酸枣仁总皂苷改善对氯苯丙氨酸诱导失眠大鼠模型学习记忆损伤机制的研究
2022-11-05衡依然段慧竹杜晨晖
衡依然,段慧竹,杜 鹤,闫 艳,杜晨晖*
酸枣仁总皂苷改善对氯苯丙氨酸诱导失眠大鼠模型学习记忆损伤机制的研究
衡依然1,段慧竹1,杜 鹤1,闫 艳2*,杜晨晖1*
1. 山西中医药大学 中药与食品工程学院,山西 太原 030619 2. 山西大学 中医药现代研究中心,山西 太原 030006
基于代谢组学技术考察酸枣仁总皂苷(total saponins of,SZSS)对失眠大鼠内源性代谢物的影响,并探讨酸枣仁总皂苷改善学习记忆的药效及其作用机制。采用对氯苯丙氨酸(-4-chlorophenylalanine,PCPA)诱导大鼠失眠模型,给予地西泮和酸枣仁总皂苷干预后,进行旷场实验及Morris水迷宫实验;采用苏木素-伊红(HE)染色观察海马组织病理变化;采用ELISA测定海马组织5-羟色氨(5-hydroxytryptamine,5-HT)、γ-氨基丁酸(γ-aminobutyric acid,GABA)及NO(nitric oxide,NO)水平。利用UPLC-Orbitrap-MS/MS技术对大鼠海马组织代谢轮廓进行分析,结合多元统计分析筛选差异性代谢物,并基于MetaboAnalyst 5.0数据库进行代谢通路及代谢网络分析,多角度探讨酸枣仁总皂苷改善学习记忆的代谢特征及作用途径。与模型组比较,酸枣仁总皂苷组大鼠学习能力显著提高(<0.05、0.01),海马组织病理结构明显改善,海马组织中5-HT、GABA及NO水平显著升高(<0.05、0.01);代谢组学共鉴定出18种与学习记忆损伤相关的潜在生物标志物,酸枣仁总皂苷主要通过苯丙氨酸、酪氨酸和色氨酸等7条主要氨基酸代谢通路发挥改善记忆损伤的作用。酸枣仁总皂苷能够通过回调海马组织内源性差异代谢物,并调控相关氨基酸代谢通路,从而改善失眠引起学习记忆减退。
酸枣仁总皂苷;失眠;学习记忆;海马;差异代谢物
酸枣仁为鼠李科植物酸枣Millvar.(Bunge) Hu ex H. F. Chou的干燥成熟种子,具有补肝、宁心、敛汗、生津的功效[1]。酸枣仁中主要含有皂苷、黄酮、脂肪油和挥发油等多种成分。其中,酸枣仁皂苷是酸枣仁中含量较高、化学结构复杂及药理作用广泛的一类有效成分[2-3]。目前已从酸枣仁中鉴定到31种皂苷类成分[4],主要有达玛烷型四环三萜类皂苷和齐墩果烷型、羽扇豆烷型、ceanothane型等五环三萜类皂苷。现代药理学研究表明,酸枣仁皂苷是酸枣仁发挥镇静安神作用的主要药效成分,在神经系统、免疫系统、心脑血管系统等方面亦表现出广泛的药理活性[5-11]。
失眠在睡眠障碍的分类中占主导地位,并在近年来成为一个日益严重的公共问题,根据我国睡眠研究会公布的最新调查结果显示,2022年中国成年人的失眠发生率高达38.2%[12]。失眠会影响学习记忆的形成和巩固,并引发焦虑、抑郁等精神疾病,严重影响工作和生活[13-15]。研究表明,失眠通过影响海马结构及神经递质的水平进而影响海马神经可塑性[16],造成学习记忆损伤;失眠通过引起氧化应激反应抑制海马中抗氧化酶的生成、活性氧的过度增加、脂质过氧化和DNA修饰等生理或病理过程而引起神经元损伤,间接造成学习记忆损伤[17];失眠可通过激活海马中炎症因子,导致神经损伤,进而造成学习记忆损伤[18]。海马对睡眠觉醒也发挥着重要的调节作用[19]。达玛烷型皂苷如人参皂苷[20-21]、知母皂苷[22]、三七皂苷[23]等均能够改善实验动物的学习记忆能力,酸枣仁皂苷类成分能够改善痴呆[24]、阿尔茨海默病(Alzheimer’s disease,AD)[25]等动物模型的学习记忆能力,但对于失眠引起的学习记忆能力减退研究未见报道。酸枣仁皂苷能否改善失眠导致的学习记忆损伤,并且如何发挥作用值得进一步研究。
对氯苯丙氨酸(-4-chlorophenylalanine,PCPA)失眠模型是评价药物镇静催眠作用中最常用的诱导睡眠障碍药理模型[26-27]。PCPA是一种特异的、不可逆的色氨酸羟化酶(tryptophan hydroxylase,TPH)抑制剂,TPH是5-羟色胺(5-hydroxytryptamine,5-HT)生物合成的限速酶。色氨酸可经TPH催化生成5-羟色氨酸(5-hydroxytryptamine,5-HTA),5-HTA进一步脱羧生成5-HT。因此,ip PCPA会抑制色氨酸羟基化而阻断5-HT的合成[28],从而导致动物外周和中枢神经系统5-HT水平显著降低,昼夜节律消失,几乎完全失眠[29]。5-HT是与睡眠关系最密切的神经递质之一,在慢波睡眠诱导和维持中具有重要作用[30]。目前以5-HT水平的显著变化作为PCPA失眠模型评价指标在文献中较为常见[31-34]。海马区是大脑中枢神经系统中最复杂的部位,主要参与机体近期记忆中的情节记忆及空间位置的学习[35]。外源性注射PCPA后,海马结构受到损伤,引起单胺类神经递质(5-HT、去甲肾上腺素、多巴胺等)、氨基酸类神经递质[γ-氨基丁酸(γ-aminobutyric acid,GABA)、谷氨酸等]及与记忆相关的细胞因子[一氧化氮(nitric oxide,NO)、白细胞介素-6(interleukin-6,IL-6)、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α等]水平变化[36],进而导致学习记忆损伤[37]。
近年来,代谢组学作为一种新型技术手段,能够更加可靠、灵敏地反映疾病或药物作用引起的机体生物功能变化,筛选调节机体的潜在生物标志物。孟喜胜等[38]对恒清II号方干预的APPswe/PS1dE9双转基因小鼠进行了代谢组学研究,发现其抗AD作用及改善认知障碍与氨基酸生物合成和代谢通路相关。邵晓妮等[39]基于1H-NMR的代谢组学技术对高尿酸血症认知功能障碍大鼠的皮层和海马进行检测,分析其诱发认知功能障碍机制可能与能量代谢的糖酵解和三羧酸循环、氨基酸代谢等有关。目前代谢组学已被广泛用于神经系统类疾病,包括疾病的诊断和机制研究、寻找药物作用靶点和生物标志物等研究[40]。
为深入探讨酸枣仁总皂苷改善失眠大鼠学习记忆的作用及其机制,本研究复制PCPA失眠大鼠模型,采用旷场实验总路程、静止时间及血清中5-HT水平评价失眠模型;通过逃逸潜伏期、目标象限停留时间、目标象限总路程、平均速度、进入目标象限次数等水迷宫行为学指标、海马组织生理病理结构变化及海马中神经递质含量,评价酸枣仁总皂苷改善PCPA失眠引起的大鼠学习记忆减退作用,结合代谢组学技术多角度探索酸枣仁总皂苷改善学习记忆损伤的可能机制。
1 材料
1.1 动物
60只SPF级雄性SD大鼠,8周龄,体质量(200±20)g,购自北京维通利华实验动物技术有限公司,动物许可证号SCXK(京)2016-0011。动物饲养于湿度(50±10)%、温度(25±2)℃、12 h昼夜循环的环境中,自由进食饮水,给药前禁食12 h。动物实验经山西中医药大学动物伦理委员会批准(批准号2021DW172)。
1.2 药品与试剂
酸枣仁总皂苷为本课题组制备,质量分数为45.75%[38];PCPA(质量分数>98%,批号GK01-QHLL)购自梯希爱(上海)生化科技股份有限公司;戊巴比妥钠(批号P11011)购自德国默克公司;地西泮(批号1901023203)购自山东信谊制药有限公司;羧甲基纤维素钠(批号J0713A)购自大连美仑生物科技有限公司;生理盐水(批号1901023203)购自邦标医疗科技有限公司;5-HT ELISA试剂盒(批号20211110)、GABA ELISA试剂盒(批号20211105)、NO ELISA试剂盒(批号2021021)购自武汉贝茵莱生物科技有限公司;质谱级乙腈、甲醇和甲酸购自美国Thermo Fisher Scientific公司。
1.3 仪器
U3000型超高效液相色谱仪、ExactiveTMOrbitrap质谱仪、88880018型涡旋仪、DNA120-230型离心浓缩仪(美国Thermo Fisher Scientific公司);CPA225D型十万分之一分析天平(德国Sartorius公司);MMT-200型大鼠水迷宫测试仪(成都泰盟软件有限公司);OFT-100型自主活动测试仪(成都泰盟科技有限公司)。
2 方法
2.1 酸枣仁总皂苷改善失眠大鼠学习记忆药效学评价
2.1.1 PCPA失眠模型的建立
(1)PCPA悬浮液的制备:用pH 7~8的碱性生理盐水配制200 mL 0.5%羧甲基纤维素钠溶液,少量多次地加入PCPA粉末中,不断搅拌,制成质量浓度为40 mg/mL的悬浮液。
(2)PCPA失眠大鼠模型的复制:取48只SD大鼠,每日8: 00—9: 30时ip PCPA溶液(400 mg/kg),连续注射4 d。另取12只大鼠作为对照组。造模第3天眼眶取血,每组随机取6只。血样于4 ℃静置30 min,3500 r/min离心10 min,得血清样品,按照ELISA试剂盒说明书测定正常大鼠与造模后大鼠血清中5-HT水平,模型大鼠血清中5-HT水平显著低于正常大鼠(<0.001),表明PCPA大鼠失眠模型制备成功。
2.1.2 分组及给药 造模后的大鼠随机分为模型组、地西泮(2 mg/kg)组和酸枣仁总皂苷低、高剂量(10、20 g/kg)组,每组12只。每日8: 00—9: 00时,各给药组ig相应药物(20 mL/kg),对照组和模型组ig等体积生理盐水,1次/d,连续7 d。
2.1.3 旷场实验 给药第3天进行旷场实验,在给药30 min后将大鼠放置于中心区域进行测试,测试时间6 min。前1 min为适应期,记录5 min内大鼠总路程和静止时间。
2.1.4 Morris水迷宫实验 给药第4天进行Morris水迷宫实验。实验在黑色内壁的圆形水池中进行,水池等分为4个象限(平面直角坐标系中里的横轴和纵轴所划分的4个区域,右上的称为第I象限,左上的称为第II象限,左下的称为第III象限,右下的称为第IV象限),目标平台位于第IV象限水面下1 cm。水温恒定(22±2)℃,光线恒定,无日光变化影响。实验过程采用视频跟踪分析系统进行记录。
(1)定位航行实验:选择离目标平台的最远方位(第II象限)固定位点将大鼠面朝内壁放入水中,记录各组大鼠从进入水中到找到并爬上目标平台停留15 s所用的时间,即大鼠的逃避潜伏期。若大鼠在90 s内没有找到目标平台,则对其进行引导,并在平台上停留15 s,所有大鼠每天训练2次,持续2 d。
(2)空间探索实验:撤去平台,将各组大鼠在同一位置面向池内壁放入水中,记录大鼠在60 s内行动轨迹图、游泳速度及其穿越目标象限的次数、时间、路程。
2.1.5 大鼠取材与处理 末次给药1 h后,各组大鼠ip 10%戊巴比妥钠麻醉后处死,断头,经颅前正中线剪开头皮,小心撬开颅骨,在冰盒上取下大鼠脑组织,剥离海马,放入4%多聚甲醛固定液中,每组3份。经石蜡切片后进行苏木素-伊红(HE)染色,在光学显微镜下对海马组织进行病理观察并采集图像。各组剩余大鼠海马样品,称定质量后加入10倍量生理盐水,4 ℃匀浆180 s,3000 r/min离心15 min,收集上清液,按照ELISA试剂盒说明书测定海马组织中5-HT、GABA和NO水平。
2.2 海马组织代谢组学分析
2.2.1 海马组织样品的处理 精密称取“2.1.5”项下的对照组、模型组、地西泮组和酸枣仁总皂苷低剂量组大鼠海马组织30 mg,加入300 μL预冷的含0.1%甲酸的甲醇溶液,4 ℃匀浆3 min(频率60 Hz),4 ℃离心10 min,取上清液250 μL置于离心浓缩仪中浓缩至干。加入100 μL 70%甲醇水溶液复溶,涡旋使完全溶解,4 ℃、13 000 r/min离心5 min,上清液过0.22 μm微孔滤膜进行分析。
2.2.2 质控样品的处理 取20 μL各组海马组织样品溶液混合作为质控样品。质控样品在运行之前、样品组间和运行之后进样,以评估整个分析过程的稳定性和重现性。
2.2.3 色谱条件 Acquity UPLC®HSS T3色谱柱(100 mm×2.1 mm,1.8 μm),流动相为0.1%甲酸水溶液(A)-乙腈(B),梯度洗脱:0~1 min,2% B;1~3 min,2%~50% B;3~14 min,50%~60% B;14~18 min,60%~90% B;18~19 min,90%~100% B;19~20 min,100% B;20~22 min,100%~2% B。柱温50 ℃;体积流量0.35 mL/min;进样量2 μL。
2.2.4 质谱条件 ESI离子源;正、负离子扫描;扫描模式Full MS/dd MS2;鞘气体积流量35 arb(ESI+)、35 arb(ESI−);辅助气体积流量10 arb(ESI+)、10 arb(ESI−);喷雾电压3.5 kV(ESI+)、2.5 kV(ESI−);毛细管温度320 ℃;离子源温度350 ℃(ESI+)、300 ℃(ESI−);一级全扫描分辨率70 000、二级扫描分辨率为17 500;扫描范围/100~1500;动态排除持续时间8 s;NCE能量分别设为20、30、50 V。
2.3 代谢组学数据处理
UPLC-MS数据收集完毕后,导出raw格式文件,采用Compound Discoverer 3.3软件进行图谱对比查看、去噪音、解卷积和归一化等数据预处理,导出csv格式的二维数据矩阵,包括/、保留时间和峰面积。使用SIMCA-P 14.1软件对处理数据依次进行主成分分析(principal component analysis,PCA)、正交偏最小二乘判别分析(orthogonal partial least squares discriminant analysis,OPLS-DA),根据VIP>1及<0.05筛选出潜在代谢标记物,将筛选的差异代谢物导入Metabo Analyst 5.0数据库进行高通量代谢通路分析。
2.4 统计分析

3 结果
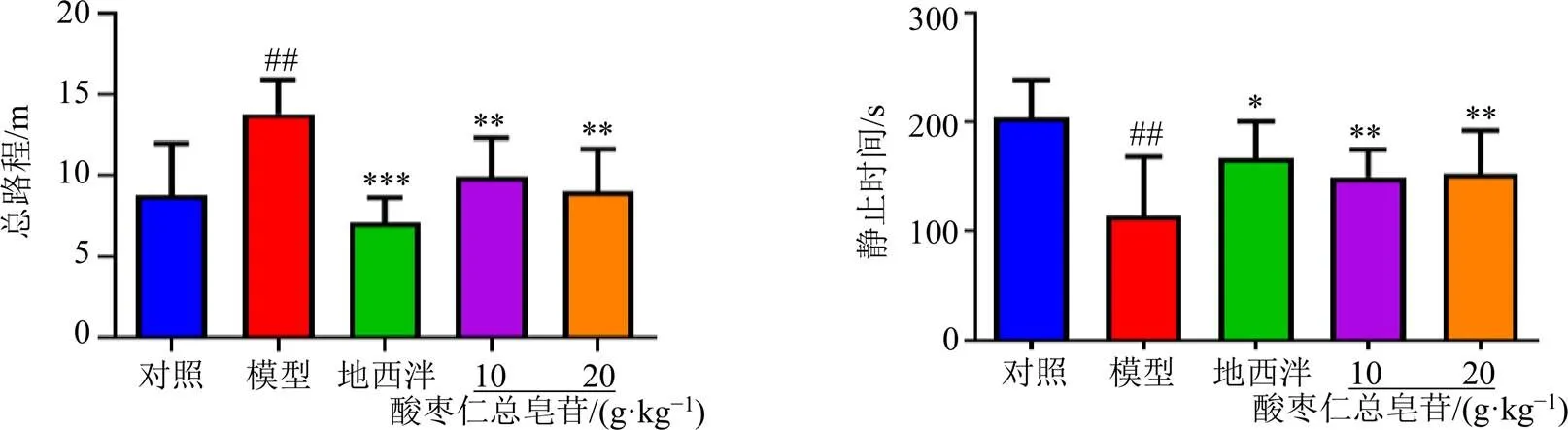
3.1 旷场实验结果
如图1所示,与对照组比较,模型组大鼠总路程显著增加(<0.01),静止时间显著降低(<0.05);与模型组比较,各给药组大鼠总路程显著降低(<0.01、0.001),静止时间显著升高(<0.05、0.01),表明酸枣仁总皂苷具有镇静催眠的作用。
3.2 Morris水迷宫行为学实验结果
3.2.1 定位航行实验结果 以逃避潜伏期为指标评价失眠大鼠空间学习能力。在经过4次的引导训练后,各组大鼠逃避潜伏期均有不同程度的下降,第4次训练即最终定位航行测试结果,见图2。与对照组比较,模型组大鼠逃避潜伏期显著增加(<0.05);与模型组比较,地西泮组和酸枣仁总皂苷低剂量组大鼠逃避潜伏期显著降低(<0.05、0.01)。
与对照组比较:#P<0.05 ##P<0.01 ###P<0.001;与模型组比较:*P<0.05 **P<0.01 ***P<0.001,下图同
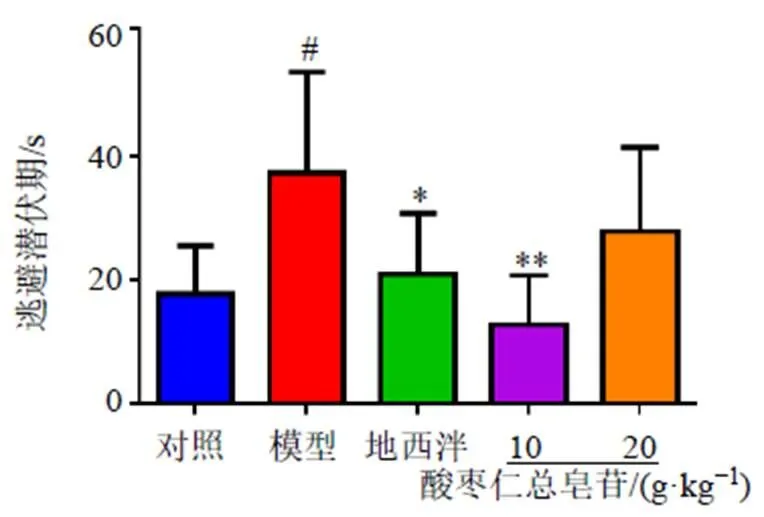
图2 酸枣仁总皂苷对失眠大鼠定位巡航能力的影响(, n = 8)
3.2.2 空间探索实验 采用大鼠水迷宫测试仪跟踪成像系统对各组大鼠在水池中的运动轨迹进行记录,见图3。原平台所在象限为第IV象限,对照组大鼠入水后能够自主地去第IV象限寻找平台,轨迹较为松散;模型组大鼠的轨迹集中分布在入水的第I象限内,较少去探索其他象限;地西泮组大鼠入水后活动减少,出现静止漂浮水面的现象,个别大鼠在水中无规律的运动;酸枣仁总皂苷低、高剂量组大鼠在平台所在的第IV象限内徘徊,并在入水后有直线进入第IV象限的趋势,提示给予酸枣仁总皂苷后,大鼠学习记忆能力有所回调。
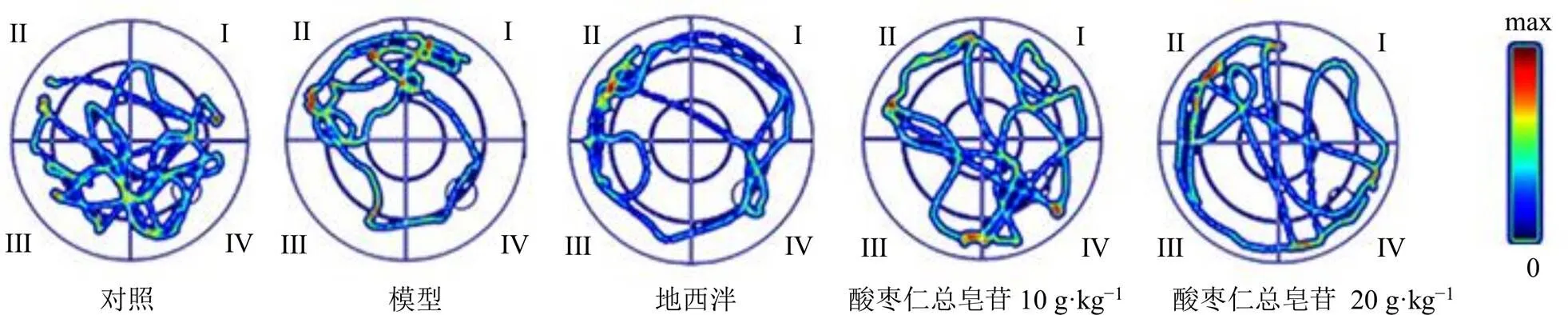
图3 各组大鼠Morris水迷宫运动轨迹图
利用GraphPad Prism软件对目标象限停留时间、进入目标象限路程、平均速度和进入目标象限次数4个指标进行分析。如图4所示,与对照组比较,模型组大鼠在撤去平台后进入第IV象限时间、路程、次数及平均速度均显著降低(<0.01),在实验中观察到其多在入水处徘徊或无目的的探索情况;与模型组比较,各给药组大鼠进入目标象限的路程、停留时间、次数及平均速度均显著延长(<0.05、0.01),表明酸枣仁总皂苷能够改善失眠大鼠的学习记忆能力。
3.3 大鼠海马病理切片结果
如图5所示,模型组大鼠海马组织出现明显损伤,在海马齿状回区胞质空泡化严重,同时神经元细胞出现皱缩,染色加深,胞核胞质分界不清,可能影响到信号的传导及神经元的产生。与模型组相比较,各给药组大鼠海马胞质空泡化损伤均有不同程度改善,但仍存在神经元萎缩现象,酸枣仁总皂苷低剂量组大鼠海马切片较为接近对照组,与行为学结果一致。
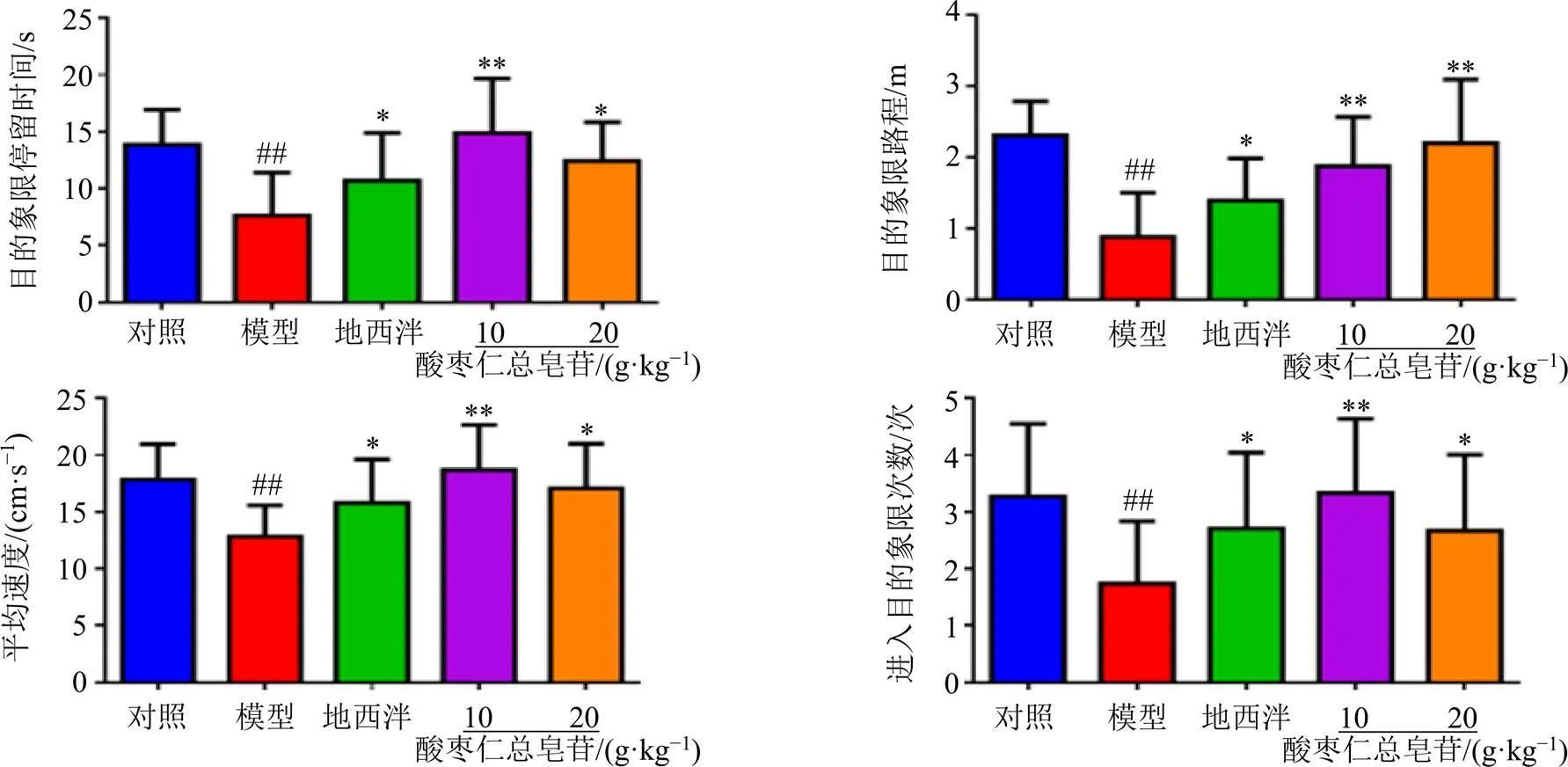
图4 酸枣仁总皂苷对失眠大鼠空间探索能力的影响(, n = 8)

黑色箭头表示胞核胞质分界不清,黄色箭头表示神经元胞质空泡化
3.4 酸枣仁总皂苷对失眠大鼠海马组织中5-HT、GABA和NO水平的影响
如图6所示,与对照组比较,模型组大鼠海马组织中5-HT、GABA和NO水平均显著降低(<0.01);与模型组比较,各给药组海马组织中5-HT、GABA和NO水平均显著升高(<0.05、0.01)。
3.5 海马代谢组学研究结果
3.5.1 海马组织样品液质轮廓分析 为考察各组大鼠体内代谢组学的整体变化,对原始数据进行预处理,采用SIMCA-P软件对海马组织样品进行PCA分析,在正、负离子模式下质控样品中分别有100%、99%峰的RSD值<30%,表明该方法有较好的稳定性及重复性,数据可靠。如图7所示,正、负离子模式下对照组与模型组明显分开,且组间距离较远,表明模型组与对照组大鼠的内源性代谢物存在较大差异,而各给药组均可回调这种差异。
3.5.2 潜在标志物预测分析 为了对比酸枣仁总皂苷对大鼠海马组织样品内源性代谢物的影响,进一步对对照组和模型组、模型组和酸枣仁总皂苷低剂量组进行有监督的OPLS-DA分析,结合VIP>1、<0.05筛选潜在生物标志物的候选变量。图8-A、B为对照组和模型组的OPLS-DA图,2组可沿X轴明显分开,组间差异较大,表明模型组大鼠代谢轮廓发生改变,即在正、负离子模式下,ip PCPA显著影响了大鼠海马中的内源性代谢物。200次模型验证结果表明(正离子模式下2=0.996、2=0.954,负离子模式下2=0.996、2=0.967),模型可靠,具有良好的预测能力,可用于后续差异代谢物的筛选。S-plot图中红色点即为VIP>1的代谢物,表明这些代谢物对组间贡献度较大,可能是两组间的差异代谢物。同时对模型组和酸枣仁总皂苷低剂量组进行OPLS-DA分析,结果见图8-C、D。
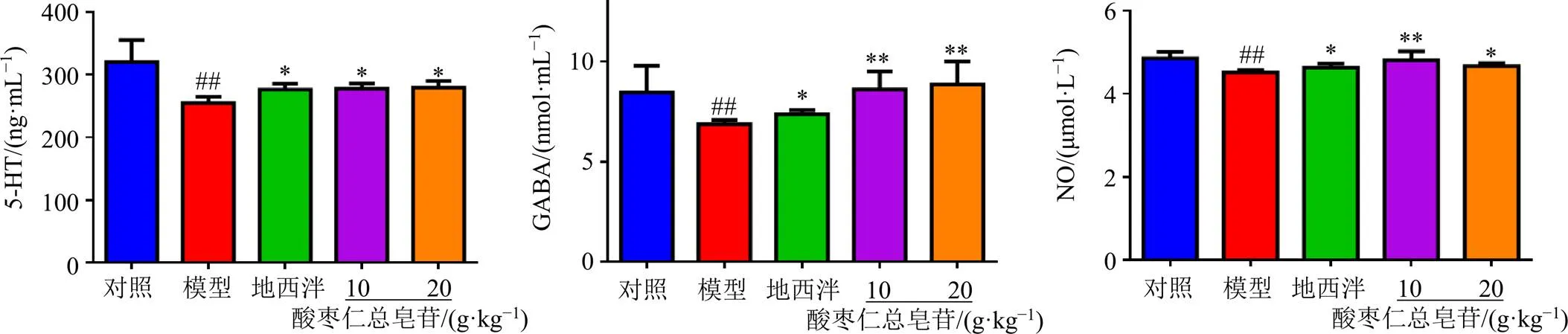
图6 酸枣仁总皂苷对失眠大鼠海马组织中5-HT、GABA和NO水平的影响(, n = 6)

图7 正 (A)、负(B) 离子模式下各组大鼠海马样品PCA分析

图8 正 (A, C)、负离子 (B, D) 模式下各组大鼠海马样品的OPLS-DA得分图、模型验证图及S-plot图
3.5.3 海马组织样品差异代谢物的确定 将筛选得到的差异性代谢物与mzcloud、mzvault、HMDB等数据库对化合物信息进行比对,共鉴定出18个差异代谢物。为了进一步分析模型组及各给药组海马组织样品中18个内源性差异代谢物的动态变化规律,将各组差异性代谢物的峰面积数据导入Graphpad软件绘制箱图,结果见图9。与对照组比较,模型组10种氨基酸类(精氨酸、苯丙氨酸、脯氨酸、蛋氨酸、酪氨酸、赖氨酸、色氨酸、组氨酸、焦谷氨酸、苏氨酸)及2种酰胺类(谷氨酰胺、油酰胺)和其他类(二苄胺、胆碱、肌苷、胞嘧啶)共16种代谢物含量均显著降低(<0.05、0.01、0.001),十六酰胺、十八烯酸2种差异代谢物含量显著升高(<0.001);与模型组比较,酸枣仁总皂苷低剂量组10种氨基酸、2种酰胺类代谢物和肌苷、胞嘧啶共14种差异代谢物含量显著上调(<0.05、0.01、0.001),十六酰胺、十八烯酸2种差异代谢物含量显著下调(<0.05、0.001)。
3.5.4 代谢通路分析 将酸枣仁总皂苷显著回调的16种差异代谢物(精氨酸、苯丙氨酸、脯氨酸、蛋氨酸、酪氨酸、赖氨酸、色氨酸、组氨酸、焦谷氨酸、苏氨酸、油酰胺、谷氨酰胺、肌苷、胞嘧啶、十六酰胺、十八烯酸)分别导入MetaboAnalyst 5.0数据处理中心进行京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)代谢通路富集和网络构建,基于拓扑分析进行权重计算,通过对值、Holm、误判率(FDR)及通路影响值的综合分析,得到代谢通路富集气泡图,结果见图10。酸枣仁总皂苷可通过7条主要代谢通路调控大鼠海马代谢,最重要的是苯丙氨酸、酪氨酸和色氨酸生物合成代谢通路,其通路影响值可达到1。其余比较关键的代谢通路按通路影响值由高到低排列依次为苯丙氨酸代谢、精氨酸生物合成、精氨酸和脯氨酸代谢、组氨酸代谢、半胱氨酸和蛋氨酸代谢以及丙氨酸、天冬氨酸和谷氨酸代谢等。本研究中所涉及到的代谢通路均是氨基酸代谢通路,提示酸枣仁总皂苷能够调控海马组织中氨基酸代谢水平。

图9 各组大鼠海马样品差异代谢物的相对含量(, n = 6)
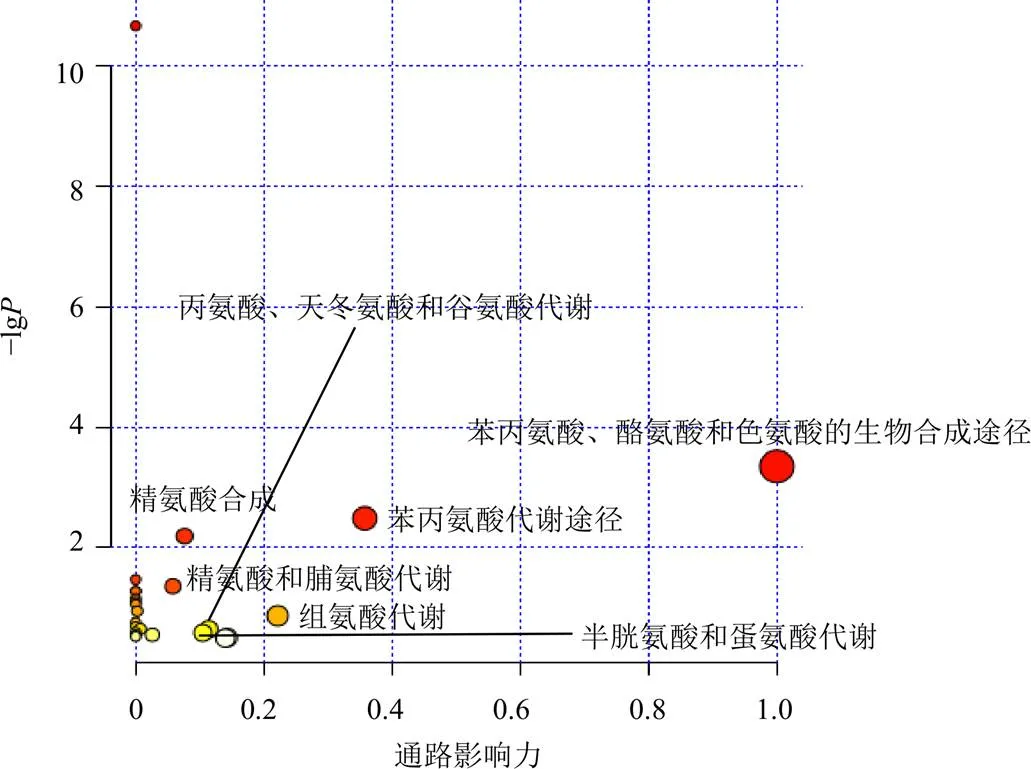
图10 酸枣仁总皂苷调控的代谢通路图
代谢组学结果表明,酸枣仁总皂苷能够通过多种途径调控失眠大鼠体内异常代谢,主要涉及到7条主要相关氨基酸代谢通路,为了进一步明确酸枣仁总皂苷在体内代谢机制,结合KEGG及相关富集分析方法,绘制代谢途径网络图,见图11。
4 讨论
本研究建立PCPA大鼠失眠模型,利用旷场及Morris水迷宫等行为学实验评价酸枣仁总皂苷对学习记忆损伤的药效作用,采用HE染色法观察大鼠海马组织病理变化,ELISA测定海马组织中5-HT、GABA及NO水平,并结合代谢组学技术对大鼠海马组织代谢轮廓进行分析,采用多元统计分析筛选差异性代谢物,基于MetaboAnalyst 5.0数据库进行代谢通路及代谢网络分析,从多角度探索酸枣仁总皂苷改善失眠大鼠学习记忆损伤的作用途径。
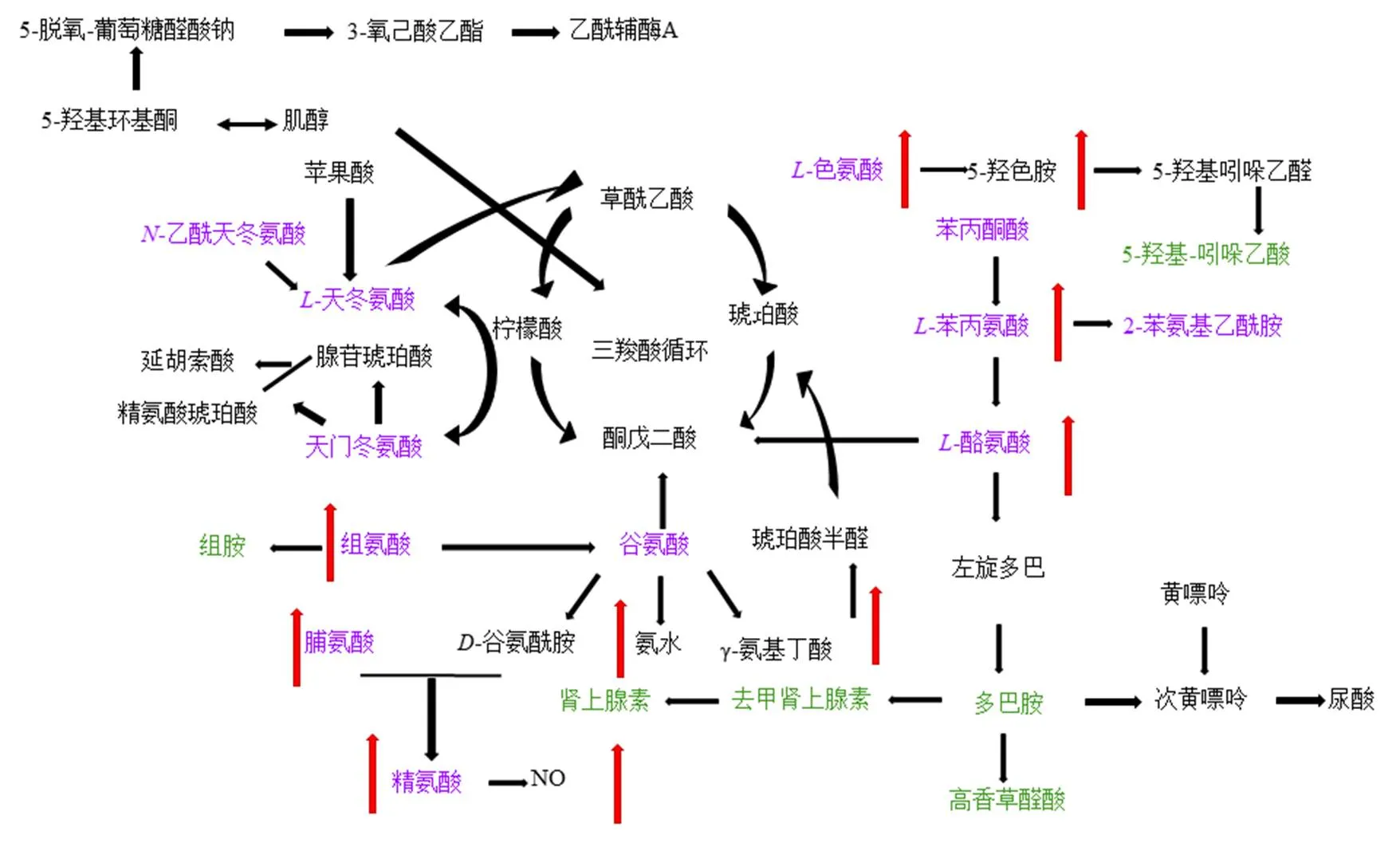
紫色为氨基酸,绿色为神经递质及激素类物质,红色箭头为酸枣仁总皂苷显著上调的内源性物质
研究表明,慢波睡眠与快速眼动睡眠能够巩固学习记忆的形成,且与神经递质水平的变化有关[41]。5-HT能够参与机体睡眠-觉醒过程,并且与动物觉醒呈正相关,对慢波睡眠的发生和维持起着重要作用[42],5-HT水平升高有利于维持睡眠状态[43],本研究结果显示,酸枣仁总皂苷可通过上调海马组织中5-HT水平维持睡眠状态。GABA是抑制性神经递质,可阻断中枢神经兴奋传导和过度放电,当其表达下降时,促觉醒核团功能增强,诱发觉醒[44]。酸枣仁总皂苷能够显著上调海马组织中GABA水平,从而发挥改善学习记忆的作用。NO能够调节神经递质释放、突触可塑性的发育,与学习记忆的形成过程有着紧密的联系[45-46]。本研究表明酸枣仁总皂苷可显著回调海马组织中5-HT、GABA与NO水平,从而发挥直接改善学习记忆损伤的作用。
本研究发现,低、高剂量的酸枣仁总皂苷均能显著减少大鼠旷场实验的总路程,延长静止时间,增加水迷宫指标中目标象限停留时间、平均速度及进入次数,提高海马组织中5-HT、GABA、NO的水平,但低剂量组效果稍优于高剂量组,为非线性量-效关系。中药的内在成分极其复杂,不符合中药剂量-效应关系,可能是中药各种成分之间产生拮抗作用的结果,从而降低其药理活性[47]。此外,中药通常还具有双向调节的作用,低剂量具有促进效应,高剂量具有抑制效应。研究表明,GABAA受体参与调节学习记忆[48],酸枣仁皂苷B能激活GABAA受体的表达[49],从而改善学习记忆,但高剂量的酸枣仁皂苷B会使GABAA过表达,造成学习记忆损伤[50]。因此,酸枣仁总皂苷改善学习记忆损伤未呈现量效关系可能是上述原因造成的。
苯丙氨酸、酪氨酸和色氨酸的生物合成途径是酸枣仁总皂苷调控海马组织中最为主要的一条代谢途径。苯丙氨酸和酪氨酸与体内神经递质水平息息相关,苯丙氨酸可以经过苯丙氨酸羟化酶的作用合成酪氨酸[51],进而产生多巴胺、去甲肾上腺素以及肾上腺素,这些兴奋性神经递质均密切参与睡眠-觉醒过程,是引起失眠的重要神经递质[52-53]。色氨酸是参与控制神经中枢的重要代谢物,可由TPH催化合成重要的神经递质5-HT[54]。本研究结果显示,酸枣仁总皂苷可能通过苯丙氨酸、酪氨酸和色氨酸的生物合成途径,调控苯丙氨酸、酪氨酸和色氨酸等氨基酸及代谢途径的中间产物多巴胺、去甲肾上腺素等神经递质水平改善失眠,进而改善模型动物的学习记忆损伤。
精氨酸与脯氨酸代谢途径是调控学习记忆的重要途径[55]。精氨酸是NO的直接前体,可促进脑内神经元型一氧化氮合酶、c-Fos的表达及新生神经元的生成,增强学习记忆能力。NO作为重要的信号分子,介导、参与学习记忆相关的特定生理过程[56-57]。脯氨酸是组成酪氨酸激酶2(proline-rich tyrosine kinase 2,Pyk2)的重要氨基酸。Pyk2通过激活蛋白酪氨酸激酶通路,增强-甲基--天冬氨酸受体活性,使蛋白质磷酸化增强,促使钙离子大量流入突触后神经元,因而产生较强的兴奋性突触后电位,影响海马突触可塑性,进而改善记忆损伤[58]。本研究中酸枣仁总皂苷能够显著回调海马组织中NO水平,上调精氨酸和脯氨酸的水平,进而改善失眠引起的记忆损伤。组氨酸代谢途径是维持脑内组胺稳态的重要途径[59]。组氨酸是组胺的前体,组胺是促进觉醒和维持觉醒状态的一种兴奋性神经递质。研究表明,下丘脑后部的结节乳头体核内组胺能神经元具有促进和维持觉醒的作用[60]。组胺也可以通过增强基底核的胆碱能神经的活性,增加海马内胆碱含量而改善认知能力[61]。本研究表明酸枣仁总皂苷可以显著上调海马组织中组氨酸水平,从而改善学习记忆损伤。
综上所述,本研究采用PCPA复制失眠大鼠模型,整合药效学和代谢组学的方法,从整体水平上探讨了酸枣仁总皂苷改善失眠导致学习记忆损伤的作用机制。酸枣仁总皂苷能够显著回调水迷宫行为学指标,改善失眠引起的大鼠记忆损伤,并使海马胞质空泡化损伤恢复,显著上调海马组织中5-HT、GABA、NO水平,进而改善学习记忆损伤。此外,酸枣仁总皂苷能够恢复PCPA引起的海马中氨基酸代谢紊乱和代谢轮廓异常,进而发挥改善学习记忆损伤的作用。本研究在代谢层面发现调控学习记忆的氨基酸通路,并提供了代谢物类别和相对含量层面的信息。失眠模型动物海马代谢谱的改变对研究失眠模型下的学习记忆损伤过程具有重要价值,但其代表的生物学意义仍需要谨慎解释和验证,需进一步在细胞、蛋白等分子水平上深入研究。本研究将为进一步深入探讨酸枣仁总皂苷改善PCPA失眠大鼠学习记忆提供研究基础,为酸枣仁总皂苷的新药创制研究提供科学参考。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典 [S]. 一部. 2020: 382.
[2] KIM J H. Chemical compounds in the seeds ofvar.[J]., 2016, 4(1): 13-28.
[3] Zhang F X, Li M, Qiao L R,. Rapid characterization ofby UPLC/Qtof MS with novel informatics platform and its application in evaluation of two seeds fromspecies [J]., 2016, 122: 59-80.
[4] 李旭, 和建政, 陈彻, 等. 酸枣仁镇静催眠活性成分及药理作用研究进展 [J]. 中华中医药学刊, 2022, 40(2): 23-31.
[5] 任晓宇, 李廷利. 酸枣仁汤对慢性睡眠剥夺小鼠肝功能和特定肠道菌的影响[J]. 药物评价研究, 2020, 43(2): 226-231.
[6] You Z L, Xia Q, Liang F R,. Effects on the expression of GABAA receptor subunits by jujuboside A treatment in rat hippocampal neurons [J]., 2010, 128(2): 419-423.
[7] Manayi A, Nabavi S M, Daglia M,. Natural terpenoids as a promising source for modulation of GABAergic system and treatment of neurological diseases [J]., 2016, 68(4): 671-679.
[8] Wang X X, Ma G J, Xie J B,. Influence of JuA in evoking communication changes between the small intestines and brain tissues of rats and the GABAA and GABAB receptor transcription levels of hippocampal neurons [J]., 2015, 159: 215-223.
[9] Liang Y, Yang X, Zhang X J,. Antidepressant-like effect of the saponins part of ethanol extract from SHF [J]., 2016, 191: 307-314.
[10] Fu Q, Yuan H M, Chen J,. Dammarane-type saponins fromjujube and their inhibitory effects against TNF-α release in LPS-induced RAW246.7 macrophages [J]., 2016, 16: 169-173.
[11] 杜晨晖, 裴香萍, 张敏, 等. 基于1H-NMR代谢组学的酸枣仁改善失眠大鼠睡眠作用机制研究[J]. 中草药, 2019, 50(10): 2405-2412.
[12] 王俊秀. 中国睡眠研究报告 [M]. 北京: 社会科学文献出版社, 2022: 11.
[13] Cordeira J, Kolluru S S, Rosenblatt H,. Learning and memory are impaired in the object recognition task during metestrus/diestrus and after sleep deprivation [J]., 2018, 339: 124-129.
[14] 李雪丽, 张斌. 睡眠障碍与焦虑抑郁障碍的联系 [J]. 中国临床医生杂志, 2018, 46(2): 131-133.
[15] Lopez J, Roffwarg H P, Dreher A,. Rapid eye movement sleep deprivation decreases long-term potentiation stability and affects some glutamatergic signaling proteins during hippocampal development [J]., 2008, 153(1): 44-53.
[16] 刘雪平, 郑敏, 郝跃伟, 等. 吡格列酮对胰岛素抵抗大鼠认知功能及脑组织AGE-RAGE损伤途径的影响 [J]. 山东大学学报: 医学版, 2008, 46(10): 954-958.
[17] Gupta Y K, Gupta M, Kohli K. Neuroprotective role of melatonin in oxidative stress vulnerable brain [J]., 2003, 47(4): 373-386.
[18] Wadhwa M, Prabhakar A, Ray K,. Inhibiting the microglia activation improves the spatial memory and adult neurogenesis in rat hippocampus during 48 h of sleep deprivation [J]., 2017, 14(1): 222.
[19] 魏文静, 仝立国, 仲启明, 等. 蛇床子催眠活性组分对对氯苯丙氨酸致失眠大鼠海马钟基因与氨基酸类神经递质表达的影响 [J]. 中草药, 2018, 49(11): 2614-2619.
[20] Yan X D, Hu G Y, Yan W M,. Ginsenoside Rd promotes non-amyloidogenic pathway of amyloid precursor protein processing by regulating phosphorylation of estrogen receptor alpha [J]., 2017, 168: 16-23.
[21] 杨岩涛, 赵佳柱, 肖佳妹, 等. 人参皂苷治疗阿尔茨海默病的药理作用研究进展 [J]. 中国药理学通报, 2021, 37(12): 1638-1643.
[22] Kopra J J, Panhelainen A, Af Bjerkén S,. Dampened amphetamine-stimulated behavior and altered dopamine transporter function in the absence of brain GDNF [J]., 2017, 37(6): 1581-1590.
[23] Yan S J, Li Z, Li H,. Notoginsenoside R1increases neuronal excitability and ameliorates synaptic and memory dysfunction following amyloid elevation [J]., 2014, 4: 6352.
[24] 范军朝, 宋俊杰, 陈勇. 酸枣仁皂苷缓解阿尔茨海默病模型小鼠的机制研究 [J]. 中国比较医学杂志, 2020, 30(9): 32-37.
[25] 朱洁, 蔡玉芳, 袁红卫, 等. 酸枣仁皂苷对Alzheimer病大鼠认知功能障碍改善的实验研究 [J]. 江西中医药大学学报, 2017, 29(5): 74-76.
[26] 全世建, 林杏娥, 刘妮. PCPA大鼠失眠模型的证候属性研究 [J]. 中医药学刊, 2006, 24(3): 450-451.
[27] 任俊华. 基于质谱分析技术的针刺对失眠模型大鼠脑组织蛋白差异表达的研究 [D]. 郑州: 河南中医学院, 2014.
[28] Borbély A A, Neuhaus H U, Tobler I. Effect of-chlorophenylalanine and tryptophan on sleep, EEG and motor activity in the rat [J]., 1981, 2(1): 1-22.
[29] 夏天吉, 闫明珠, 王智, 等. 大小鼠失眠模型和评价方法研究进展 [J]. 中国实验动物学报, 2022, 30(3): 428-435.
[30] Nieto R, Kukuljan M, Silva H. BDNF and schizophrenia: From neurodevelopment to neuronal plasticity, learning, and memory [J]., 2013, 4: 45.
[31] Hua Y, Guo S, Xie H,.mill. var.(Bunge) Hu ex H. F. Chou seed ameliorates insomnia in rats by regulating metabolomics and intestinal flora composition [J]., 2021, 12: 653767.
[32] Yan Y, Li Q, Du H Z,. Determination of five neurotransmitters in the rat brain for the study of the hypnotic effects ofaqueous extract on insomnia rat model by UPLC-MS/MS [J]., 2019, 17(7): 551-560.
[33] Murray N M, Buchanan G F, Richerson G B. Insomnia caused by serotonin depletion is due to hypothermia [J]., 2015, 38(12): 1985-1993.
[34] Persico A M, Altamura C, Calia E,. Serotonin depletion and barrel cortex development: Impact of growth impairment vs. serotonin effects on thalamocortical endings [J]., 2000, 10(2): 181-191.
[35] 王金鑫, 惠洋, 高旭. 成体海马神经再生与神经退行性疾病 [J]. 国际遗传学杂志, 2018, 41(3): 207-211.
[36] Díaz A, de Jesús L, Mendieta L,. The amyloid-beta25-35 injection into the CA1 region of the neonatal rat hippocampus impairs the long-term memory because of an increase of nitric oxide [J]., 2010, 468(2): 151-155.
[37] 郭杰, 尹晓刚. 酸枣仁汤对老年失眠模型大鼠学习记忆能力及脑内神经递质含量的影响 [J]. 中国药房, 2016, 27(22): 3085-3087.
[38] 孟胜喜, 霍清萍, 王兵, 等. 恒清II号方对阿尔茨海默病小鼠认知功能和粪便代谢组学影响研究 [J]. 现代中西医结合杂志, 2020, 29(24): 2628-2634.
[39] 邵晓妮, 黑亚南, 孙英凯, 等. 高尿酸血症认知功能障碍模型大鼠皮层和海马的代谢组学研究 [J]. 中国药理学通报, 2021, 37(2): 196-202.
[40] 崔小芳. 酸枣仁总皂苷的肠道菌群体外代谢转化研究 [D]. 太原: 山西中医药大学, 2020.
[41] Venner A, de Luca R, Sohn L T,. An inhibitory lateral hypothalamic-preoptic circuit mediates rapid arousals from sleep [J]., 2019, 29(24): 4155-4168.e5.
[42] 胡文卓, 贾力莉, 马澜, 等. 佛手柑内酯对PCPA失眠大鼠神经递质及学习记忆的影响 [J]. 时珍国医国药, 2020, 31(4): 821-823.
[43] 王婷婷, 郝蕾, 王芮, 等. 酸枣仁-延胡索配伍对肝郁型失眠小鼠的作用及机制研究 [J]. 中药新药与临床药理, 2021, 32(5): 619-626.
[44] 刘静, 郭欣, 黄娜娜, 等. 柴胡桂枝干姜汤治疗失眠的功效网络研究[J]. 中草药, 2019, 50(21): 5145-5153.
[45] Moosavi M, Abbasi L, Zarifkar A,. The role of nitric oxide in spatial memory stages, hippocampal ERK and CaMKII phosphorylation [J]., 2014, 122: 164-172.
[46] Sestakova N, Puzserova A, Kluknavsky M,. Determination of motor activity and anxiety-related behaviour in rodents: Methodological aspects and role of nitric oxide [J]., 2013, 6(3): 126-135.
[47] 何婷, 赵薇, 华国栋. 从中药量效关系论临床用药剂量的适当性 [J]. 现代中医临床, 2014, 21(6): 37-39.
[48] Liu Y, Zong X D, Huang J,. Ginsenoside Rb1regulates prefrontal cortical GABAergic transmission in MPTP-treated mice [J]., 2019, 11(14): 5008-5034.
[49] Song P P, Zhang Y, Ma G J,. Gastrointestinal absorption and metabolic dynamics of jujuboside A, A saponin derived from the seed of[J]., 2017, 65(38): 8331-8339.
[50] Amani M, Mohammadian F, Golitabari N,. Postnatal GABA A receptor activation alters synaptic plasticity and cognition in adult wistar rats [J]., 2022, 59(6): 3585-3599.
[51] 邓媛媛, 王军, 姚佳希, 等. 双酚S暴露对小鼠脑-肠-菌轴神经递质代谢稳态的影响 [J]. 南京医科大学学报: 自然科学版, 2020, 40(5): 663-668.
[52] Xiao F, Du Y, Lv Z Q,. Effects of essential amino acids on lipid metabolism in mice and humans [J]., 2016, 57(4): 223-231.
[53] Trushina E, Dutta T, Persson X M T,. Identification of altered metabolic pathways in plasma and CSF in mild cognitive impairment and Alzheimer’s disease using metabolomics [J]., 2013, 8(5): e63644.
[54] 周恩, 朱磊, 沈洪. 中医药对色氨酸代谢的研究进展 [J]. 中医药临床杂志, 2022, 34(3): 580-585.
[55] Lv X F, Feng Y Y, Ma R,. Effects of peppermint essential oil on learning and memory ability in APP/PS1transgenic mice [J]., 2022, 27(7): 2051.
[56] 淮盼盼, 赵建强, 王奕, 等. 长时间应用大剂量的左旋精氨酸对幼鼠学习记忆功能和神经系统可塑性的抑制作用 [J]. 山西医科大学学报, 2011, 42(11): 864-868.
[57] 吕梦, 王雅泽, 赵迪, 等. 基于粪便代谢组学技术的逍遥散抗抑郁作用机制研究 [J]. 中草药, 2020, 51(13): 3482-3492.
[58] 杜雨生, 张钊, 何新. 富含脯氨酸的酪氨酸激酶2与突触可塑性 [J]. 中国药理学与毒理学杂志, 2019, 33(10): 860.
[59] Mahmood D. Histamine H3receptors and its antagonism as a novel mechanism for antipsychotic effect: A current preclinical & clinical perspective [J]., 2016, 10(4): 564-575.
[60] Chen Z, Shen Y J. Effects of brain histamine on memory deficit induced by nucleus basalis-lesion in rats [J]., 2002, 23(1): 66-70.
[61] 何超. 组胺促觉醒系统对内嗅皮层神经元活动的调控及空间学习记忆的影响 [D]. 重庆: 第三军医大学, 2015.
Mechanism of total saponins ofon learning and memory impairment in-4-chlorophenylalanine-induced insomnia rats
HENG Yi-ran1, DUAN Hui-zhu1, DU He1, YAN Yan2, DU Chen-hui1
1. School of Chinese Medicine and Food Engineering, Shanxi University of Chinese Medicine, Taiyuan 030619, China 2. Modern Research Center for Traditional Chinese Medicine, Shanxi University, Taiyuan 030006, China
To study the effect of total saponins of Suanzaoren (, SZSS) on endogenous metabolites in insomnia model rats based on metabolomics, and explore the effect and mechanism of SZSS on learning and memory.-4-chlorophenylalanine (PCPA) was used to induce insomnia model in rats, diazepam and SZSS were given for intervention. Open field experiment and Morris water maze experiment were carried out; Hematoxylin-eosin (HE) staining was used to observe the pathological changes of hippocampus; Levels of 5- hydroxytryptamine (5-HT), γ-aminobutyric acid (GABA) and NO (nitric oxide) in hippocampus were measured by ELISA. UPLC-Orbitrap-MS/MS was used to analyze the metabolic profile of hippocampus, combined with multivariate statistical analysis to screen out the different metabolites, and based on MetaboAnalyst 5.0 database, metabolic pathway and metabolic network were analyzed, metabolic characteristics and action ways of SZSS to improve learning and memory were discussed from various angles.Compared with model group, learning ability of rats in SZSS group was significantly improved (< 0.05, 0.01), pathological structure of hippocampus was significantly improved, levels of 5-HT, GABA and NO in hippocampus were significantly increased (< 0.05, 0.01). A total of 18 potential biomarkers related to learning and memory impairment had been identified by metabonomics. SZSS mainly played a role in improving memory impairment through seven major amino acid metabolic pathways such as phenylalanine, tyrosine and tryptophan.SZSS can improve the learning and memory loss caused by insomnia by regulating endogenous differential metabolites in hippocampus and regulating related amino acid metabolic pathways.
total saponins of; insomnia; learning and memory; hippocampus; differential metabolites
R285.5
A
0253 - 2670(2022)21 - 6759 - 12
10.7501/j.issn.0253-2670.2022.21.013
2022-07-15
山西省科技厅中央引导地方科技发展资金项目(YDZJSX2021C025);山西省自然科学基金面上项目(20210302123237,20210302123470);山西省卫生健康委“十大晋药”项目(ZYCZL2020007);山西中医药大学青年科学家培育项目(2021PY-QN-07)
衡依然(1995—),女,硕士研究生,研究方向为中药新产品开发与利用。Tel: 18622138832 E-mail: hengyiran1@163.com
闫 艳,副教授,博士,研究方向为中药质量控制及中药体内过程分析。Tel: (0351)7018379 E-mail: yanyan520@sxu.edu.cn
杜晨晖,教授,博士,研究方向为中药药效物质基础。Tel: (0351)179982 E-mail: dch@sxtcm.edu.cn
[责任编辑 李亚楠]
