes (mm)
IVPP V 18833.2 is a fragmentary left ramus. Only the portion anterior to the p3 is preserved. Its teeth, such as the c, p1, and p2, are generally similar to that of V 18833.1 in both size and shape. V 18833.3 is basically similar to the m1 of V 18833.1 in morphology, but slightly different in the development of a feeble tubercle on the posterior margin of the tooth(Fig. 1B1), which could be considered as the embryonic hypoconulid. V 18833.4 is a right p4,and similar to that of V 18833.1 in both shape and size. V 18833.5 bears a broken right m1.The preserved protoconid and metaconid are the same as that of V 18833.1 and V 18833.4 in morphology and no smaller than the latter two in size.

Fig. 1 Selected specimens of Nyctereutes tingi from Yegou A. fragmentary right ramus with i1, i3, c and p1–m2 (IVPP V 18833.1, 2 cm scale bar);B. right m1 (V 18833.3, 1 cm scale bar); 1. occlusal view, 2. lingual view, 3. buccal view
Comparisons and discussion Based on the morphology and size of the mandible and the lower carnassial (m1) summarized in Tedford et al. (1995, 2009) and Daguenet and Sen(2019), the Yegou specimens are undoubtedly assigned toNyctereutesof Canidae.
The Yegou specimens are obviously larger thanNyctereutessinensisfrom Yushe of Shanxi and Zhoukoudian, and extantN. procyonoidesas well, but closer in size toN. tingifrom Yushe (Table 1).
The ramus of V 18833.1 is relatively robust with the anterior half especially deep, and therefore different fromNyctereutessinensis, which has anteriorly shallow but posteriorly deep ramus so that the dorsal margin of the ramus strongly slopes down anteriorly (see Tedford and Qiu, 1991:fig. 1B; Pei, 1934:fig. 5D). Consequently, the anterior portion of the ramus ofN.sinensisis much slimmer, e.g., the depth of V 350-2 from Loc. 1 of Zhoukoudian is only 13.8 mm anterior to the p3, that of V 12368.1–2 from Ningyang of Shandong is only 11.2–13.3 mm. In terms of this feature, the Yegou specimen V 18833.1 is closer to the holotype ofN. tingiF:AM 97030 from Yushe, the same dimension of which is 15.1 mm.
In addition, the m1 talonid of V 18833.1 and V 18833.3 is relatively simple in construction, bears no other tubercles except the hypoconid and entoconid, and consequently differs fromNyctereutessinensis, since the m1 talonid of the latter somewhat tends to be complicated, e.g., the entoconid ofN. sinensisfrom Loc. 1 of Zhoukoudian normally splits into 2–3 cuspids (Pei, 1934), and an extra cuspid, i.e., the hypoconulid, is even developed on the m1 ofN. sinensisfrom Renzidong, Fanchang of Anhui (Liu and Qiu, 2009). It should be noted here that a transverse cristid developed between the hypoconid and entoconid, and subsequent division of the talonid into two unequal parts are the primary morphological features distinguishingN. tingifromN. sinensis(Tedford and Qiu, 1991). According to such morphological criterion, the Yegou specimens could be referred toN. tingiwithout any doubt,which further confirms the assertion of the existence ofN. tingiin the Nihewan Basin proposed by Farjand et al. (2021).
The m2 is quite simple on V 18833.1 in morphology. Namely a cingulid is slightly developed only on its anterobuccal margin, and the entoconid is crest-like on the talonid with no other accessory cuspids except the entoconid and hypoconid. The Yegou specimens are exactly in accordance withNyctereutes tingiin the morphology of m2 as well.
Based on the morphology and size of both the rami and teeth described above, the Yegou specimens are apparently closer toNyctereutes tingias a whole, although they are fragmentary and lacking certain key diagnostic parts, such as the subangular lobe and angular process of the mandible, etc.
Nyctereutes tingiwas originally established based on the material from the Mazegou and Gaozhuang formations in the Yushe Basin (Tedford and Qiu, 1991). It is a fairly primitive species at an evolutionary stage that can be roughly correlated toN. donnezaniof Ruscinian from Europe. The species was also reported at Lingtai of Gansu (Huang et al., 1993) and Danangou of the Nihewan Basin (Farjand et al., 2021). The known records indicate thatN.tingiis distributed only in Pliocene strata. For that reason, the discovery ofN. tingiat Yegou proves that the Pliocene strata does exist in the Nihewan Basin.
FossilNyctereuteshas a continuous stratigraphic distribution in the strata from Early Pliocene onwards in China and has an evolutionary trend of gradual reduction in body size (Pei,1934; Teilhard de Chardin and Pei, 1941). The dental measurements ofN. tingifrom the Yushe Basin clearly reveal such a trend: teeth from the Gaozhuang Formation are generally larger than those from the Mazegou Formation (based on WANG Xiao-Ming’s unpublished data). In dental size, the material from Yegou is much closer to that from the Mazegou Formation (Late Pliocene), but smaller than that from the Gaozhuang Formation (Early Pliocene).
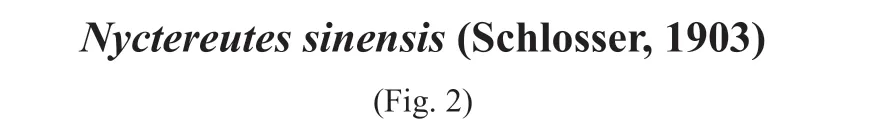
Material Fragmentary cranium lacking the rostrum (IVPP V 18834.1, Fig. 2),fragmentary right ramus with the m2 (V 18834.2), fragmentary right ramus with only alveoli of the p1–3 preserved (V 18834.3)
Measurements See Table 2.
Description V 18834.1 is an incomplete cranium. The nasals, premaxillae and maxillae are broken off, and both of the zygomatic arches and the right tympanic bulla are also missing.It is distinctly larger than the extant speciesNyctereutesprocyonoides. In lateral view, the dorsal profile of the frontal and parietal first runs posterodorsally and reaches its highest point approximately above the postglenoid process, then it runs horizontally posteriorly (Fig. 2D, E).In dorsal view, the frontal area is fairly broad, and its maximal breadth between the postorbital processes is 53.5 mm, which is comparable to the greatest neurocranium breadth. The central portion of both frontals bulges strongly. Consequently, a deep depression is formed along the frontal suture between the frontals. The vulpine groove is also developed on the surface adjacent to the frontal postorbital process, which indicates that the frontal sinus is developed but does not expand into the tip of the postorbital process. The postorbital process is rather robust. The posterodorsal rim of the orbit is not straight but curved and concave. The strong and ridge-like parasagittal crest also makes a concave curve and extends from the postorbital process posteromedially and joins its counterpart slightly posterior to the frontoparietal suture, which makes the postorbital process very pointed (Fig. 2A). The sagittal crest is also massive and sticks out of the parietal surface about 3.0 mm. The sagittal crest, especially its anterior portion, is broad, and a central longitudinal groove runs throughout along the midline. The postorbital constriction is not strong, and the width is 37.1 mm, which is broader than the distance between the orbits (less than 33.2 mm) and about the same as the braincase width. The surface of the parietal is relatively smooth, and not as rough as that of the extant species. In ventral view, the tympanic bulla is strongly inflated, especially at the middle and posterior portions. The anterolateral side is less dilated, and the anterior margin aligns with the postglenoid process and does not exceed it (Fig. 2B).
V 18834.2 is a fragmentary ramus with only the m2 still in position. The dorsal profile of the horizontal ramus significantly slopes down anteriorly. As a result, the anterior portion of the ramus is shallower, with a depth of only 14.4 mm anterior to the p3. The m2 (about 5.9 mm in width) is similar to that of V 18833.1 ofNyctereutes tingiin morphology, but evidently narrower. A cingulid is developed anterobuccally, and more distinct than in V 18833.1. The metaconid is quite small but discernible.

Fig. 2 Fragmentary cranium of Nyctereutes sinensis (IVPP V 18834.1) from Yegou A. dorsal view, B. ventral view, C. occipital view, D. left lateral view, E. right lateral view
V 18834.3 only has preserved the portion anterior to the p4, including the alveoli of p1–3.The anterior portion of the ramus is shallower, and the depth is about 13.2 mm anterior to the p3.
Comparisons and discussion Although the cranium of V 18834.1 is incomplete, the preserved portion could be compared with the species inNyctereutes. It is distinctly larger than the extantN. procyonoides, but roughly comparable in size toN. sinensisfrom Yushe of Shanxi(Table 2).
Regarding the cranium morphology, V 18834.1 appears to have combined characteristics of bothNyctereutessinensisandN. tingi. On the one hand, V 18834.1 is quite similar to the specimen JNTZ 7571 ofN. sinensisfrom Tuozidong, Nanjing in the following aspects: 1)almost equal size; 2) robust, pointed and protruding postorbital process; 3) curved and ridgelike parasagittal crest; 4) high and broad sagittal crest with a central longitudinal groove running throughout; and 5) anterior margin of the tympanic bulla not exceeding the postglenoid process (Liu et al., 2007:26, fig. 2.4). Moreover, the cranium V 18834.1 also resembles that ofN. sinensis(=Liu, 2019:Nyctereutescf.N. tingi) from the Nihewan Basin described by Liu(2019), such as NNNM 13–18 (see Liu, 2019: fig. 23), in all the features listed above except for the 5th one. Nevertheless, it is worth noting herein that neither central bulging of the frontal nor the depression along the frontal suture could be discerned in JNTZ 7571 and NNNM 13–18.
On the other hand, V 18834.1 is also similar to the specimens ofNyctereutes tingifrom Yushe of Shanxi in the following characteristics: 1) the dorsal profile of the parietal is relatively flat but not convex in lateral view; 2) the neurocranium is laterally inflated so poorly that its maximal width is almost equal to that across the postorbital processes; and 3) the postorbital constriction is so weak that its width is evidently greater than the distance between the orbits.

Table 2 Measurements of the crania of selected species in Nyctereutes (mm)
As a whole, nevertheless, the Yegou specimens seem closer toNycetreutes sinensis, with reservation of some primitive features similar toN. tingi. It should be noted herein that the cranium V 18834.1 is incomplete, and the analysis mentioned above is largely based on the reconstruction (see the white plaster part in Fig. 2). Whether the reconstruction is constructed correctly has a direct bearing on the conclusion of the similarity between V 18834.1 andN.tingi. In other words, its similarity toN. tingican not be assured at the moment. Under such circumstances, it seems more reasonable to assign the Yegou specimens described herein toN. sinensis. And if that’s the case, there would be occurrence of bothNyctereutesspecies at Yegou, Nihewan Basin, a similar case like in the Yushe Basin (Tedford and Qiu, 1991).
The morphological differences between V 18834.2–3 and V 18833.1 are as follows: 1)the ramus of the former is relatively slim, the anterior portion of which is especially shallow and the dorsal profile of which slopes signifcantly; 2) the m2 is relatively narrow, but the cingulid and entoconid are much better developed on the tooth. In these morphological characters, nevertheless, V 18834.2–3 exactly show the diagnostic features forN. sinensis.
In terms of the chronological distribution,Nyctereutessinensishas extensive Late Pliocene and Early Pleistocene records, such as those from the Yushe Basin of Shanxi(Tedford and Qiu, 1991), the Nihewan Basin of Hebei (Teilhard de Chardin and Piveteau,1930; Liu, 2019), Wushan of Chongqing (Huang and Zhong, 1991), Ningyang of Shandong(Zhang, 2001), Fanchang of Anhui (Liu and Qiu, 2009), Mianchi of Henan (Zdansky, 1924),Tuozidong, Nanjing of Jiangsu (Liu et al., 2007), and so on. In addition, someNyctereutesfossils from Middle Pleistocene horizons, such as Loc. 13 and Loc. 1 of Zhoukoudian,were also assigned toN.“sinensis” (Pei, 1934; Teilhard de Chardin and Pei, 1941), but the systemtic identification of theseNyctereutesfossils is still doubtful and disputed. In terms of morphology, they are actually closer to the extant species rather than the earlier fossil species(Soria and Aguirre, 1976; Tedford and Qiu, 1991).
The differences between the Yegou specimen V 18834.1 and the Zhoukoudian specimens ofN.“sinensis” lie in the following features: the frontal postorbital process of the former is more protruding; the postorbital constriction is not strong; the surface of the parietal is relatively smooth. All these features are basically consistent with theNyctereutesfossils from Nihewan and Tuozidong, Nanjing (Teilhard de Chardin and Piveteau, 1930; Liu et al.,2007; Liu, 2019). The morphology of V 18834.1 actually presents one more support for the judgement and analysis of Soia and Aguirre (1976), Tedford and Qiu (1991), and so on.
If the reconstruction of V 18834.1 is reliable, it would be slightly more primitive than the other known crania assigned toNyctereutessinensis(such as those from Nihewan and Tuozidong) with a flat parietal and less dilated neurocranium. In this respect, the specimens described herein seem to be no later than that from the Mazegou Formation of the Yushe Basin in age.

MaterialFragmentary ramus with p3–m1 (IVPP V 18835, Fig. 3).
MeasurementsSee Table 3.
DescriptionV 18835 is a left broken ramus with an absence of the portion anterior to the p3 and most of the ascending ramus. The horizontal ramus is relatively slim. The depth is 37.7 mm anterior to the p3, and 43.4 mm posterior to the m1, respectively. In occlusal view, the p3, p4 and m1 are almost rectangular in shape. The principal cusp of the p3 leans posteriorly and is massive but compressed buccolingually. The posterior accessory cusp is low. There is no discernible anterior accessory cusp developed, but a swollen enamel ridge is formed anteriolingually around the base of crown, probably representing a rudimentary cuspid.Feeble cingulids are discernible at the anterior and posterior margins of the tooth. The p4 is slightly larger than the p3. The principal cusp is robust and tall but leans posteriorly. It has anterior and posterior accessory cusps of equal size. The three cusps align almost in a straight line oblique to the long axis of the tooth, with the anterior accessory cusp positioned lingually and the posterior one buccally. The anterior cingulid is less developed than the posterior one.The anterior portion (where the paraconid is located) of the m1 is slightly broader than the posterior portion (the talonid). The paraconid is slightly longer than the protoconid, but they are almost equal in height. The metaconid is developed at the posterolingual corner of the protoconid, with only half the height of the latter. The two cusps are separated by a shallow groove. The talonid is robust and broad, and the trigonid is only 78% of the total length of the tooth. The hypoconid and entoconid are distinct. The hypoconid is almost displayed as a ridge parallel to the long axis of the tooth. The entoconid is low and conical on the lingual side.There is a curved ridge connecting the hypoconid and entoconid at the posterior margin of the talonid. Several cuspids or tubercles developed on it could be regarded as the rudimentary hypoconulid. A weak cingulid is developed on both the anterior and posterior margins of the tooth, respectively.
Comparisons and discussion In terms of dental size and morphology, the Yegou specimen described above (especially the m1) is conspicuously closest toPachycrocutain the Hyaenidae. It could be distinguished fromP. brevirostrisandP. licentiby its smaller size, relatively narrow premolars and an m1 with the more developed metaconid and talonid.The Yegou specimen virtually approximatesP. pyrenaicaandP. perrieriin both size and morphology.
In dental length, the Yegou specimen is nearly the same asPachycrocuta pyrenaicaandP. perrieri, and completely falls in the variation range of these two species (Table 3), but in dental width, the Yegou specimen is closer toP. pyrenaicafrom Yushe of Shanxi, and distinctly narrower thanP. perrieri(distinctly less than the mean of it, see Table 3). Morphologically,P.pyrenaicais different fromP. perrieriin its much narrower premolars (mainly P2/p2, P3/p3)(Howell and Petter, 1980; Qiu, 1987).
It seems not convincing enough to identify a species solely on the basis of tooth size.The lower carnassial of V 18835 would provide extra robust evidence for that.Pachycrocuta pyrenaicais relatively primitive in shape: the shearing blades of the carnassials (the P4 and m1) are relatively short. It is shown as a lower value in the proportion of the trigonid to the total length of the tooth on the m1 (i.e., Tri/m1 in Table 3) (Howell and Petter, 1980).The trigonid of the Yegou specimen is relatively short (about 78% of the total length) and comparable toP. pyrenaicafrom other localities, but shorter thanP. perrieri(completely out ofthe variation range of the latter, see Table 3). Moreover, the metaconid of the m1 usually is reduced or vanishes inP. perrieri(Howell and Petter, 1980; Qiu, 1987). For example, the metaconid on the m1 of JNTZ 6401 from Tuozidong, Nanjing is vestigial in the shape of an enamel tubercle on the posterolateral side of the protoconid near its base (Liu et al., 2007). The metaconid on the m1 of the Yegou specimen is small,but it is completely separated from the protoconid by a distinct shallow notch and acting as an independent cusp (see Fig.3B, C), which is quite differtent from the vestigial metaconid on the m1 ofP. perrieri.

Table 3 Measurements of the lower teeth of selected species in Pachycrocuta (mm)
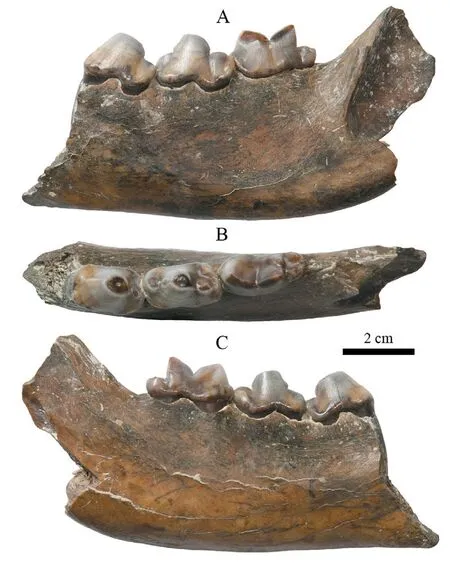
Fig. 3 Fragmentary right ramus of Pachycrocuta pyrenaica with p3–m1 (IVPP V 18835) from Yegou A. buccal view, B. occlusal view, C. lingual view
Pachycrocuta pyrenaicawas first discovered at Serrat d’en Vacquer, France.Depéret (1890) originally named it asHyaena arvernensisvar.pyrenaica. However,“Hyaena arverensis” should be a junior synonym of “Hyaena perrieri” (Howell and Petter, 1980; Qiu, 1987). In fact, as early as in 1954, Viret already clarified the situation,but emphasized that the specimens from Serrat d’en Vacquer, France, didn’t belong toperrieriat all, but represented another independent species “Hyaena donnezani” (Viret, 1954). Howell and Petter (1980) attributed the authorship of the species name to Charles Depéret based on the principle of priority. They also pointed out that the species didn’t belong to the extant genusHyaenaBrisson, 1762, but to the fossil genusPachycrocutaKretzoi, 1938. The name of the species was consequently revised asP. pyrenaica(Depéret, 1890).
Regarding size and morphology, certain similarities do exist betweenPachycrocuta pyrenaicaandP. perrieri. Sometimes it is difficult to distinguish them from each other. As a result,P. pyrenaicawas even questioned to be a probable synonym ofP. perrieri(Werdelin and Solounias, 1991; Turner et al., 2008). The Yegou specimen V 18835 bears the typical morphology ofP. pyrenaica(based on the standard of Howell and Petter, 1980), which provides powerful surport for the validity of the species. If only the dental length is taken into consideration, it would be difficult more or less to differentiate the Yegou specimen from small-sizedP. perrieri. For example, taking the Yegou specimen V 18835 asP. pyrenaicaand the Tuozidong specimen JNTZ 6401 from Nanjing asP. perrieri, the length of the p3, p4 and m1 would be 21.2, 22.5, 25.0 and 20.8, 23.0, 25.8 mm, respectively, while the corresponding width would be 13.2, 12.6, 12.0 and 14.5, 14.4, 13.6 mm, respectively. It is easy to notice that the p4 and m1 of V 18835 are slightly shorter than that of JNTZ 6401, but on the contrary, its p3 is longer than that of the latter. Nevertheless, the teeth of V 18835 are distinctly narrower than those of JNTZ 6401 (data for the latter from Liu et al., 2007). In addition, JNTZ 6401 could also be distinguished from V 18835 in m1 in the following characteristics: the relatively long trigonid (21.6 mm, 83.7% of the total length); the short talonid; and the reduced metaconid as a dependent tubercle. The present authors have no intention of discussing in depth the validity ofP. pyrenaica, which is beyond the scope of the present paper, but the Yegou specimen of V 18835 would provide a good example to support the conclusion of Viret(1954) and Howell and Petter (1980).
In terms of morphology,Pachycrocuta pyrenaicais undoubtedly more primitive thanP.perrieri, which is probably derived from the former (Schütt, 1972; Howell and Petter, 1980;Qiu, 1987). Their geochronological distribution also supports the following hypothesis:P.pyrenaicawas only found in the strata before Villafranchian in Europe, such as Serrat d’en Vacquer of France (Depéret, 1890), Layna (Crusafont and Sondaar, 1971) and La Calera II(Adrover et al., 1976) of Spain, and Odessa Catacomb of Ukraine (Howell and Petter, 1980),whileP. perrierioccured slightly later in the strata of Villafranchian and afterwards, such as Etouaires (Croizet and Jobert, 1828) and Saint-Vailler (Viret, 1954) of France, Villaroya and Puebla de Valverde of Spain (Croizet and Jobert, 1828), ValdArno of Italy (Weithofer, 1889),Hajnáčka of Slovakia (Fejfar, 1964) and so on. Fossil records ofP. pyrenaicaare relatively scarce in China, only known from Yushe of Shanxi (Qiu, 1987) and Zanda of Xizang (Tseng et al., 2016, =P. perrieri). In contrast, records ofP. perrieriare more extensive. It has been known from Yushe of Shanxi and the Nihewan Basin of Hebei (Qiu, 1987), Ningyang of Shandong(Zhang, 2001), Tuozidong, Nanjing of Jiangsu (Liu et al., 2007), Wushan of Chongqing(Huang and Zhong, 1991, =P. licenti; Zhang, 2001), and Shanyangzhai, Qinhuangdao of Hebei(Liu, unpublished). The geochronological distribution of the twoPachycrocutaspecies in China is similar to that in Europe. Except in the Yushe Basin,P. perrieriis only distributed in the Early Pleistocene (equivalent to middle and late Villafranchian). AlthoughP. perrieriandP. pyrenaicaco-occurred in the Yushe Basin, the former is only known from the upper part of the Mazegou Formation, the age of which is equivalent to the early Villafranchian,whileP. pyrenaicais known from the lower part of the Mazegou Formation, the Gaozhuang Formation, and even the Mahui Formation, the age of which is equivalent to or even older than the Ruscinian (Qiu, 1987; Deng and Hou, 2011). The discovery ofP. pyrenaicaat Yegou,the Nihewan Basin not only expands the distribution of this species but also provides reliable evidence to determine the age of the Yegou fauna.

MaterialAnterior half of left ramus with the i2, c, and p4 (IVPP V 18836.1, Fig. 4);fragmentary right ramus with i3 (V 18836.2).
MeasurementsSee Table 4.
DescriptionV 18836.1 is the anterior half of a left ramus with the portion posterior to the p4 missing. The symphysis is well preserved with the portion beneath the canine slightly broken. In rostral view, the symphysis is nearly rectangular with a height greater than the width. Multiple nutrient foramina could be discerned on the surface. In lingual view, the symphysis has a nearly vertical rough surface. The upper part is slightly wider than the lower.The posterior margin terminates approximately at the posterior edge of the canine (Fig. 4C). In buccal view, the mesial portion of the symphysis, especially the upper third, distinctly protrudes anteriorly so much that the incisor row is conspicuously anterior to the canine (Fig. 4A). The ventral margin of the symphysis is at about the same level as that of the horizontal ramus due to the missing mandibular flange, but the dorsal margin of the symphysis is apparently higher than that of the horizontal ramus, so the implant positions for the incisor and canine are higher than the cheek teeth (Fig. 4A, C). The robust ridge-like mental crest is developed beneath the canine, but most of the ventral portion is missing, and the mandibular flange is missing as well.However, judging from the surrounding structures, the mandibular flange might be very well developed. The p1 and p2 are not developed at all, so there is a diastema of 38.1 mm between the canine and p3. The dorsal margin of the ramus within the diastema slopes up anteriorly from the anterior end of the p3. Approximately beneath the midpoint of the diastema, a mental foramen of 10.7 mm in diameter is developed right above the ventral margin of the ramus.There is another mental foramen of 6.6 mm in diameter posterodorsal to the anterior mental foramen. The distance between them is 14.0 mm. Beneath the smaller mental foramen, there is an anteroventrally running ridge, which indicates the existence of a mandibular flange (although not preserved). In occlusal view, the dorsal edge of the diastema forms a sharp crest that is lingually concave (Fig. 4B).
In occlusal view, the incisors and the canine are densely aligned and form a unitary anteriorly convex arc (Fig. 4B). The i1 and i3 have already fallen off. Only the oval alveoli are preserved. Judging from the shape of the alveoli, i1 to i3 are gradually increasing in size.The i2 is conical and slightly bends posteriorly. There are two distinct crests developed on the anteromedial and posterodistal sides respectively. Both crests are serrated, but the posterodistal one is sharp. Although the p3 has shed off, the gourd-shaped alveolous (length/width: 12.9 mm/7.4 mm) clearly indicates that the tooth has two detached roots. The p4 is large and compressed buccolingually. In occlusal view, it appears approximately rectangular. The principal cusp is robust, tall and leans posteriorly. Two thin and sharp lateral crests run from the principal cusp forward and backward respectively. The anterior lateral crest is serrated.The two blunt-tipped accessory cusps are nearly equal in size and height. There is a second cuspid posterior to the posterior accessory cusp. A weak cingulid is developed only on the posterolingual margin of the tooth.
Comparisons and discussionAlthough the material is quite limited, a series of important anatomical features have been revealed: the mental crest and mandibular flange are developed at the symphysis; the incisor row is anteriorly convex; the canine, incisors and molars have lateral crests with serration; the implant position of the incisors and canine is higher than that of the check teeth. By these convincing features, the Yegou specimens described above should be undoubtedly assigned to the sabre-tooth group, namely the Machairodontinae. Based on the size and the morphology of the mandibular flange and the mental crest, they should definitely be excluded fromMegantereon. Consequently, onlyMachairodusandHomotheriumare applicable herein for futher discussion about its assignment.
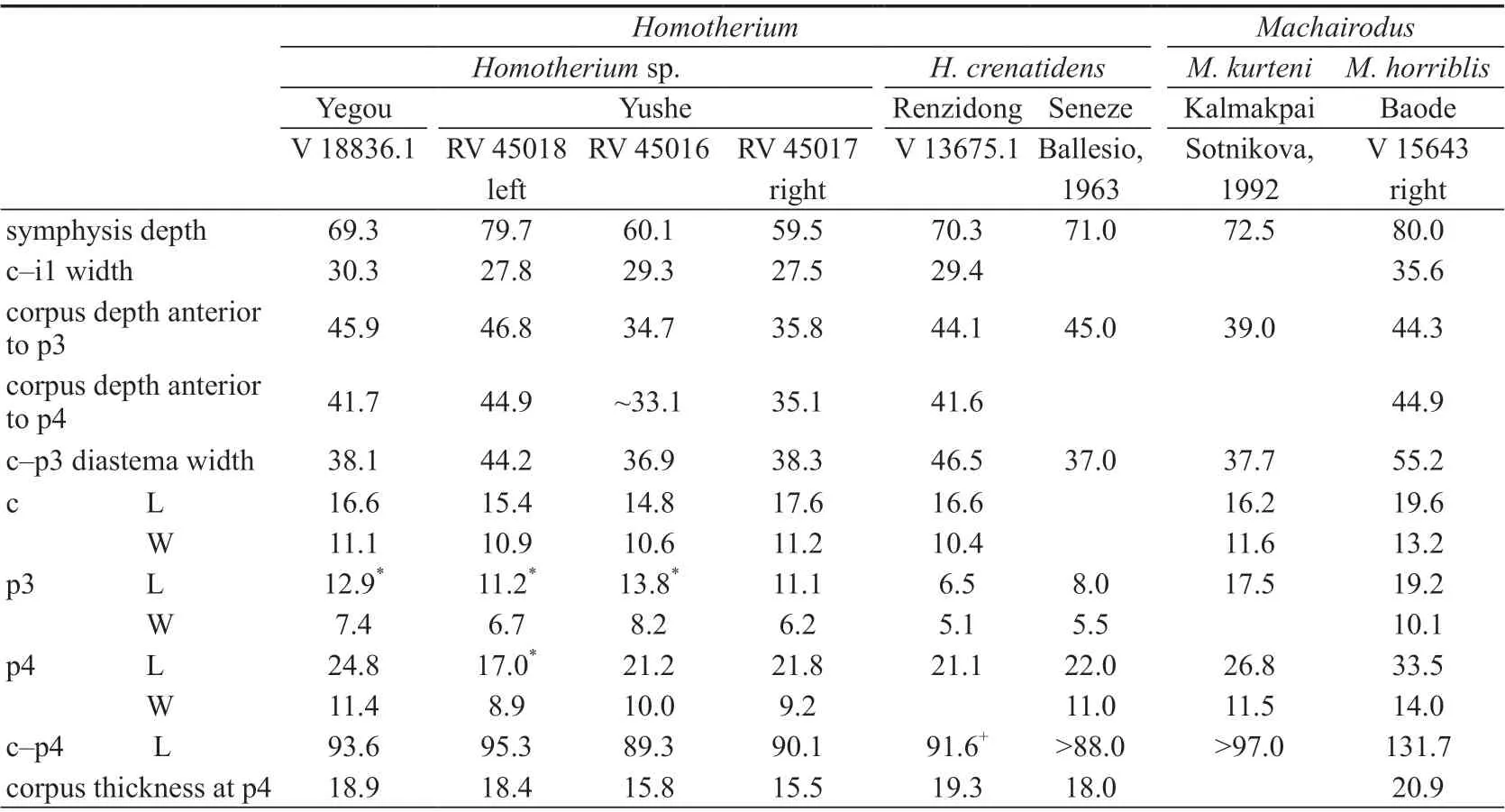
Table 4 Measurements of the lower teeth and mandibles of selected machairodontines (mm)
It is generally accepted thatMachairodusandHomotheriumare phylogenetically close to each other with the latter probably derived from the former (de Beaumont, 1975, 1978; Kurtén and Anderson, 1980; Sharapov, 1989; Sotnikova, 1992; Liu, 2003). It is not troublesome to distinguish the two genera when the specimens are plentiful and complete. However, it is rather difficult when the material is scarce and fragmentary. Teilhard de Chardin and Leroy(1945) once assigned the 3 mandibles from Yushe (RV 45016–45018 in Table 4, Teilhard de Chardin and Leroy, 1945: fig. 6) toMachairoduspalanderi(=Epimachairoduspalanderi).Judging from the current knowledge, there seem to be at least two differences between these mandibles of Yushe and other mandibles identified asMachairodus: 1) the Yushe specimens are smaller, especially RV 45016 and RV 45017 (see measurements in row 1, 3, 4, 9, 10 of Table 4), and even smaller thanM. kurteni; 2) the premolars of both the large individual (RV 45018) and the slender individuals (RV 45016 and RV 45017) are smaller than the species ofMachairodus, especially the p3. InMachairodus, the p3 is generally large. Even inM.palaterithat has a relatively reduced p3, its length can still reach 15.1 mm (Zdansky, 1924);the p3 usually has bifurcated two roots (Sharapov, 1989; Sotnikova, 1992:fig. 4); there are 2–3 cusps developed on the p3 (Sotnikova, 1992; Anton et al., 2004; Qiu et al., 2008). Among the mandibles from Yushe, the p3s of RV 45016 and RV 45018 are single-rooted; the p3 of RV 45017 is likely to be two-rooted, but they are fused together at least at the upper part; the p3s of RV 45016 and RV 45018 have fallen off, and only the alveolous is preserved; the p3 of RV 45017 is preserved, but it has only one cusp. It can thus be concluded that compared with the Miocene species ofMachairodus, the p3 of the Yushe specimen has been greatly reduced, so the assignment of them toMachairodusis not convincing.

Fig. 4 Fragmentary left mandible of Homotherium sp. with i2, c and p4 (IVPP V 18836.1) from Yegou A. buccal view, B. occlusal view, C. lingual view
Sotnikova (1992) pointed out the differences betweenMachairodusandHomotheriumin the implant and arrangement of their incisors and canine when studying the specimens ofM. kurtenifrom Kazakhstan. InHomotherium, the incisors and canine are densely aligned, so there is no gap between the central incisor and the canine; the incisors are upright and their tips are dorsally pointing. InMachairodus, there is a small gap between the canine and the incisors;the incisors are implanted obliquely with the tips pointing anterodorsally. Judging from these differences, the Yushe specimens are apparently consistent withHomotherium, but distinct fromMachairodus(see Teilhard de Chardin and Leroy, 1945:fig. 6).
Nonetheless, the Yushe specimens are not completely the same asHomotherium. RV 45018 is relatively robust. Its mandible is comparable to that ofHomotheriumcrenatidensfrom Renzidong, Anhui and Seneze, France, while RV 45016 and RV 45017 are distinctly slenderer. Moreover, their p3 is relatively developed and markedly larger than those later species inHomotherium(see Table 4). How to identify the Yushe specimens is obviously beyond the scope of this paper, but at least two conclusions could be drawn herein from above comparisons: 1) they should be assigned toHomotheriumrather thanMachairodus; 2) they probably represent a relatively primitive form in theHomotheriumclade, which is consistent with the age of the stratum they came from.
Although the assignment of the Yushe specimens toHomotheriumis not confirmed at the moment, the Yegou specimen shows some unambivalent morphological features ofHomotherium. The incisors and the canine are densely aligned with no gap between them.The incisors are upright and pointing dorsally. The incisors and the canine are so aligned that they form an anteriorly convex arc with the mesial ones more anteriorly implanted. There are serrations on all the preserved teeth. The last two features are more developed than the species ofMachairodus. Moreover, the size and robustness of the mandible are completely consistent with that ofH. crenatidensfrom Renzidong, Anhui (see Table 4). The only difference is its bigger premolars.
Presently, the taxonomical significance of the size of premolars (mainly the P3 and p3) amongHomotheriumspecies is still disputed. As a highly specialized felid, the anterior premolars ofHomotheriumprobably have lost their physiological function. Under such circumstances, the size and morphology of those nonfuctional premolars would change significantly. For that reason, it is highly dangerous to distinguish species based on these changes (e.g., Ficcarelli, 1979).
However, judging from the general evolutionary trend ofHomotherium, the gradual degeneration of the premolars is an indisputable fact. Their morphology (e.g., cusp and root) and size virtually reflect the evolutionary stage of the species.Homotheriummight consequently be divided into several different evolutionary stages or different species (e.g.,Sharapov, 1989). To that end, the authors observed and measured some specimens (see Table 4)and found that the evolutionary stages ofHomotheriumcould indeed be established roughly.For the species of the middle Villafranchian and afterwards (e.g., Renzidong, Anhui and Seneze, France), the length of the p3 generally does not exceed 10.0 mm. The p3 ofH. cuiifrom Middle Pleistocene Jinniushan is completely lost (Zhang et al., 1993). For the species of and before the early Villafranchian (e.g., Yushe, Shanxi), the length of the p3 is generally no less than 10.0 mm. The length of the p3 alveolous of the Yegou specimen is 12.9 mm and comparable to RV 45016 from Yushe. It is much larger than the species of the middle Villafranchian and afterwards. Based on the measurements in Table 4, the Yegou specimen is situated somewhere in betweenMachairoduskurtenifrom Kazakhstan andH. crenatidensfrom Renzidong of Anhui, and most likely is a transitional form between them (relatively speaking, the Yushe specimens are not good enough to represent such transitional form: the teeth of RV 45018 are somewhat distorted while the mandibles of RV 45016 and RV 45017 are so slender that they probably represent a different phylogenetic clade). It could be inferred herein that the Yegou specimens probably represent an early form ofHomotheriumor an earliest representative ofH. crenatidens(because the morphology and size of the mandible V 18836.1 are highly consistent with the specimens from Renzidong of Anhui). Unfortunately, V 18836.1 is the only identifiable specimen collected at Yegou. It is impossible to evaluate and analyse the variation of the p3 comprehensively, and the possibility of the big-sized p3 of V 18836.1 as an individual variation can not be ruled out at the moment. Although the premolars ofHomotheriumgenerally tend to be reduced, some individuals with extreme variation did occur time and again. For example,H. crenatidensfrom Val d’Arno of Italy have reduced premolars, but there is still one p3 with a length of 12.0 mm (Ficcarelli, 1979), comparable to the Yegou specimen. If only based on such an isolated p3, it would be less credible to presume that the Yegou specimen as an early primitive form ofHomotherium. Nonetheless, if the associated animals, such asNyctereutes tingiandPachycrocuta pyrenaica, are taken into account, the present authors would be more confident of such a presumption, and consequently identify the Yegou specimens herein asHomotheriumsp. for the moment. Further identification and inference still await more and better material in the future to be clarified.

Material Fragmentary right maxilla with the P3–M2 (IVPP V 18837.1, Fig. 5C) and left and right mandibular corpora with the p2–m3 (V 18837.2, V 18837.3, Fig. 5A) of the same individual, fragmentary right maxilla with the M1–3 (V 18837.4), 1 left and 1 right P2 (V 18837.5–6), fragmentary right mandibular corpus with the p2–m3 (V 18837.7, Fig. 5B), distal portions of two right humeri (V 18837.8, Fig. 6D; V 18837.9), proximal portion and distal portion of the same left Mc III (V 18837.10, Fig. 6A; V 18837.11), proximal portion of the left Mc IV (V 18837.12), right talus (V 18837.13, Fig. 6E), right calcaneus (V 18837.14, Fig. 6G),proximal portion of the right Mt III (V 18837.15, Fig. 6B), proximal portion of the right Mt II (V 18837.16, Fig. 6C), and the ungual phalanx of central digit (V 18837.17, Fig. 6F).
27.Geese: Geese are associated with the earth mother, maternity80, fertility, truth, love, constancy, vigilance, providence81, silliness, stupidity, female sexuality, wind, innocence, cowardice82, and the good housewife (Olderr 1986, 58)Return to place in story.
Measurements See Tables 5–12.
Description IVPP V 18837.1 is a fragmentary maxilla with the P3–M2. The facial crest terminates above the anterior margin of the M1. The preorbital fossa is located above the P3,and the foramen fossa is above the P4. In occlusal view, the center of the posterior margin of the hard palate is approximately level with the posterior portion of the M2 (Fig. 5C1). On V 18837.4, the major palatine foramen is visible and level with the posterior portion of the M2,whereas the minor palatine foramina are not discernable.
On V 18837.1, the size of the upper cheek teeth gradually decreases from the P3 to M2(Fig. 5C1). The parastyle is only broadened on the P3–M1, where it is bifid and made up oftwo ribs and one median cleft. The anterior rib and median cleft are not developed on the M2.The size of the median cleft gradually decreases from the P3 to M1. The mesostyle is only slightly elongated on the P3–M1, but not on the M2. The labial wall of the paracone and the metacone is labially concave, and their lingual walls are lingually convex. The angle between the anterior lingual and posterior lingual walls tend to become smaller from the P3 to M2, but all less than 90°. The anterolabial corner of the postfossette extends more labially than the posterolabial corner of the prefossette on the cheek teeth. The hypoconal groove is shallow,and the hypoconal constriction is distinct. The counting of enamel plications in the form of“anterior wall of the prefossette.posterior wall of prefossette-anterior wall of postfossette.posterior wall of postfossette/pli caballin” is as follows: P3 = 6.10-7.2/4, P4 = 6.8-6.2/3, M1 =6.7-6.1/1, M2 = ?-3.1/1. The protocone length index (= protocone length/tooth length×100) is as follows: P3 = 23.8, P4 = 29.2, M1 = 35.0, M2 = 27.3. The protocone width/length index (=width/length×100) is as follows: P3 = 70.8, P4 = 64.4, M1 = 61.3, M2 = 70.0.
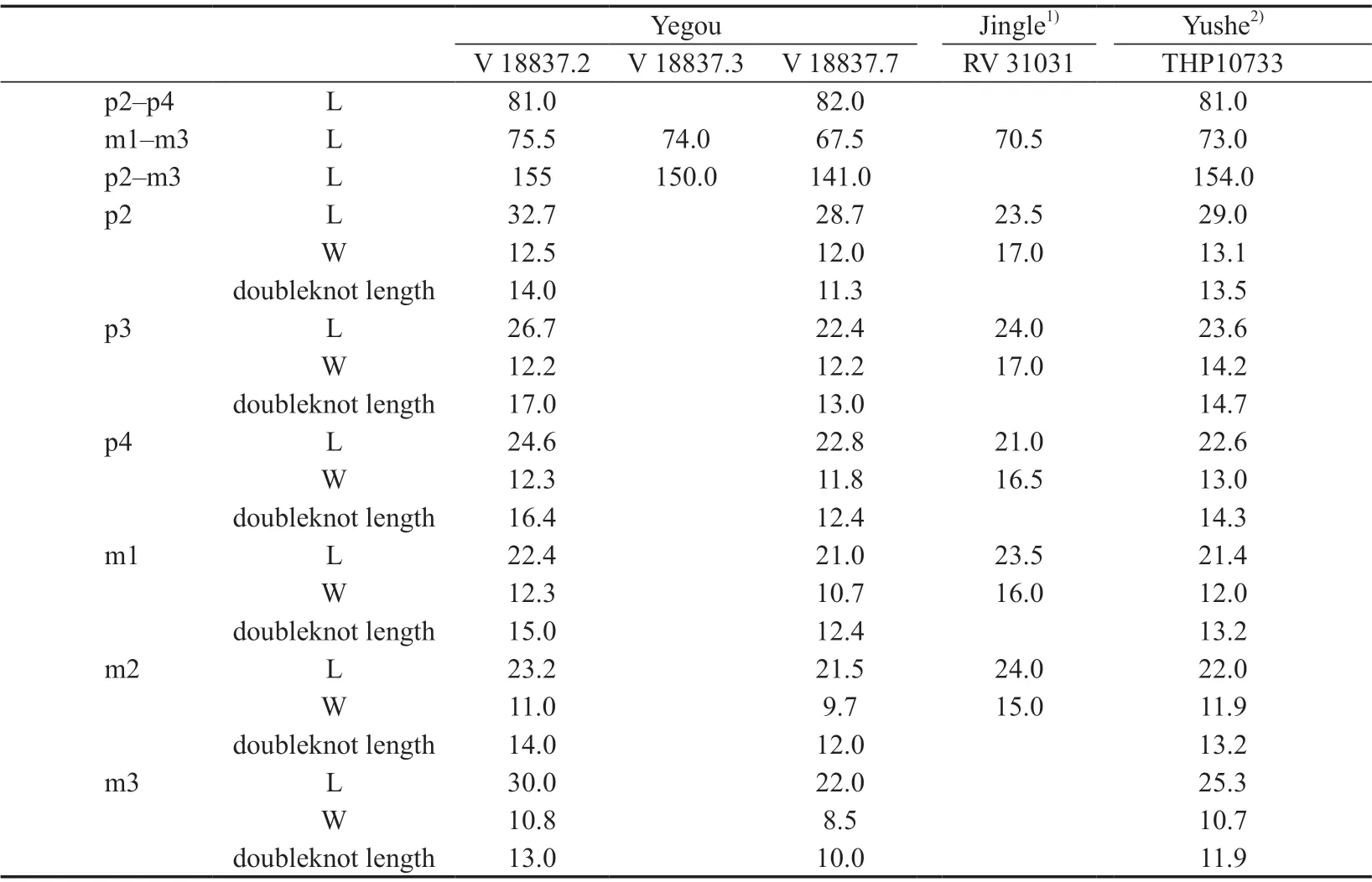
Table 5 Mesurements of the lower cheek teeth of Hipparion (Plesiohipparion) huofenense (mm)

Table 6 Measurements of the upper cheek teeth of Hipparion (Plesiohipparion) houfenense (mm)

Fig. 5 Fragmentary maxilla and mandibular corpora of Hipparion (Plesiohipparion) houfenense from Yegou A. right mandibular corpus with p2–m3 (IVPP V 18837.3); B. right mandibular corpus with p2–m3(V 18837.7); C. right maxilla with P3–M2 (V 18837.1); 1. occlusal view, 2. buccal view
The size of V 18837.4 is small. The major palatine foramen lies between the M2 and M3.The parastyle is elongated on the M1 but not bifid. It is not elongated on either the M2 or the M3.Its extent of labial protruding gradually decreases from the M1 to M3. The hypoconal groove is shallow, and the hypoconal constriction is distinct. The counting of enamel plications is as follows:M1 = 6.8–3.1/3, M2 = 6.9–3.2/2, M3 = 5.8–4.2/2. The protocone length index: M1 = 34.1, M2 =31.8, M3 = 19.7. The protocone width/length index: M1 = 69.7, M2 = 68.6, M3 = 57.6.
V 18837.5 and V 18837.6 are the left and right P2, respectively. The anterostyle is not inflated. The size of the hypoconal groove is comparable to that of the hypoconal constriction.The left P2 is slightly smaller. The mesostyle extends more labially. The protocone is relatively short and small. Its lingual margin is concave lingually. The protocone of the right P2 is long and robust. Its lingual margin is straight but anterolabially oblique. The counting of enamel plication of the two teeth is 4.2–3.1/1 and 3.4–4.2/2, respectively. The protocone length index:21.5 and 25.1. The protocone width/length index: 61.6 and 72.9.
Mandible and mandibular cheek teeth (Fig. 5A–B): The larger left and right mandibular corpora should belong to the same individual. Only the right horizontal ramus is relatively complete (V 18837.3, Fig. 5A). The depth beneath the p2 on the lingual side is 72.0 mm. The depth between the p4 and m1 is 88.5 mm. The depth beneath the m3 is 98.0 mm.
The p2 is slightly longer than the p3. The paraconid is robust. The anterior lobe of the double-knot (the metaconid) is rounded and about half the size of the posterior lobe (themetastylid). The double-knots of the p3 and p4 are triangular and equal in size. The doubleknots of m1–3 are also triangular, but the metaconids gradually become bigger than the metastylid from the m1 to m3. The isthmus is short on the p2–4 but distinct. It is not developed on m1–3. The lingual flexid is U-shaped. It is the shallowest on the p2, and gradually becomes deeper afterwards. The ectoflexid is also shallow on the p2, but gradually becomes deeper afterwards. The pli caballinid and the pli antecaballinid are distinct on p3–m3 of the smaller specimen V 18837.7 (Fig. 5B), whereas only the pli antecaballinid is distinct on the larger specimen V 18837.3 (Fig. 5A). There are no pli caballinid and pli antecaballinid developed on the p2 of either specimen. The protostylid is developed on both specimens, but the pli hypostylid and the ectostylid are not developed. On V 18837.7, the m3 is slightly longer than the m2, which is similar to the holotype RV 31031 from Jingle. The m3 of V 18837.3 is distinctly larger than the m2 and similar to THP10733 from Yushe.

Table 7 Measurements of the humerus of Hipparion (Plesiohipparion) houfenense (mm)

Table 8 Measurements of the metacarpal III of Hipparion (Plesiohipparion) houfenense (mm)
Humerus (V 18337.8, Fig. 6D): The lateral supracondylar crest extends posteriorly and distally to the posterior margin of the lateral ligament fossa, weakens medially, and terminates behind the articular surface of the lateral epicondyle. The distal breadth of the articular surface of the medial condyle (33.0 mm) is about twice of that (16.0 mm) of the lateral condyle. Thetrochlear depth on the medial condyle side (37.0 mm) is about 1.3 times of that (28.0 mm) on the lateral condyle side. The sagittal crest between the medial and lateral condyles is rounded anteriorly but sharp posteriorly and extends posterolaterally.

Table 9 Measurements of the calcaneus of Hipparion (Plesiohipparion) houfenense from Yegou (mm)

Table 10 Measurements of the talus of Hipparion (Plesiohipparion) houfenense (mm)
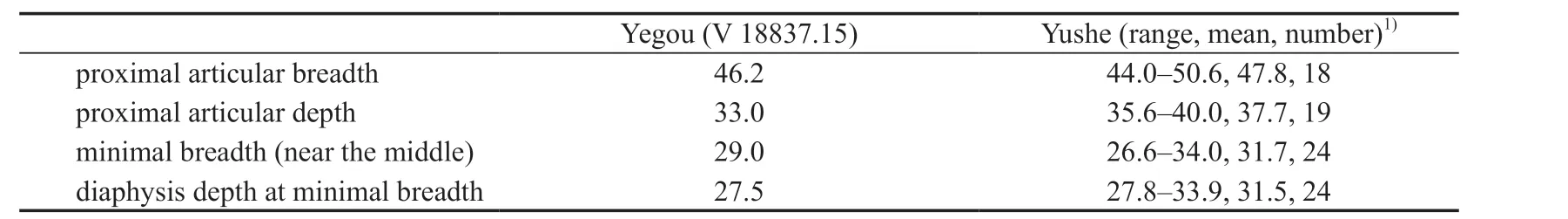
Table 11 Measurements of the metatarsal III of Hipparion (Plesiohipparion) houfenense (mm)
Mc II: The proximal half is preserved. The volar side of the diaphysis is concave. The depth of the diaphysis is greater than that of the contact surface with the Mc III. The proximal articular surface is flat, perpendicular to the diaphysis, and slightly higher than the proximal articular surface of the Mc III when it is in anatomical position. There are two separated articular facets with the Mc III on the medial side and one with the Mc I on the lateral side.
Talus (V 18837.13, Fig. 6E): The talus is asymmetrical. The lateral condyle is distinctlywider than the medial condyle, but the depth of the former is distinctly less than the latter. The slope of the medial wall of the lateral condyle is gentle. The dorsoplantar distance between its distal end and the distal articular facet is 10.0 mm. The medial wall of the medial condyle is nearly vertical and extends posteriorly about half of the depth of the body. The distal portion of medial wall does not bend medially. The depth at the median groove of the trochlea is about 70% of that of the lateral condyle. There are 4 articular facets with the calcaneus. The medial facet is a long strip in shape, with its long axis proximodistally oriented. The medial margin is convex, whereas the lateral margin is concave. It is 31.0 mm long and 13.5 mm wide. The lateroproximal facet is a transverse and anteriorly convex curved surface and it is mediodistally oriented. The proximolateral margin intersects with the lateral wall of the lateral condyle. The tounge-shaped facet or lateral facet is small, with a pointed proximal margin and a rounded distal margin. The proximodistal dimension is 9.0 mm, and the mediolateral dimension is 5.0 mm. The lateroproximal facet and the lateral facet are not connected with each other. The laterodistal facet is rectangular, anteriorly wider, and posteriorly narrower.The anteroposterior dimension is 10.0 mm, and the proximodistal dimension is 7.0 mm. The distal facet is triangular. A concave rough surface medially stretches into the facet from the middle part of its lateral margin and terminates behind the distal end of the medial condyle.The width of the distal facet is 45.5 mm, and the depth is 34.0 mm. The posterior pointed protrusion exists, but it is weak. The medial proximal protuberance is distinct and posterior to the proximal end of the medial condyle of the trochlea. It extends posteriorly about 6.0 mm.The distal protuberance is relatively strong and protrudes medially. The two protuberances are connected by a shallow ridge between them.
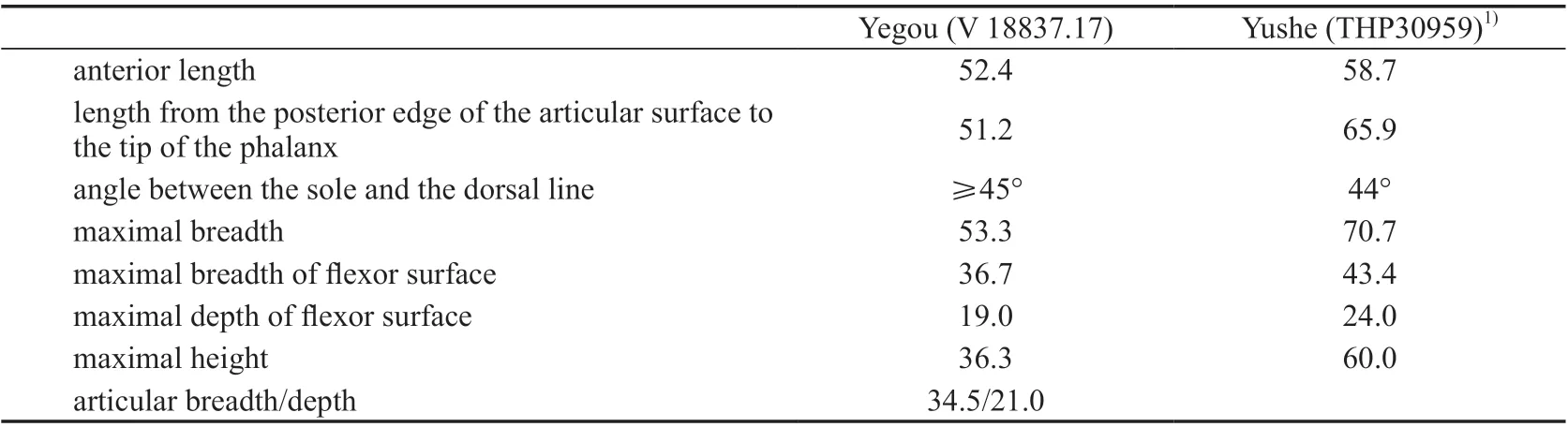
Table 12 Measurements of the ungual phalanx of Hipparion (Plesiohipparion) houfenense (mm)

Fig. 6 Limb bones of Hipparion (Plesiohipparion) houfenense from Yegou A. proximal portion of left Mc III (IVPP V 18837.10: 1. proximal view, 2. anterior view);B. proximal portion of right Mt III (V 18837.15: 1. proximal view, 2. anterior view);C. proximal portion of right Mt II (V 18837.16: anterior view);D. distal portion of right humerus (V 18837.8: posterior view); E. right talus (V 18837.13: anterior view);F. ungual phalanx of the central digit (V 18837.17: anterior view);G. right calcaneus (V 18837.14: 1. lateral view, 2. medial view)
Calcaneus (V 18837.14, Fig. 6G): The calcaneal tuberosity is damaged. The medial articular facet is a long strip in shape and slightly concave. Its posteromedial margin leans medially.Its length is 35.2 mm, and its width is 16.3 mm. This facet corresponds to the medial articular facet of the talus. The proximolateral articular facet is nearly circular in outline. It is divided into two parts perpendicular to each other by a transverse ridge. Its width is 22.3 mm, and its depth is 21.0 mm. The tongue-shaped or lateral articular facet is rectangular and connected with the proximolateral articular facet, with a width of 8.4 mm and a length of 16.2 mm. The distolateral articular facet is trapezoid in outline, and the distolateral margin is pointed with a dorsodistal dimension of 20.0 mm and a plantarodistal dimension of 7.2 mm. The distal articular facet is laterally wider and medially narrower. The plantaromedial third of the facet suddenly shrinks. In lateral view, the angle between the lateral margin of the distal articular facet and the dorsodistal margin beneath the tongue-shaped facet is acute.
Mt III (V 18837.15, Fig. 6B): Only the proximal portion is preserved. The depth is slightly less than the breath at the middle of the diaphysis. The plantar side is distinctly concave. The vascular impression is distinct on the lateral side of the proximal Mt III, which intersects with the dorsal margin of the diaphysis about 3 cm beneath the proximal articular facet at an angle of about 45°. Then it runs distally along the lateral margin of the Mt IV. There is a concave non-articular rough surface that extends anteromedially into the center of the proximal articular facet from the plantarolateral corner. The proximal articular surface consists of the medial articular facet with the small cuneiform (os tarsalia I+II), the middle articular facet with the big cuneiform (os tarsale III), and the lateral articular facet with the cuboid(os tarsale IV). The articular facet with the small cuneiform is the smallest among them. It is semi-circular and intersects with the facet with the big cuneiform at an angle larger than 90°.The articular facet with the big cuneiform is the largest and horseshoe-shaped. Both wings of the facet as well as the plantar and dorsal ends are prominent. The facet intersects with the articular facet with the cuboid at an angle of about 120°. The latter is relatively large, flat and rectangular. There are 2 isolated articular facets on the medial and lateral sides of the proximal articular surface, articulating with the Mt II and Mt IV respectively.
Ungual phalanx of the central digit (V 18837.17, Fig. 6F): The angle between the sole and the dorsal line is about 45°. The rough area for the insertion of the flexor digitorum profundus is distinct and large on the posterior surface where there is a protuberance in the center. The articular facet is semi-circular. The anterior margin is slightly convex, and the posterior margin is straight. It is made up of two concave surfaces: the lateral one and the medial one.
Comparisons and discussionHipparion houfenenseor its conformis species have been known from Jingle, Shanxi (Teilhard de Chardin and Young, 1931), Pucheng, Shaanxi(Jen, 1965, =Hipparioncf.huofenense), Huoxian, Shanxi (Tong et al., 1975, =Hipparioncf.huofenense), Yushe, Shanxi (Qiu et al., 1980), Weinan, Shaanxi (Xue, 1981), Yuxian,Hebei (Tang and Ji, 1983, =Hipparioncf.huofenense), Lingtai, Gansu (Zhang et al., 1999:p2–m3 length, 145.5 mm). Among them, the material from Huoxian has been excluded fromH. houfenense(Qiu et al., 1980); the material from Weinan has been assigned toH.(Proboscidipparion)pater(Qiu et al., 1987); the material from Pucheng has been assigned toH.(Plesiohipparion)huangheense(Qiu et al., 1987)
The smaller mandible from Yegou (V 18837.7) is close to the specimens from Jingle,whereas the larger mandibles (V 18837.2–3) are close to the Yushe specimens. The main difference between these forms lies in that m3 of the former lacks an extra posterior lobe. The size of the Yegou upper cheek teeth is comparable to the Yushe specimens, but the protocone length index is greater, which means the protocone of the former is more rounded than that of the latter. The Mc III, Mt III and the ungual phalanx from Yegou are all smaller than the Yushe specimens, and the humerus and the talus are also smaller than the specimens from Jingle and Yushe. Due to the limited sample size, the smaller dimensions probably do not indicate primitiveness but rather individual variations. For example, the specimens with greater protocone length index represent aged individuals, whereas the smaller limb bones represent young individuals.

MaterialProximal portion of the left Mc II (IVPP V 18838.1, Fig. 7D), distal portion of the left Mc III (V 18838.2, Fig. 7A), distal portions of 2 left Mt IIs (V 18838.3, Fig. 7B;V 18838.4), distal portions of 2 left Mt IVs (V 18828.5; V 18838.6, Fig. 7C), right patella(V 18838.7, Fig. 7E), distal portion of the left radius (V 18838.8, Fig. 7G), left calcaneus (V 18838.9, Fig. 7F), proximal portion of the left calcaneus (V 18838.10).
MeasurementsSee Table 13.
DescriptionIVPP V 18838.8 is the distal portion of a radius (Fig. 7G). The diaphysis is robust and anteroposteriorly compressed. The medial margin is flat and thick, while the lateral margin is compressed and thin. The anterior surface is slightly convex, and the posterior surface is slightly concave. The distal outline is triangular. There is a tuberosity with a rough surface on the lateral side, which contributes to the maximal distal breadth. The anterior portion of the distal articular facet is concave, whereas the posterior portion is convex. The distal articular facet is divided into the larger medial part and the smaller lateral part by a low ridge. The lateral part articulates with the ulnar carpal, and the medial part articulates with the radial carpal. The medial articular facet with the radial carpal is semi-circular and intersects with the articular facet with the ulnar carpal approximately at a right angle.
Only the proximal half of the Mc II is preserved (V 18838.1, Fig. 7D). The proximal outline is triangular. On the medial side, there is a triangular rough surface beneath the proximal articular surface. Posterior to this surface, there is a long bar-like protuberance.There is another protuberance medial to the surface. The articular facet with the trapezium (os carpale I) is the largest and it is semi-circular. It is concave in the middle and separated from the long strip-like and anteroposteriorly oriented articular facet with the trapezoid (os carpale II) by a sharp ridge. The articular facet with the Mc III is oval in shape with its major axis anteroposteriorly oriented. It is separated from the facet with the trapezoid by a proximally convex low ridge.
Only the distal half of the Mc III is preserved (V 18838.2, Fig. 7A). The diaphysis is broad and dorsovolarly compressed. The lateral side of the bone is rougher than its medial side.The proximal margin of the distal articular facet on the dorsal side is proximally convex. The sagittal crest is not seen on the dorsal side, but it is strong on the volar side where it extends proximally and connects to a large tuberosity.
The patella is complete (V 18838.7, Fig. 7E). It is a large sesamoid bone that is robust and rectangular in outline. The dorsal surface of the bone, for insertion of tendons, is convex and rough. The articular surface with the patellar surface of the femur is also rectangular. There is a broad and rounded ridge dividing the surface into the lateral and the medial parts.
Calcaneus: V 18838.9 is an incomplete calcaneus with only the tuberosity broken off(Fig. 7F). V 18838.10 preserves only the tuberosity and part of the body. Its body is stout.The tuberosity is robust, and its posterior surface is rough. Curved ridges can be seen on the mediodistal margin of the tuberosity. The lateral side of the body proximal to the cochlear process is slightly convex, but the medial side is slightly concave. Both sides are smooth. The articular surface with the talus is divided into two parts by the calcaneal groove. The lateral facet is circular and convex, and the medial facet is tadpole-shaped and concave. The calcaneal groove is deep and extends first distally then distolaterally beneath the lateral facet, where it becomes shallow and merges with the distal margin of the lateral facet. The distal articular facet with the cuboid (os tarsalia IV+V) is semi-circular, concave and intersects with the medial facet with the talus at an angle slightly larger than 90°.
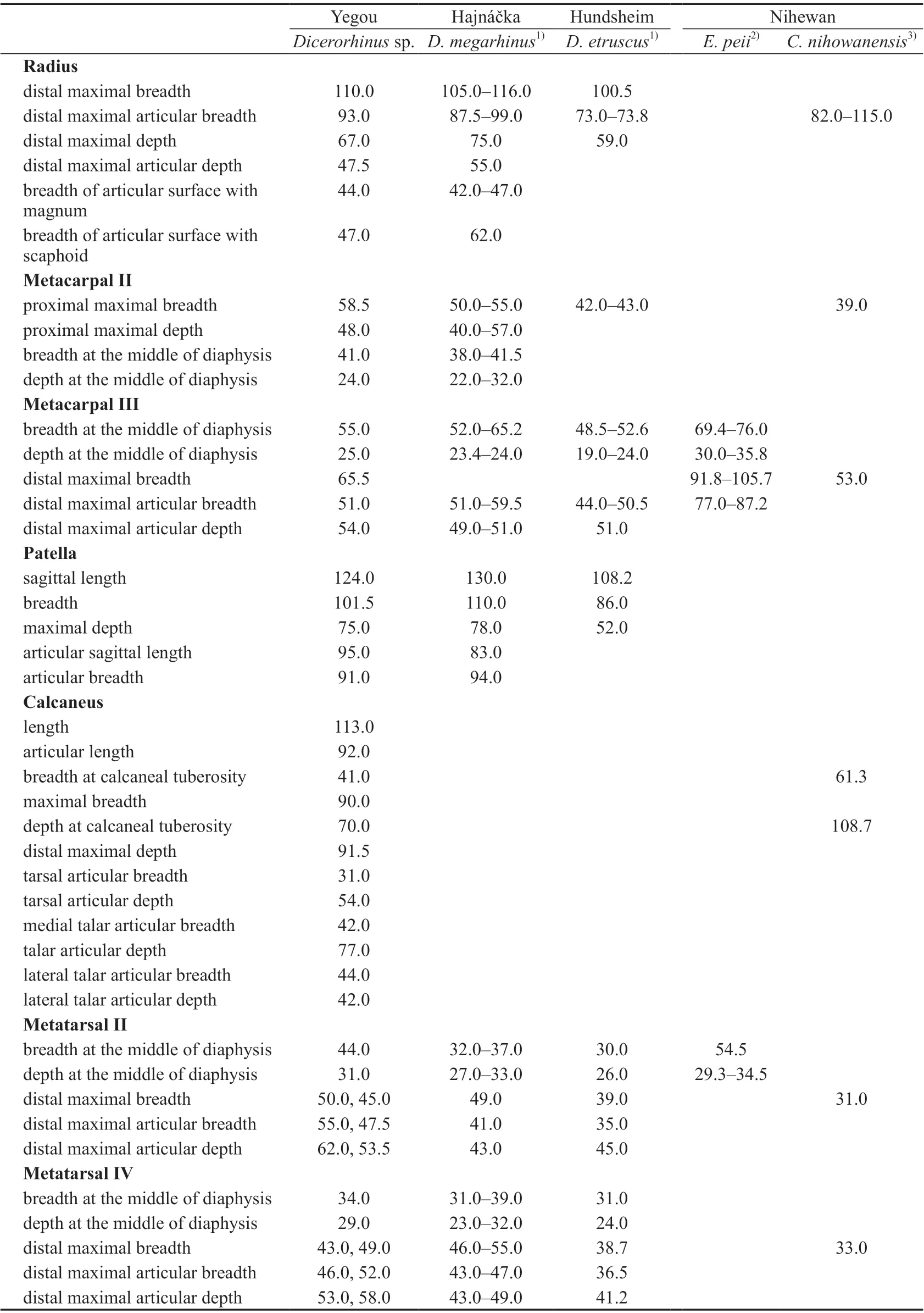
Table 13 Limb bone measurements of selected species in Dicerorhinus, Elasmotherium and Coelodonta (mm)
There are two Mt IIs, with one being larger than the other. Only the distal protion is preserved (V 18838.3, Fig. 7B). The depth of the diaphysis is larger than the breadth. The medial margin is relatively flat and thicker than the lateral margin.The medial surface is rough. The maximal breadth of the distal articular facet is at the midpoint of the depth. The sagittal crest is only prominent on the plantar side.The articular surface is higher than the diaphysis surface on the dorsal side of the larger specimen, and at the same level on the smaller specimen. The sagittal crest is robust and distinctly proximally exceeds the articular surface on the plantar side of the larger specimen, but only slightly on the smaller specimen. The difference in size and morpholoty probably reflects not only variation, but also interspecific distinction.
There are two Mt IVs as well. One is larger, and the other is smaller. Only the distal portion is preserved (V 18838.6, Fig.7C). The morphology is basically the same as that of the Mt II.

Fig. 7 Limb bones of Dicerorhinus sp. from Yegou A. distal portion of left Mc III (IVPP V 18838.2: anterior view); B. distal portion of left Mt II (V 18838.3: anterior view); C. distal portion of left Mt IV (V 18838.6: anterior view); D. proximal portion of left Mc II(V 18838.1: anterior view);E. right patella (V 18838.7: 1. anterior view,2. posterior view); F. left calcaneus (V 18838.9: anterior view); G. distal portion of left radius(V 18838.8: anterior view)
Comparisons and discussionThree rhinocerotid species have been known from the Early Pleistocene deposits of the Nihewan Basin, including the distinctly small-sizedDicerorhinusmercki(=Stephanorhinus kirchbergensis), medium-sizedCoelodonta nihowanensis, and large-sizedElasmotherium peii(Teilhard de Chardin and Piveteau, 1930;Deng and Zheng, 2005:Elasmotherium caucasicum; Tong et al., 2014). Among them, only the limb bones of the the latter two species have been unearthed (Teilhard de Chardin and Piveteau, 1930; Deng and Zheng, 2005). In terms of the measurements (Table 13), the sizes of the Mc III, calcaneus, and Mt II described here are distinctly smaller than the corresponding elements of Early PleistoceneElasmotherium peiifrom the same basin. The size of the radius is similar, but that of Mc II–III and Mt II, IV is distinctly greater than Early PleistoceneCoelodonta nihowanensisfrom the same basin. AlthoughDicerorhinusmerckiis also known from the Nihewan Basin, there are only limb bones from Yegou, which lack decisive diagnostic morphological characters. Consequently, the new material is temporarily identified asDicerorhinussp.
A Mc II that is close to the Yegou specimen is known from the red clay layer of Renjiagou,Lingtai, Gansu (Zhang et al., 1999: Rhinocerotidae gen. et sp. indet.). The proximal length/width is 55.0 mm/41.0 mm, and the minimal breath/depth of the diaphysis is 39.5 mm/23.3 mm. The magnetostratigraphic age of the horizon is 3.5–3.4 Ma (Sun et al., 1998; Zhang et al., 1999).
The size of the Yegou specimen is also very close to that of theDicerorhinusmegarhinus(Christol, 1835) from Hajnáčka, southern Slovakia (Fejfar, 1964). Based on the coexistingMimomys hajnackensisFejfar, 1961, its geological age should be Early Villanyian, > 3 Ma(Repenning et al., 1990).

MaterialLeft horn core with the pedicle and partial frontal bone but without the tip(IVPP V 18839.1, Fig. 8B), left horn core without the pedicle but with the tip slightly damaged(V 18839.2), 1 left and 1 right fragmentary mandibular corpora with the p2–m3 (V 18839.3; V 18839.4, Fig. 8A), fragmentary right mandibular corpus with the p2 alveoli and the p3–m3 (V 18839.5), fragmentary right mandibular corpus with the p3–p4 (V 18839.6), fragmentary right mandibular corpus with the m3 (V 18839.7).
MeasurementsSee Table 14.
DescriptionV 18839.1 (Fig. 8B) is a fragmentary horn core with the dorsal half missing and a small portion of frontal bone preserved at the pedicle. The supraorbital foramen can be seen anterior to the pedicle of the core. The postcornual fossa is deep and oval on the lateral side of the pedicle. The horn core slightly bends caudally. There are coarse and parallel longitudinal ridges (or grooves). Judging from the suture preserved on the medial side, the minimal distance between the pedicles is about 2.4 cm. The horn core extends caudolaterally.The other specimen does not have the frontal bone preserved, and the tip is slightly damaged (V 18839.2). Its curvature is similar to that of V 18839.1. Although the rostral surface is damaged,it seems that the longitudinal ridges (or grooves) are not as coarse as that of V 18839.1.
All mandibular specimens are incomplete. The length of p2–p4 is 38% of that of the lower cheek teeth row, but greater than that of m2–m3. The p2 is single lobed with a pointed protoconid and two roots. The p3 and p4 both have anterior, middle, and posterior valleys. The middle and posterior valleys can both reach the labial side. The metaconid and the paraconid are not fused completely on the p4. The protoconid is pointed. The morphology of lower molars is similar to that of other gazelles, but the goat fold is present anterolaterally in young individuals.
Comparisons and discussionThe material described in this paper is slightly larger thanGazella blackifrom Xiaohongao, Hefeng and Renjiagou, Lingtai, and distinctly smaller thanG.sinensisfrom the Nihewan Basin (Table 14). The average ratios of the p2–p4 length to the p2–m3 length in percentage for these three forms are 38.2% (Xiaohongao), 36.2% (Renjiagou), and 35.3%(Nihewan) respectively. The material here is therefore closer toG. blackithan toG. sinensis.
When studying the gazelle fossils from North China, especially Yushe, Chen (1997a,b)pointed out thatGazella blackiis a valid species, and it is a typical form for the Late Pliocene of North China. The magnetostratigraphic age provided by Chen (1994) is ca. 3 Ma.

Fig. 8 Selected dentition and horn core of Gazella blacki from Yegou A. fragmentary right mandibular corpus with p2–m3 (IVPP V 18839.4: 1. buccal view,2. occlusal view; 3 cm scale bar);B. fragmentary left horn core (V 18839.1: 1. lateral view, 2. anterior view, 3. medial view; 2 cm scale bar)
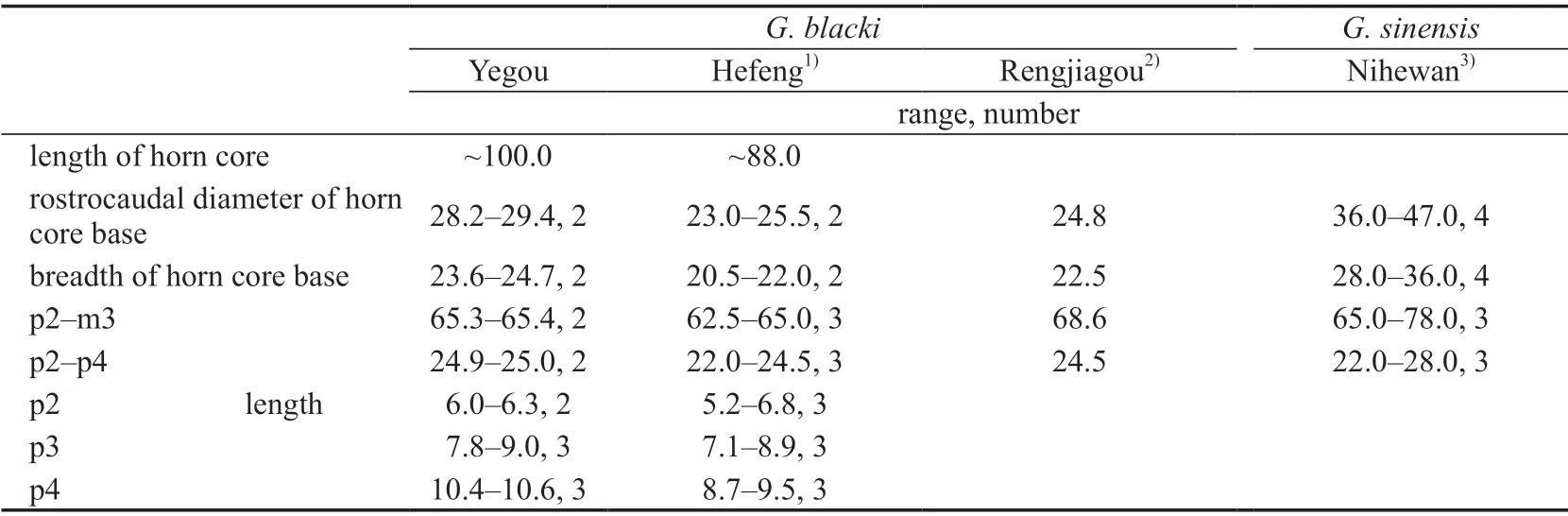
Table 14 Measurements of the horn core and cheek teeth of Gazella blacki and G. sinensis (mm)
Gazellacf.G. blackialso occurs in many Early Pleistocene faunas, such as Wenxi,Houma, (Tang, 1980a), Xihoudu, Ruicheng (Chia and Wang, 1978), Xicun, Tunliu, Shanxi(Zong et al., 1982), and Longdan, Dongxiang, Gansu (Qiu et al., 2004). In this regard,G.blackiis probably a long-lasting species spanning from the Late Pliocene to Early Pleistocene.

Material Antler fragment (IVPP V 18841.1, Fig. 9A), fragmentary right maxilla with the P2–M3 (V 18841.2, Fig. 9B), the right P3 (V 18841.3), distal portion of the right radius(V 18841.4, Fig. 9C), distal portion of the left tibia (V 18841.5, Fig. 9D), 1 left and 1 right tali(V 18841.6, Fig. 9E; V 18841.7), proximal portions of 2 Mt III-IVs (V 18841.8, Fig. 9F; V 18841.9).
Measurements See Table 15.

Fig. 9 Antler, upper cheek teeth, and limb bones of Axis shansius from Yegou A. antler fragment (IVPP V 18841.1: 1. lateral view, 2. medial view); B. fragmentary right maxilla with P2–M3 (V 18841.2: occlusal view); C. distal portion of right radius (V 18841.4: 1. anterior view, 2. distal view);D. distal portion of left tibia (V 18841.5: 1. posterior view, 2. distal view);E. left talus (V 18841.6: posterior view); F. proximal portion of left Mt III–IV(V 18841.8: 1. anterior view, 2. proximal view)
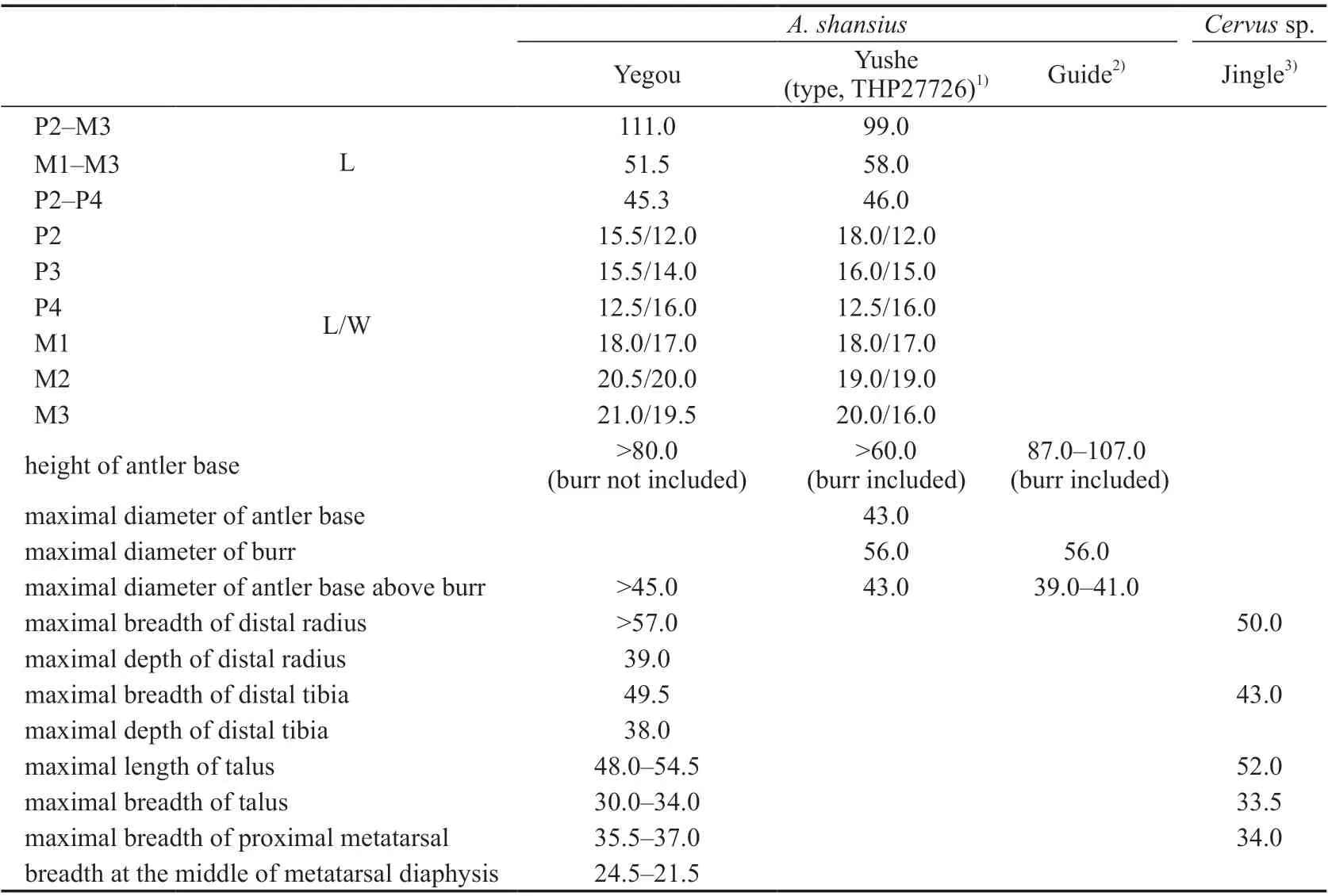
Table 15 Measuements of antlers, limb bones and cheek teeth of Axis shansius and Cervus sp. (mm)
Description The antler (V 18841.1, Fig. 9A) is fragmentary. Only the portion where the main beam and the brown tine bifurcates is preserved. The lateral side of the main beam is also damaged. The medial surface is concave, and longitudinal grooves and ridges can be seen. The lateral surface is flat, and the grooves and ridges are not as clear. Judging from the longitudinal orientation of the main beam, the angle between it and the brown tine is probably greater than 90°.
Enamel rugosity is distinct on the surface of the upper cheek teeth (V 18841.2, Fig. 9B).Spurs are developed on the buccal side of the crescent crests formed by the protocone and the metaconule. The cingulum is developed on the lingual and medial side. The lingual side of the P2 and P3 is double-lobed, and the protocone and the metaconule are developed. The P4 is single-lobed. The parastyle and the mesostyle are more developed than the metastyle on M1–3.The extent of buccal protruding of the parastyle is greater than that of the metastyle. The length of P2–4 is 88% of that of M1–3.
Only the distal portion of the radius (V 18841.4, Fig. 9C) is preserved, and the lateral side is damaged. The medial articular facet is convex, and the lateral articular is posteriorly positioned and concave.
Only the distal portion of the tibia (V 18841.5, Fig. 9D) is preserved. The articular portion is complete. The articulation with the talus is carried out by two facets. They are two anteroposteriorly oriented parallel concave grooves. The lateral one is wider than the medial one, but the latter one is deeper. The articulation with the lateral malleolus (vestigial distal end of the fibula) is carried out by two small facets on the lateral side. The anterior one is larger,and the posterior one is smaller. They are separated by a notch.
There are two tali. One is left (18841.6, Fig. 9E), and the other one is right (18841.7).The right one is smaller, but their morphologies are the same.
There are two metatarsals (V 18841.8–9, Fig. 9F) and both preserve only the proximal half. The groove on the dorsal side is narrow and extends distally to a very low position. On the medial side of the proximal end of the groove, there is a distinct vascular impression. The plantar groove is wider. The proximal plantar foramen is located at the proximal end of the plantar groove, and it proximodorsally penetrates the proximal metatarsal and opens at the center of the articular surface with the tarsals.
Comparisons and discussion In terms of the size of the limb bones (Table 15), the Yegou material is very close toCervussp. from Jingle (Teilhard de Chardin and Young,1931). In terms of the morphology and size of the antler, the Yegou specimen (V 18841.1) is comparable to Early PleistoceneAxis shansiusfrom Zone III of Yushe, Shanxi, but the antler is slightly longer. It is closer to the antler of Late PlioceneA. shansiusfrom Guide, Qinghai (Zheng et al., 1985). The dimensions of the upper cheek teeth are also generally consistent with the holotype ofA. shansius.

Material Fragmentary right maxilla with the M2–3 (IVPP V 18840, Fig. 10A).
Measurements M1: length, 10.8 mm, anterior width, 12.3 mm, posterior width, 11.3 mm; M2: length, 10.8 mm, anterior width, 12.0 mm, posterior width, 10.3 mm.
Description It is a small-sized muntjac. The molars are low-crowned. The morphologies of the M2 and M3 are similar. There is strong rugosity on the enamel surface. The protocone is more developed than the hypocone, and the paracone is more developed than the metacone.The parastyle and the mesostyle are more developed than the metastyle. The paracone rib is stronger than the metacone rib. Both teeth are 4-rooted. The only difference is that the M2 is slightly larger than the M3.
Comparisons and discussion The morphology and size of the teeth are consistent withMuntiacus. Due to the fact that there is no antler, it is hard to identify it to the species level.
Material Right M2 (IVPP V 18842.1, Fig. 10C), right M3 (V 18842.2, Fig. 10B), distal portion of the left humerus (V 18842.3, Fig. 10E), manual 4th proximal phalanx (V 18842.4,Fig. 10D).
Measurements See Tables 16–18.
Description The molars (IVPP V 18842.1–2, Fig. 10B, C) are large and anteriorly wider. The sloping of the lingual wall of the molars is gentle, so the flaring of the crown is not distinct. The protocone and metaconule are nearly equal in size, but the protocone extends more lingually. The parastyle is more developed than the mesostyle. The metastyle is extremely weak or not developed. The paracone rib is more robust than the metacone rib. The shrinking of the posterior lobe of the M3 is more distinct than that of the M2. The metastyle is relatively distinct on the M3.

Fig. 10 Specimens of Muntiacus sp. and Paracamelus sp. from Yegou A. Muntiacus sp. (1 cm scale bar): fragmentary right maxilla with M2–3 (IVPP V 18840: occlusal view);B–E. Paracamelus sp. (5 cm scale bar): B. right M3 (V 18842.2: 1. occlusal view, 2 buccal view);C. right M2 (V 18842.1: 1. occlusal view, 2. buccal view);D. manual 4th proximal phalanx (V 18842.4: 1. anterior view, 2. posterior view, 3. proximal view);E. distal portion of left humerus (V 18842.3: posterior view)
Only the distal portion of the humerus is preserved (V 18842.3, Fig. 10E). The lateral ligament pit is deep. The distal maximal breadth is located where the lateral epicondylar crest is in contact with the margin of the ligament pit. The medial ligament pit is shallow. There is a robust and short ridge posteroproximal to it. The trochlea of humerus is broader than the capitulum of humerus. The groove between them is shallow. The anteroproximal margin of the trochlea is more proximally positioned on the medial side. The posteroproximal margin of the trochlea extends proximally into the olecranon fossa. The posterodistal margin of the medial epicondyle meets the posterodistal margin of the diaphysis at an angle slightly less than 90°.
The distal articular surface of the proximal manual phalanx of 4th digit (V 18842.4, Fig.10D) is damaged, but the length of the bone is measurable. The proximal lateral ligament tuberosity is more prominent than the medial one, but the shape and size of the muscle insertion area on both sides are similar. The proximal articular surface is elliptical in shape.There is a depression at the center of the posterior portion of the surface that divides it into two parts. The lateral part is narrower and higher than the medial part. The distal end is slim and narrow. The ligament tuberosity on both sides is weak.
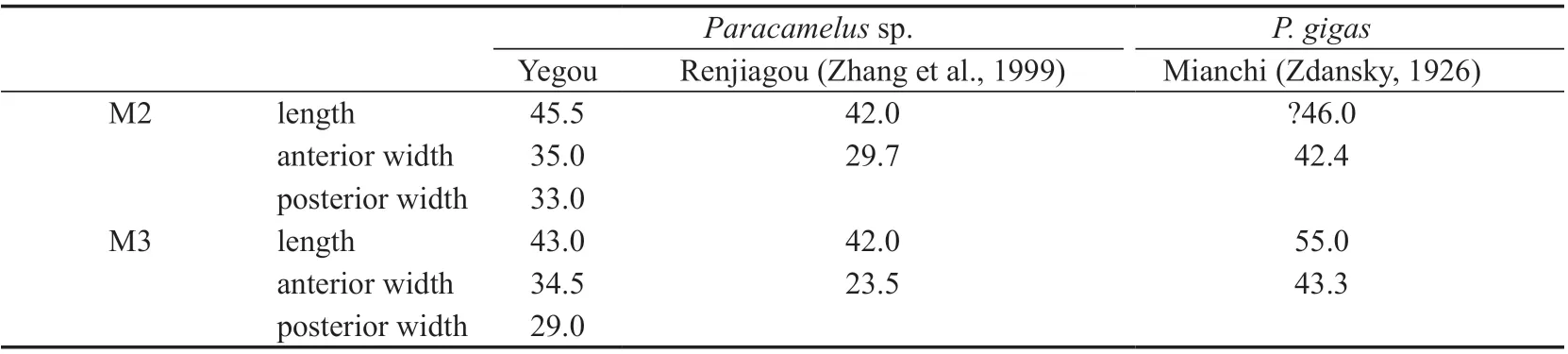
Table 16 Measurements of upper molars of Paracamelus sp. and P. gigas (mm)

Table 17 Measurements of the humerus of Paracamelus sp. from Yegou (mm)

Table 18 Measurements of the fourth proximal phalanx of selected camelins (mm)
Comparisons and discussionMaterial ofParacamelusgigasknown from the Quaternary strata in the Nihewan Basin is limited to one calcaneus and the distal portion of one radius (Teilhard de Chardin and Piviteau, 1930). That ofParacamelussp. from the Late Pliocene strata is also limited to one talus (Tang, 1980b). Yegou is therefore the first locality in the Nihewan Basin that has yielded dental remains.
There is material of maxilla with complete dentition, humerus, talus, and metatarsal known from the Upper Pliocene at Renjiagou, Lingtai, Gansu assigned toParacamelussp.(Zhang et al., 1999). The size (M2 and M3 length, 42.0 mm) and morphology (including shrinking posterior lobe, developed parastyle and mesostyle, weak metastyle) are the same as that of the Yegou specimen, but the humerus is neither described nor figured, so it can not be compared with the Yegou specimen.
The phalanx (V 18842.4) from Yegou is distinctly larger than that ofParacamelusgigasfrom Loc. 1, Zhoukoudian (Young, 1932) and Mianchi, Henan (Zdansky, 1926), and even larger than that of extant camels.
3 Composition and age of the Late Pliocene Hipparion houfenense fauna in Nihewan Basin
The localities or horizons that yield fossils of large mammals includingHipparionare relatively rare in the Nihewan Basin. In addition to Yegou and Laowogou (Cai et al., 2004),there are still the following localities: 1) the red clay layer of Luanshigedagou, Hongya Village, yieldingHipparionsp. andChilotheriumsp. (Huang et al., 1974: Layer 3); 2) the red clay layer of the “Yuxian Formation” of the upper portion of the Huabaogou section (Layer 3), Xiyaozitou Village, yieldingPostschizotheriumsp.,Viverrasp.,Hipparion houfenense,Gazella blacki,Gazellaspp., andAntispiroides hopeiensis(Wang, 1982; Li et al., 2008); 3) the red clay layer of the “Huliuhe Formation” of the lower portion of the Huabaogou section (Layer 1), Xiyaozitou Village, yieldingCanissp.,Canismulticuspus,Nyctereutessinensis,Hipparioncf.H. hippidiodus,Hipparionsp.,Palaeotragusspp.,Gazella blacki, andGazellasp. (Wang,1982; Li et al., 2008); and 4) the red clay layer of the Danangou section, Dongyaozitou Village,yieldingLynx variabilis,Nyctereutescf.sinensis,Hipparioncf.H. houfenense,Paracamelussp.,Antilospirayuxianensis, andPalaeotragus progressus(Layer 1 of Tang, 1980b and Tang and Ji, 1983; Layer 1–2 of Zheng and Cai, 1991 and Cai et al., 2004, 2013; Li et al., 2008).
If the horizon of the YegouHipparion houfenensefauna can be correlated to Layer 9 of the Laowogou section (Cai et al., 2004), Layer 1 or 3 of the Huabaogou section (Wang, 1982),and Layer 1 (Tang, 1980b; Tang and Ji, 1983) or Layer 1–2 (Zheng and Cai, 1991; Cai et al.,2004, 2013) of the east cliff section of Danangou, then the large mammals of the Late Pliocene mammalian fauna of the Nihewan Basin should tentatively include at least the following 27 forms:


Although most of these fossils have not been systematically studied (species with [-])or have only been briefly described (species with [+]) and need further taxonomic revision,their Pliocene age (compared with the classic Early Pleistocene Nihewan fauna) is affirmative.These taxonomic names are tentatively retained here for the following discussion in the sense of the fossil entities they represent, including the unidentified species and nomina nuda.Among them,Viverra,Nyctereutes,Dicerorhinus,Muntiacus,Gazella,Axisare extant genera.There are no extant species. Such a dominant proportion of extinct species is not possessed by any other Early Pleistocene faunas.
The key features of the Late PlioceneHipparion houfenensefauna of the Nihewan Basin represented by the 10 large mammals described here are as follows: 1) the coexistence ofGazella blackiandHippaion(Plesiohipparion)houfenense; 2) the coexistence ofNyctereutes tingiandN. sinensis; 3) the appearance ofPachycrocuta pyrenaica; and 4) the coexistence ofAxis shansiusandParacamelus.
According to the first feature, the age of the fauna is directly comparable to those of the red clay of Layer 1, 3 of the Huabaogou section near Xiyaozitou Village in Yuxian Basin (Wang,1982), the Jingle red clay of the Renjiagou section, Lingtai, Gansu (Zhang et al., 1999), the red clay of Xiaohongao, Hefeng, Jingle, Shanxi (Teilhard de Chardin and Young, 1931; Chen,1994), the Mazegou Formation of Yushe, Shanxi (Flynn et al., 1991; Deng and Hou, 2011).According to the second feature,Nyctereutes tingionly has Pliocene stratigraphic records in the Yushe Basin (Tedford and Qiu, 1991) and Leijiahe, Lingtai, Gansu (Huang et al., 1993)so far, while the coexistence ofN. tingiandN. sinensisonly occurs in the Yushe Basin and the Nihewan Basin. According to the third feature,Pachycrocuta pyrenaicais only known from strata before Villafranchian in France, Spain, and Ukraine (Depéret, 1890; Crusafont and Sondaar, 1971; Adrover et al., 1976; Howell and Petter, 1980), whereas in China, it is only known from the lower Mazegou Formation, the Gaozhuang Formation of Pliocene, and even the upper Mahui Formation of Upper Miocene in the Yushe Basin (Qiu, 1987; Deng et al.,2010; Deng and Hou, 2011). According to the fourth feature, in addition to the Late Pliocene records ofAxis shansiusin Guide, Qinghai (Zheng et al., 1985) and Laowogou, Daodi Village, the Nihewan Basin (Cai et al., 2004), the holotype of this species comes from Zone III of Yushe, Shanxi (belongs to the Lower Pleistocene);Paracamelusalso has both Pliocene records, such as Danangou, the Nihewan Basin (Tang, 1980b) and Renjiagou, Lingtai (Zhang et al., 1999), and Pleistocene records (Teilhard de Chardin and Piveateu, 1930; Teilhard de Chardin and Trassaert, 1937; Young, 1932). Taken as a whole, the age of the YegouHipparion houfenensefauna should be the Late Pliocene before 2.6 Ma.
The Mazegou Formation of the Yushe Basin, Shanxi is the horizon yielding the richest fossils of Late Pliocene large mammals, and 65 forms have been currently preliminarily identified (Flynn et al., 1991; Deng and Hou, 2011; Wang et al., 2017), which contains nearly all species of large mammals from Late Pliocene localities in North China. It is therefore a helpful standard to compare with when trying to determine if the age of a fauna is Late Pliocene. For example, the sand layer of Guide and the Leijiahe Formation of Lingtai can be correlated the Mazegou Formation to by sharingAnancus sinensis; the Leijiahe Formation can also be correlated to the Mazegou Formation by sharingHipparion pater.Accordingly, based on the combinations ofNyctereutes tingi–N. sinensisandHomotheriumsp.–Dicerorhinussp.–Paracamelussp., the Yegou horizon can also be correlated to the Mazegou Formation.
Nyctereutes tingi,Pachycrocuta pyrenaica,Homotheriumsp.,Hipparion(Plesiohipparion)houfenense, andGazella blackiof the Late Pliocene YegouHipparion houfenensefauna of the Nihewan Basin are more primitive thanN. sinensis,Pachycrocuta brevirostris licenti,Homotheriumcrenatidens,Hipparion(Proboscidipparion)sinense, andGazella sinensisof the Early Pleistocene fauna from the Xiashagou-Nihewan Village area,respectively. Consequently, the age of the former fauna should be older.
In the Nihewan Basin, the localities or horizons that yield common faunal elements with the YegouHipparion houfenensefauna include the following: 1) Layer 9 of Laowogou,sharingHipparionandAxis shansius(Cai et al., 2004); 2) Layer 1 of the east cliff section,Danangou, Dongyaozitou Village, sharingHipparioncf.H. houfenense(Tang, 1980b; Tang and Ji, 1983); 3) Layer 1 of Huabaogou, Xiyaozitou Village, sharingNyctereutessinensisandGazella blacki(Wang, 1982); and 4) Layer 3 of Huabaogou, Xiyaozitou Village, sharingHipparion houfenenseandGazella blacki(Wang, 1982). Based on biostratigraphic correlations(Li et al., 2008; Cai et al., 2013), Layer 3 of the Huabaogou section can be correlated to Layer 9 of Laowogou; Layer 1 of Huabaogou can be correlated to Layer 2 of Laowogou; Layer 1 of the east cliff section of Danangou can be correlated to Layer 16–19 of Laowogou. Accordingly,the horizon of the YegouHipparion houfenensefauna can be generally correlated to the above horizons, especially Layer 9 of Laowogou.
Presently, the magnetostratigraphic dating results of the horizons of theHipparion houfenensefauna characterized byHipparion(Plesiohipparion)houfenenseandGazella blackiare not completely consistent. The red clay of Xiaohongao, Jingle has been dated to 3 Ma (Chen, 1994). The red clay of Renjiagou, Lingtai has been dated to 3.5–3.4 Ma (Sun et al., 1998; Zhang et al., 1999). The Mazegou Formation has been dated to 3.6–2.6 Ma (Flynn and Qiu, 2013). The fossil layer of the east cliff section, Danangou, Dongyaozitou Village has also been dated to 3.04–2.58 Ma (Liu et al, 2021). It can be inferred from these results that the age of the Yegou horizon should fall within the range of 3.6–2.6 Ma. Only the red clay of Layer 1, 3 of the Huabaogou section is unexpectedly younger, the magnetostratigraphic age of them ranges 1.95–1.77 Ma (Zhu et al., 2007; Deng et al., 2008; Deng, 2011). The most likely reason for this is that depositional hiatus in the Huabaogou section may have been ignored by these investigators. This, once again, proves the importance of the mutual compatibility between biostratigraphy and magnetostratigraphy. For the magnetostratigraphic dating results to be robust, it is necessary to carefully verify if the correlation of polarity chrons to GPTS is compatible with biostratigraphy based on fossils, especially when contradictions arise. If contradictions arise, the results are subject to cautious reinterpretation, as is the case for the results mentioned above.
4 Features of Late Pliocene and Early Pleistocene mammalian faunas (large) in Nihewan Basin
In recent years, many Late Pliocene horizons yieldingHipparion houfenensehave been discovered in North China, such as Layer 1 (Tang, 1980b; Tang and Ji, 1983) or Layer 1–2(Zheng and Cai, 1991; Cai et al., 2004, 2013) of the Danangou section, Dongyaozitou Village and Layer 1, 3 of the Huabaogou section, Xiyaozitou Village in the Nihewan Basin (Wang,1982), Renjiagou, Lingtai (Zhang et al., 1999) and the Leijiahe Formation (Huang et al.,1993) in Lingtai, Gansu, the Mazegou Formation of Yushe (Flynn et al., 1991; Deng and Hou,2011) and Xiaohongao, Jingle (Zhou, 1988; Chen, 1994) in Shanxi, the sand layer of Guide in Qinghai (Zheng et al., 1985), and so on. The discovery of these Late Pliocene horizons with fossils of large mammals provides us with a better understanding about the general features of theHipparion houfenensefauna in China. It should be noted thatLynx variabilis,Hipparioncf.H. houfenense,Paracamelussp.,Antilospirayuxianensis, etc. in the “Pliocene–Early Pleistocene transitional mammalian fauna” derived from Layer 1 of the east cliff section of Danangou in the Nihewan Basin (Tang, 1980b; Tang and Ji, 1983) should be Late Pliocene in age (Zheng and Cai, 1991; Cai et al., 2004, 2013). Based on the fossils of large mammals derived from the above horizons, the Late Pliocene mammalian fauna (large) in the Nihewan Basin is characterized by having the following features: 1) few Early Pliocene forms, such asNyctereutes tingiTedford & Qiu, 1991,Pachycrocutapyrenacia(Depéret, 1890),Hipparioncf.H. hippidiodusSefve, 1927, andPalaeotragussp.; 2) several species unique to Late Pliocene,such asCanismulticuspusWang, 1982 (nomen nudum),Lynx variabilisTang, 1980,Hipparion(Plesiohipparion)houfenense(Teilhard de Chardin & Young, 1931),AntilospirayuxianensisTang, 1980,Antispiroides hopeiensisWang, 1982 (nomen nudum), andPalaeotragus progressusTang & Ji, 1983; 3) some Late Pliocene–Early Pleistocene transitional forms, such asHomotheriumsp. (Yegou),Nyctereutessinensis(Schlosser, 1903),Gazella blackiTeilhard et Chardin & Young, 1931, andParacamelussp. (Yegou, Danangou); and 4) no extant species.
In addition to the classic Early Pleistocene Nihewan mammalian fauna (large) with multiple revisions and supplements (Teilhard de Chardin and Piveteau, 1930; Qiu, 1987, 2000),numerous Early Pleistocene horizons yielding fossils of large mammals have been discovered in the Nihewan Basin in recent years, such as the Xiaochangliang site (You et al., 1980; Tang et al., 1981, 1995; Chen et al., 1999; Zhang et al., 2008), the Majuangou site (Wei et al., 2003;Cai and Li, 2004; Cai et al., 2008), the west section of Danangou (Tang et al., 1981; Li, 1984),Layer 3, 5 of the east cliff section of Danangou (Tang, 1980b; Tang and Ji, 1983), and so on.Based on the fossils of large mammals derived from these horizons, the Early Pleistocene mammalian fauna (large) in the Nihewan Basin is characterized by having the following features: 1) few Late Pliocene–Early Pleistocene transitional forms, such asZygolophodonsp. (Danangou),Nyctereutessinensis(Schlosser, 1903),Crocuta honanensis(Zdansky, 1924),Hipparionsp. (Xiaochangliang, Majuangou, Danangou),Elasmotheriumsp. (Nihewan Village),andPostschizotherium chardinivon Koenigswald, 1932; 2) a lot of species unique to the Early Pleistocene, includingMammuthus trogontheriiPohlig, 1885,CanischiliensisZdansky,1924,Canisc. var.palmidensTeilhard de Chardin & Piveteau, 1930,Eucyon minor(Teilhard de Chardin & Piveteau, 1930),Erictis pachygnatha(Teilhard de Chardin & Piveteau, 1930),Meles chiaiTeilhard de Chardin, 1940,Lutra licentiTeilhard de Chardin & Piveteau, 1930,Casmaporthetes progressus(Qiu, 1987),Pachycrocutalicenti(Pei, 1934),Homotherium crenatidensFabrini, 1890,Megantreon nihowanensis(Teilhard de Chardin & Piveteau, 1930),Sivapentaheracf.S. pleistocaenicus(Zdansky, 1925),Lynx shansiusTeilhard de Chardin,1945,Hipparion(Proboscidipparion)sinense(Sefve, 1927),Equus sanmeniensisTeilhard de Chardin & Piveteau, 1930,Coelodonta nihowanensisKahlke, 1969,Muntiacus bohlini(Teilhard de Chardin, 1940),Eucladoceros bouleiTeilhard de Chard & Piveteau, 1930,Nipponicervus elegans(Teilhard de Chardin & Piveteau, 1930),Elaphurus bifurcatesTeilhard de Chardin& Piveteau, 1930,Spirocerus wongiTeilhard de Chardin & Piveteau, 1930,GazellasinensisTeilhard de Chardin & Piveteau, 1930, andBison palaeosinensisTeilhard de Chardin & Piveteau,1930; and 3) a considerable number of Middle to Late Pleistocene and extant species, such asCanissp. (Majuangou),Vulpessp. (Danangou),Martessp. (Xiaochangliang),Ursus thibetanusG. Cuvier, 1823,Viverrasp. (Xiaochangliang),Felissp. (Nihewan Village),Suscf.S. lydekkeriZdansky, 1928,Axissp. (Danangou),Cervussp. (Nihewan Village, Xiaochangliang),Gazellasp.(Xiaochangliang),Gazellacf.G. subgutturosa(Güldenstädt, 1780),Ovissp. (Nihewan Village),Ovis shantungensisMatsumoto, 1926,Bisonsp. (Xiaochangliang), and so on.
AcknowledgementsThis work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (Grant No: XDB26000000), National Science Foundation of China (Grants No: 41772018, 41172150) and Basic Scientific Research Fund of the Institute of Geology, Chinese Academy of Geological Sciences (No: 1001). Prof. YAO Pei-Yi and Prof.YAO Zhen of Institute of Geology, Chinese Academy of Geological Sciences, Mr. CHEN Xing-Qiang of China University of Geosciences participated in the field investigation and fossil collection. Mr. WANG Zhao of Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences (IVPP) participated in the excavation and prepared the fossils.Mr. GAO Wei of IVPP photographed the specimens. Prof. QIU Zhang-Xiang of IVPP gave valuable comments and suggestions during the preparation of the manuscript. Prof. ZHANG Zhao-Qun and Prof. LI Qiang of IVPP provided helpful comments that improved the content of the article when reviewing the manuscript. We would like to express our profound gratitude to these institutions and individuals.
杂志排行

古脊椎动物学报(中英文)的其它文章
- Ulanodon, a new name for the Hyracodontid Ulania Qi,1990 (Perissodactyla, Mammalia)
- First discovery of dinosaur eggs in Nanhu Gebi of Hami,Xinjiang, China
- A new species of Pararhizomys (Tachyoryctoidinae,Muroidea) from Linxia Basin of Gansu Province
- Nothosaurus luopingensis sp. nov. (Sauropterygia)from the Anisian, Middle Triassic of Luoping,Yunnan Province, China