12个竹种的45S rDNA和5S rDNA分布特性比较*
2022-11-05徐川梅王子晗金旭胤吴心妍黄坚钦
徐川梅 王子晗 谢 峰 金旭胤 吴心妍 陈 荣 黄坚钦
(浙江农林大学 省部共建亚热带森林培育国家重点实验室 杭州 311300)
竹类植物属于禾本科(Gramineae)竹亚科(Bambusoideae)(张颖等, 2021),种类繁多,全世界约有1 500个种,可分为温带木本竹种、新热带木本竹种、古热带木本竹种和草本竹种4种类型,其中温带和新热带木本竹种染色体数目为2n=46~48,新热带木本竹种染色体数目为2n=70~72,草本竹种染色体数目为2n=22~24(Guoetal., 2019)。目前,竹类植物相关研究主要集中在栽培生理及分子生物学等领域,特别是相关的分子生物学研究取得了一定突破,毛竹(Phyllostachysedulis)、莪莉竹(Olyralatifolia)、Raddiaguianensis、瓜多竹(Guaduaangustifolia)和芸香竹Boniaamplexicaulis等5个竹种全基因组测序工作已完成(Pengetal., 2013; Zhaoetal., 2018; Guoetal., 2019);但竹类植物相关的细胞遗传学研究进展十分缓慢,仅有45S rDNA在毛竹、斑竹(Phyllostachysreticulata‘Lacrima-deae’)、龟甲竹(Phyllostachysedulis‘Heterocycla’)、茶秆竹(Pseudoasaamabilisvar.amabilis)、菲白竹(Sasafortunei)、日本矮竹(Sasapygmea)、铺地竹(Sasaargenteastriatus)和白绿竹(Bambusaoldhamii)等竹种染色体上的分布特点报道,大部分竹种的细胞遗传学研究工作还停留在染色体计数或传统的核型分析等初级细胞遗传学研究水平(李秀兰等, 1999; 2001; 陈瑞阳等, 2003; 徐川梅等, 2009a; 2009b; 贾芳信等, 2016; Zhouetal., 2017),这些研究仅能够揭示竹类植物染色体数目和形态大小等信息,并不能对一些竹种的染色体结构及核型等进行精细分析。
核糖体RNA(rDNA)基因广泛存在于植物基因组中,主要包括45S rDNA和5S rDNA 2种类型,其中45S rDNA主要由串联重复序列18S、5.8S和28S rRNA基因和非转录间隔区组成,重复单元长度约10 kb,主要负责形成核仁组织区(Osujietal., 1998; Katsiotisetal., 2000; Heslop-Harrison, 2000)。5S rDNA也是串联重复序列,主要包括一个长约120 bp的编码区和一个非转录间隔区(Zhangetal., 2016)。由于45S rDNA和5S rDNA在植物进化过程中高度保守,很多植物染色体上均可产生清晰的荧光信号,可为染色体相关研究提供一些重要参考信息,这使其成为一个非常好的染色体物理标记,在一些植物倍性鉴定、染色体结构分析、系统分类和起源进化等研究中发挥了重要作用(Luoetal., 2019; Zhangetal., 2016)。
植物基因组中的rDNA区域是遗传重组的热点区域,同源染色体间不等交换或重组,往往会导致rDNA拷贝数产生一定差异或者扩增出新的rDNA序列(Tsangetal., 2008)。此外,rDNA重复序列内部及邻近区域也是转座子高频插入区域,在物种形成和分化过程中,转座子插入或删除进一步促进了rDNA位点的进化,使物种核型不断变化,进而产生一定的遗传分化(Pedrosa-Harandetal., 2006; Raskinaetal., 2008)。因此了解不同物种的rDNA位点数及其在染色体上的分布特征,可为植物比较基因组学及分子系统学等研究提供重要参考信息。由于rDNA主要是一些串联重复序列,可利用荧光原位杂技术将这些rDNA在染色体上的分布位置进行可视化,且荧光信号强弱与rDNA拷贝数呈一定的正相关,因此,本研究选择毛竹、花毛竹(Phyllostachysedulis‘Tao kiang’)、龟甲竹、金镶玉竹(Phyllostachysaureosulcata‘Spectabilis’)、白哺鸡竹(Phyllostachysdulcis)、紫竹(Phyllostachysnigra)、斑竹、黄秆乌哺鸡竹(Phyllostachysvivaxf.aureocaulis)、早竹(Phyllostachysviolascens)、人面竹(Phyllostachysaurea)、大明竹(Pleioblastusgramineus)和茶秆竹(Pseudoasaamabilis)等12个竹种,首次利用双色荧光原位杂交技术分析45S rDNA和5S rDNA在这些竹种染色体上的分布,以期为竹类植物系统分类及染色体结构等研究提供相应的分子细胞遗传学资料,为相关的染色体识别提供相应的染色体物理标记。
1 材料与方法
1.1 材料
12个竹种均采自浙江农林大学翠竹园,3—6月,挖取新竹或新笋,采集新竹或新笋的嫩根为试验材料。
1.2 方法
1.2.1 材料处理 采集根尖利用0.002 mol·L-18-羟基喹啉与秋水仙素混合液室温下处理4 h,之后利用固定液(甲醇∶乙酸= 3∶1,V/V)固定,固定后的根尖置于-20 ℃冰箱保存备用。
1.2.2 染色体标本制备及制片预处理 染色体标本制备参照陈瑞阳等(2003)方法略进行改进,利用2.5%纤维素酶和果胶酶混合液37 ℃酶解根尖2~2.5 h,制备成细胞悬浮液,后进行染色体滴片。利用相差显微(Olympus BX51)对染色体制片进行观察和筛选,选取分散较开、形态较好的中期染色体制片。利用10 mg·mL-1的RNase A 37 ℃处理制片60 min,然后用4%(质量百分比)的胃蛋白酶溶液37 ℃处理10 min,再用4%(质量百分比)多聚甲醛室温固定10 min,最后用70%、90%及100%酒精对制片进行梯度脱水,每级5 min,处理后的制片用于后续荧光原位杂交试验。
1.2.3 荧光原位杂交与信号检测 荧光原位杂交参照Jiang等(1995) 方法,45S rDNA和5S rDNA序列分别来源于小麦(Triticumaestivum)的pTa71和日本莲花(Lotusjaponicas)(Pedrosaetal., 2002)。分别利用生物素(Roche,11745824910)和地高辛(Roche,11745816910)采用切刻平移法对45S rDNA和5S rDNA的质粒DNA进行标记,杂交信号经抗地高辛抗体Anti-digoxigenin-rhodamine(Roche,11207750910)和抗生物素抗体Avidin-fluorescein(Roche,11975595910)检测,用DAPI(4′,6-diamidine-2′-phenylindole dihydrochloride)对染色体进行套染,利用荧光显微镜(ZEISS,AX10)100倍油镜对荧光原位杂交结果进行观察和拍照,每个材料选取20个良好的分裂相进行观察和分析。
2 结果与分析
2.1 45S rDNA和5S rDNA在毛竹、花毛竹、龟甲竹及早竹染色体上的分布
毛竹、花毛竹、龟甲竹及早竹在系统分类上均属于刚竹属,染色体数目均为2n=48。毛竹具有1对45S rDNA信号,分布于1对同源染色体末端,5S rDNA在毛竹染色体上有2对强弱不同的荧光信号,其中较强的1对信号位于1对同源染色体末端,另1对信号非常微弱,位于另1对同源染色体的近中间区域,如白色箭头所示(图1A,表2),双色FISH检测结果显示45S rDNA和5S rDNA位于毛竹的不同染色体上,在分布位置上没有任何重叠。花毛竹也具有1对45S rDNA位点,位于1对同源染色体末端,且这对45S rDNA位点信号较强(图1B,表1),5S rDNA在花毛竹染色体上也存在2对强弱不同的荧光信号,其中较强的1对信号位于1对同源染色体末端,而另1对信号非常微弱,几乎观察不到,位于另1对同源染色体的近中间区域,如白色箭头所示(图1B,表1),45S rDNA和5S rDNA在花毛竹染色体上也没有任何重叠分布(图1B )。45S rDNA和5S rDNA在龟甲竹上的分布模式与花毛竹相似,1对较强的45S rDNA信号分布于1对同源染色体的近端部(图1C,表1),其5S rDNA信号也是2对,其中较强的1对信号位于1对同源染色体末端,另外1对微弱的荧光信号位于另1对同源染色体近中间区域,如白色箭头所示(图1C,表2),45S rDNA和5S rDNA在龟甲竹染色体上也没有任何重叠分布(图1C)。早竹45S rDNA的分布模式与毛竹相似,具有1对末端45S rDNA信号,与毛竹、花毛竹及龟甲竹的5S rDNA分布模式不同,早竹具有2对较强的5S rDNA信号,且2对信号均位于2对不同的同源染色体的近中间区域(图1D,表1),45S rDNA和5S rDNA在早竹染色体上也没有重叠分布(图1D)。

表1 12个竹种的45S rDNA和5S rDNA分布特征Tab.1 The distribution of 45S rDNA and 5S rDNA of 12 bamboo species
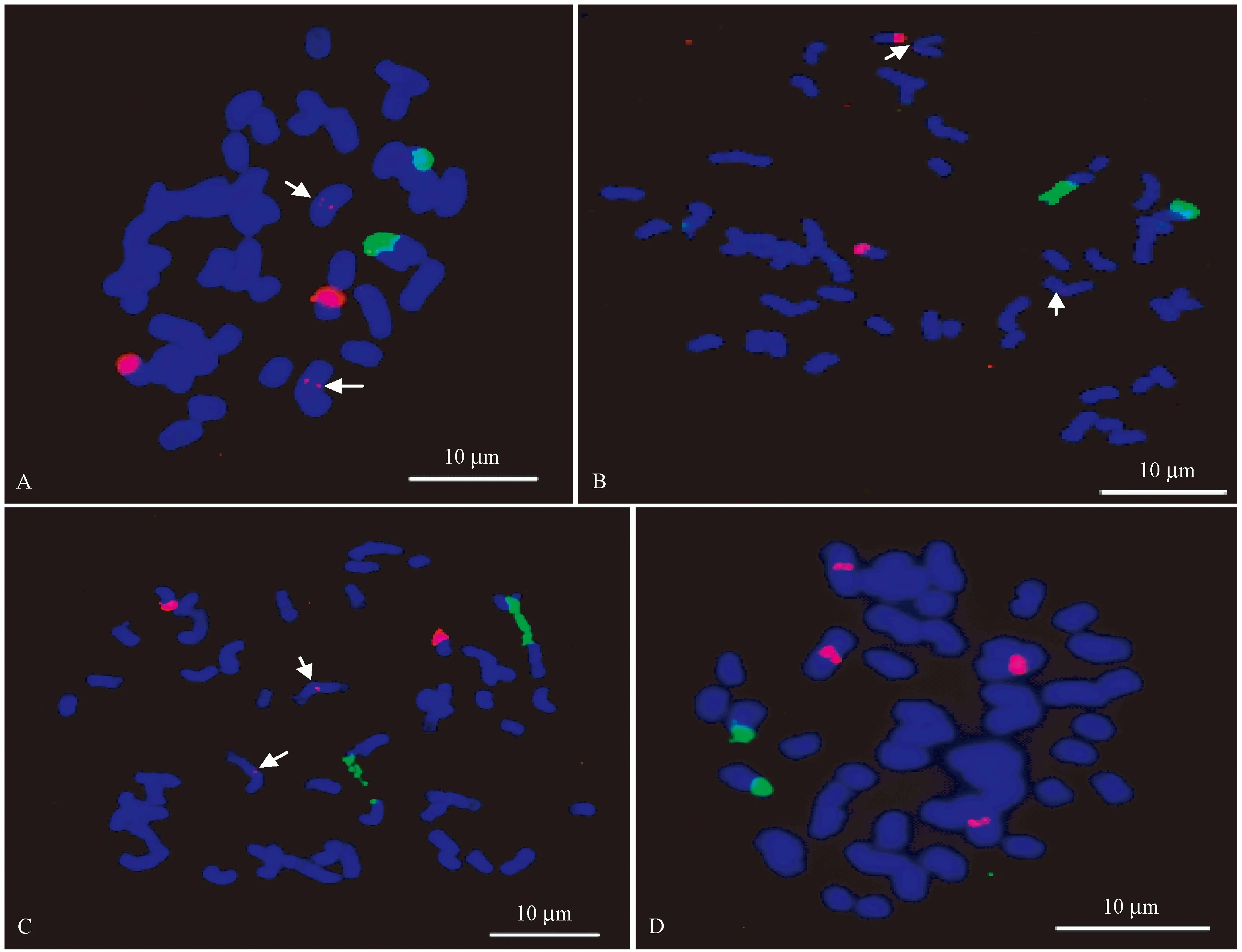
图1 45S rDNA(绿色)和5S rDNA(红色)在4个不同竹种染色体上的分布结果Fig.1 The distribution of 45S rDNA (green) and 5S rDNA (red) on the chromosome of four different bamboo speciesA-D分别为毛竹、花毛竹、龟甲竹及早竹,白色箭头所示为5S rDNA弱信号。A to D is Phyllostachys edulis, Phyllostachys edulis ‘Tao kiang’, Phyllostachys edulis ‘Heterocycla’ and Phyllostachys violascens, respectively, white arrows show the weak signals of 5S rDNA.
2.2 45S rDNA和5S rDNA在白哺鸡竹、黄秆乌哺鸡竹及斑竹染色体上的分布
白哺鸡竹、黄秆乌哺鸡竹及斑竹在系统分类上均属于刚竹属,染色体数目均为2n=48。图2A1-A3和表1为45S rDNA和5S rDNA在白哺鸡竹染色体上的分布结果,可以看出45S rDNA在白哺鸡竹染色体上有1对信号,分布于1对同源染色体端部,且其中一条染色体上的45S rDNA位点发生断裂,如图2A1所示。5S rDNA在白哺鸡竹染色体上有2对清晰明显的信号,且2对信号强度差异不大,位于2对同源染色体的近中间区域(图2A2、A3,表1),除此之外,在45S rDNA位点处也观察到了微弱的5S rDNA信号,如黄色箭头所示(图2A2),对这种特殊现象进行统计,70个分裂相中有32个分裂相可观察到这种现象,约占45.49%。图2B1和表1为45S rDNA和5S rDNA在黄秆乌哺鸡竹染色体上的分布结果,可以看出黄秆乌哺鸡竹染色体具有1对45S rDNA信号,位于1对同源染色体端部,这对信号位点也存在明显的断裂现象(图2B),黄秆乌哺鸡竹染色体上也具有2对清晰明显的5S rDNA信号,位于2对同源染色体的近中间区域(图2B1,表1),45S rDNA和5S rDNA在黄秆乌哺鸡竹染色体上也没有任何重叠(图2B)。斑竹染色体上具有1对45S rDNA信号,这对45S rDNA信号荧光强度一致,位于1对同源染色体末端,且存在一定的断裂现象(图2C,表1); 斑竹染色体上有2对5S rDNA信号,这2对5S rDNA荧光信号较弱,位于2对同源染色体的近中间区域,如白色箭头所示(图2C,表1),45S rDNA和5S rDNA在斑竹染色体上也没有任何重叠分布(图2C)。

图2 45S rDNA(绿色)和5S rDNA(红色)在3个不同竹种染色体上的分布结果Fig.2 The distribution of 45S rDNA (green) and 5S rDNA (red) on the chromosome of three different bamboo speciesA1-A3为白哺鸡竹,B为黄秆乌哺鸡竹,C为斑竹。A1 to A3 is Phyllostachys dulcis, B to C is Phyllostachys vivax f. aureocaulis and Phyllostachys reticulata ‘Lacrima-deae’, respectively.
2.3 45S rDNA和5S rDNA在人面竹、金镶玉竹及紫竹染色体上的分布
人面竹、金镶玉竹及紫竹在系统分类上均属于刚竹属,染色体数目均为2n=48。由图3A和表1可以看出人面竹具有2个45S rDNA信号位点,位于染色体端部,且其中1个45S rDNA位点有明显的断裂现象,如图3A1虚线所示。人面竹具有2对5S rDNA信号位点,其中1对位点信号强度相似,但另外1对信号表现为1强1弱,如图3A1白色和黄色箭头所示,这2对信号均位于两对同源染色体的近中间区域,且45S rDNA和5S rDNA分布于人面竹不同的染色体上(图3A)。图3B和表1为45S rDNA和5S rDNA在金镶玉竹染色体上的分布情况,可以看出金镶玉竹也具有1对45S rDNA信号位点,均位于染色体端部,其中1个45S rDNA位点存在明显的断裂现象,如图3B虚线所示,而金镶玉竹的5S rDNA分布模式比较特殊,有3个信号位点存在,其中2个位点位于2条染色体的近中间位置,且这2个位点在信号强弱上存在一定差异,如图3B白色和红色箭头所示,另外1个位点信号较强,位于1条染色体的末端,如图3B黄色箭头所示。金镶玉竹的45S rDNA和5S rDNA也是位于不同的染色体上,在分布位置上没有任何重叠(图3B)。45S rDNA在紫竹染色体上的分布情况比较特殊,虽然45S rDNA位点也是1对,但这2个位点存在较大差异,其中1个位点信号较强,但极易断裂,如图3C虚线所示,而另外1个位点信号非常微弱,甚至检测不到,位于染色体末端,如图3A3红色箭头所示。紫竹的5S rDNA分布模式也比较特殊,与金镶玉竹情况相似,也具有3个位点,其中2个位点信号非常强,以成对的形式位于1对同源染色体末端,但另外1个位点信号微弱,甚至检测不到,单独位于1条染色体的近中间区域,如黄色和白色箭头所示(图3C,表1),45S rDNA和5S rDNA也是位于紫竹的不同染色体上(图3C)。
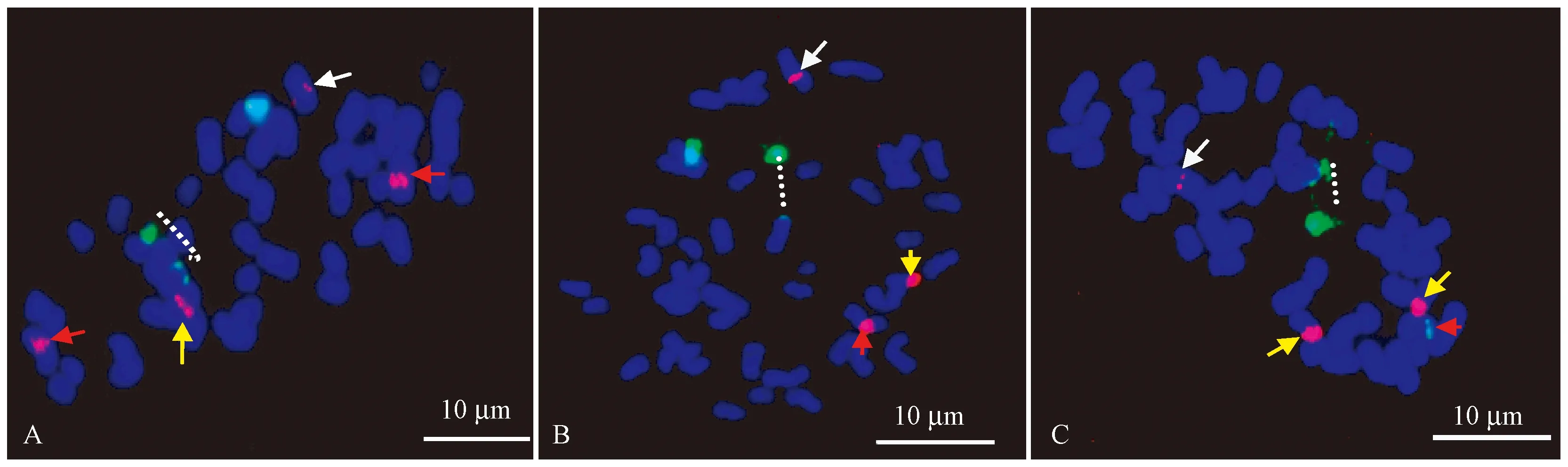
图3 45S rDNA(绿色)和5S rDNA(红色)在3个不同竹种染色体上的分布结果Fig.3 The distribution of 45S rDNA (green) and 5S rDNA (red) on the chromosome of three different bamboo species.A-C分别为人面竹、金镶玉竹和紫竹。A to C is Phyllostachys aurea, Phyllostachys aureosulcata ‘Spectabilis’,and Phyllostachys nigra, respectively.
2.4 45S rDNA和5S rDNA在茶秆竹及大明竹染色体上的分布
茶秆竹和大明竹在系统分类上分别属于茶秆竹属和大明竹属,染色体数目均为2n=48。图4A1和表1分别为45S rDNA (绿色)和5S rDNA(红色)在茶秆竹染色体上的分布,45S rDNA在茶秆竹染色体上有2个位点,分布于染色体短臂的次缢痕部位(图4A1),试验过程中观察到茶杆竹其中一条染色体上的45S rDNA位点极易断裂成2段,断开的2个45S rDNA片段均可产生很强的荧光信号,导致很多细胞中出现3个45S rDNA位点信号(图4B,红色),对这类细胞进行统计,约占55.20%(共统计154个分裂相)。5S rDNA(红色)在茶秆竹染色体上分布模式比较固定,只有1对分布位点,且信号较弱,位于染色体近中间区域(图4A1),45S rDNA和5S rDNA均位于茶秆竹不同的染色体上(图4A1)。45S rDNA(绿色)在大明竹染色体上有2个信号位点,分布于大明竹染色体末端,但这2个位点在信号大小上存在一定差异(图4 B,表2),5S rDNA(红色)在大明竹染色体上分布情况也比较特殊,以奇数形式存在,其中2个位点以成对的形式分布于染色体短臂中间,另1个位点单独分布于染色体近着丝粒区域(图4B,表2),大明竹的45S rDNA和5S rDNA也是分布于不同的染色体上(图4B)。
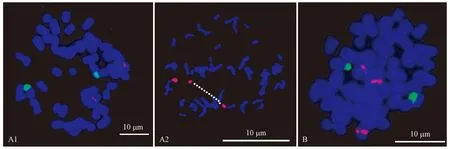
图4 45S rDNA和5S rDNA在2个不同竹种染色体上的分布Fig.4 The distribution of 45S rDNA (green) and 5S rDNA (red) on the chromosome of two different bamboo speciesA1和B分别为45S rDNA(绿色)和5S rDNA(红色)在茶秆竹和大明竹染色体上的分布,A2为45S rDNA(红色)在茶杆竹染色体上的分布。A1 and B is the distribution of 45S rDNA (green) and 5S rDNA(red)on the chromosome of Pseudoasa amabilis and Pleioblastus gramineus, respectively. A2 is the distribution of 45S rDNA (red) on the Pseudoasa amabilis.
3 讨论
本研究首次系统分析45S rDNA和5S rDNA在毛竹、花毛竹、金镶玉竹、白哺鸡竹、龟甲竹、紫竹、斑竹、黄秆乌哺鸡竹、早竹、人面竹、茶秆竹及大明竹等12个竹种染色体上的物理分布,结果表明,这些竹种的45S rDNA位点数比较一致,5S rDNA分布模式彼此间存在一定差异。Garcia等(2017)曾对1 791个物种的45S rDNA分布特性进行分析,结果发现,大部分物种的45S rDNA主要位于染色体端部,特别是只有1对45S rDNA位点存在的物种,45S rDNA更易分布于染色体末端。本研究中的12个竹种,近11个竹种的45S rDNA位点位于染色体末端,说明竹类植物45S rDNA分布模式与大多数植物的45S rDNA分布趋势一致。毛竹测序结果表明,其45S rDNA数为2个(Pengetal., 2013),另外徐川梅等(2009a)曾对毛竹、斑竹及茶秆竹等竹种的45S rDNA分布特性进行研究,结果表明,这些竹种的45S rDNA位点数均为2个,本研究与以上研究结果基本一致。另外,虽然大部分竹种具有2个末端45S rDNA位点,但一些竹种的2个45S rDNA位点在形态上却表现出了一定差异,特别是紫竹、大明竹、白哺鸡竹及金镶玉竹等,以紫竹和大明竹为例,其2个45S rDNA位点在信号强弱上存在一定差异,紫竹表现更为突出,其1条染色体上的45S rDNA信号非常强,而另1条染色体上的45S rDNA信号非常弱,几乎检测不到。一些研究证实,45S rDNA是一个脆性位点,该区域的DNA重复序列极易断裂,是一个重组热点区域(Monkheangetal., 2016; Huangetal., 2008)。研究过程中发现,黄杆乌哺鸡竹、白哺鸡竹、斑竹、金镶玉竹、紫竹及茶秆竹等竹种的45S rDNA位点区域有明显的断裂痕迹,对这些竹种的45S rDNA断裂率进行了初步统计,结果表明,这些竹种的45S rDNA断裂率介于55.2%~95.59%之间,特别是白哺鸡竹和紫竹,其45S rDNA断裂率高达90%以上,这进一步说明这些竹种的45S rDNA区域也是一个脆性区域,紫竹的45S rDNA异质性可能与45S rDNA位点断裂及易位重组等存在一定关系。此外,这些竹种的45S rDNA在断裂方式方面也存在一定差异,例如,紫竹和金镶玉竹等竹种,其45S rDNA断点靠近染色体末端部位,大部分45S rDNA片段脱离染色体,染色体末端仅留少量45S rDNA片段,而茶秆竹的45S rDNA易从中间或近中间位置断开,断开后的45S rDNA片段变成2个荧光信号较强的位点,因此存在45S rDNA断裂的细胞中可产生3个明亮的荧光信号,且这类细胞所占比例较高(约55.20%),这在一定程度上对试验结果判读造成了一定干扰。
与45S rDNA相比,5S rDNA在竹类植物上展现出了丰富的多态性分布,根据5S rDNA在12个竹种上的分布特征,可归纳为4类。第一种分布类型的竹种具有2对5S rDNA信号位点,其中1对5S rDNA信号非常强,位于染色体端部,另外1对5S rDNA信号极弱,甚至检测不到,位于染色体近中部区域,毛竹、花毛竹及龟甲竹3个竹种的5S rDNA分布模式均属于这一类型。第二种分布类型的竹种也具有2对5S rDNA信号位点,但2对5S rDNA信号强度差异不明显,且均分布于染色体近中部区域,早竹、白哺鸡竹、黄秆乌哺鸡竹、斑竹及人面竹均属于这种分布类型。第三种分布类型的竹种具有1对5S rDNA信号位点,分布于染色体近中部区域,仅茶秆竹属于这种分布类型。第四种分布类型比较特殊,这些竹种的5S rDNA存在异质性分布,具有3个明显不对称的5S rDNA位点,金镶玉竹、紫竹及大明竹均属于这种分布类型。理论上同源染色体在序列组成上是相同的,因此对应的rDNA分布模式也应完全相同,但一些竹种的45S rDNA或5S rDNA表现出了明显的异质性,一些研究证实,其他一些植物中也存在rDNA异质分布现象,例如Philodendronangustilobum、Scadoxusmultiflorus及Paspalumquadrifarium等植物,45S rDNA位点均为奇数,表现出了明显的异质性(Vasconcelosetal., 2018; Monkheangetal., 2016; Vaioetal., 2005),Philodendronbipennifolium具有3个5S rDNA位点,2个以成对形式位于1对同源染色体的长臂近端处,另外1个位点单独位于1条染色体的短臂处,Philodendronglaziovii2个5S rDNA位点在形态大小及信号强度上存在极大差异(Vasconcelosetal., 2018)。一些研究证实,即使亲缘关系很近的一些物种间,其染色体上的rDNA位点数及分布位置有时也会存在一定差异,这些差异可能由以下3个机制所导致: 一是染色体间发生了不等交换或易位,二是rDNA区域染色体结构发生了快速重排,三是多倍化后导致DNA序列丢失(Badaevaetal., 2007; Mondinetal., 2011)。因此,根据45S rDNA和5S rDNA在上述竹种中的分布结果,推测这些竹种在染色体结构上可能也发生过诸如插入、缺失、易位及不等交换等类型的染色体重组现象,导致不同竹种在45S rDNA和5S rDNA分布模式上存在较大差异,甚至一些竹种的rDNA呈现出一定的异质性分布。
一些研究者根据45S rDNA和5S rDNA在一些植物染色体上的分布特征,将45S rDNA和5S rDNA的位置关系分为独立排列(S-type arrangement,S-type)和连锁排列(Linked-type arrangement,L-type)两种类型,其中S-type是指45S rDNA和5S rDNA独立分布于不同染色体上,约86.57%的植物属于这种分布类型,L-type是指45S rDNA和5S rDNA分布于同一条染色体上,这种分布类型的植物较少,约占4.21%(Garciaetal., 2017)。在L-type分布类型中,45S rDNA和5S rDNA分布位置表现形式多样,有些植物的45S rDNA和5S rDNA位于同一条染色体臂上,且位置十分接近,例如Cucumiszambianus、Cucumismetuliferus及Brassicaoxyrrhina等(Zhangetal., 2016; Hasteroketal., 2006),还有些植物的45S rDNA和5S rDNA在位置上几乎重叠,例如Brassicafruticulosa,还有些植物的45S rDNA和5S rDNA虽然位于同一条染色体上,但是位于不同染色体臂上,例如Cucumisdipsaceus(Zhangetal., 2016)。本研究中大部分竹种的45S rDNA和5S rDNA分布于不同的染色体上,因此这些竹种的45S rDNA和5S rDNA分布类型属于S-type。但是,在白哺鸡竹的45S rDNA位点处,检测到了微弱的5S rDNA信号,且对应位置的5S rDNA信号在形状大小与45S rDNA完全相同,位置也完全重叠,只是信号较弱,因此推测白哺鸡竹部分5S rDNA位点与45S rDNA在位置关系上存在连锁关系。除白哺鸡竹外,在毛竹的45S rDNA位点处观察到了非常微弱的5S rDNA信号,这类细胞比例较低,约为21.43%,且其表现没有白哺鸡竹明显。
另外,本研究所分析的12个竹种除大明竹和茶秆竹分别属于大明竹属和茶秆竹属外,其余10个竹种均属于刚竹属,在rDNA分布模式上,茶杆竹与大明竹除了彼此间存在一定差异外,与刚竹属的其他一些竹种间也存在着较大差异。刚竹属是包含竹种较多的一个属,根据形态学特征,刚竹属竹种被分为刚竹组(PhyllostachysSect. Phyllostachys)和水竹组(PhyllostachysSect. Heteroclade)2个不同的组(马乃训等, 2014; Zhangetal., 2019),本研究所涉及的10个刚竹属竹种,除紫竹属于水竹组外,其余9个竹种均属于刚竹组,而紫竹的45S rDNA和5S rDNA分布表现出了明显的异质性,与刚竹组的其他9个竹种存在较大差异。另外,刚竹组的9个竹种中,毛竹、花毛竹及龟甲竹3个竹种的亲缘关系最近,形态特征也非常相似,而这3个竹种的45S rDNA和5S rDNA分布模式也十分相似。因此,本研究中一些竹种的细胞学研究结果在一定程度上与这些竹种的系统分类结果具有一定的一致性。
4 结论
本研究利用双色荧光原位杂交技术分析了45S rDNA和5S rDNA在毛竹、花毛竹、龟甲竹、早竹、金镶玉竹、白哺鸡竹、紫竹、斑竹、黄秆乌哺鸡竹、人面竹、茶秆竹及大明竹12个不同竹种染色体上的分布,结果表明,紫竹的45S rDNA位点存在明显的异质性,而金镶玉竹、紫竹及大明竹3个竹种的5S rDNA位点为奇数,也存在明显的异质性。此外,45S rDNA和5S rDNA除了在以上竹种中存在异质性分布外,45S rDNA在绿秆人面竹、斑竹、黄杆乌哺鸡竹、白哺鸡竹及金镶玉竹等竹种上存在明显的断裂现象。根据以上研究结果,可以推断,在进化过程中,这些竹种的染色体可能发生过断裂、易位、插入或缺失等结构变异。另外,龟甲竹和花毛竹在形态上与毛竹非常相似,这2个竹种在系统分类上属于毛竹的变种,而这3个竹种的45S rDNA和5S rDNA分布模式也非常相似,这在一定程度上与3个竹种的系统分类地位具有一定的一致性。上述45S rDNA和5S rDNA的分布特性可作为相应的染色体物理标记,用以对相关染色体的识别和鉴定。
