荒漠草原凋落物分解过程中降水量对土壤酶活性的影响
2022-11-04韩翠康扬眉余海龙李冰黄菊莹
韩翠 ,康扬眉,余海龙,李冰,黄菊莹
1.宁夏大学生态环境学院,宁夏 银川 750021;2.宁夏大学农学院,宁夏 银川 750021;3.宁夏大学地理科学与规划学院,宁夏 银川 750021
气候变暖影响了全球水文循环,导致降水时空分配格局改变(IPCC,2021)。我国作为全球气候变化的敏感区域之一,降水格局也发生了改变,主要表现为年降水总量变化不明显、极端降水事件(如最长持续干旱日数缩短)显著增加(Wu et al.,2016)。受季风环流以及地形等影响,各地降水呈现出复杂的时空格局(Feng et al.,2015)。就西北地区而言,时间上表现为夏秋两季降水增多、空间上表现为东部生态区降水减少而西部生态区降水增加的趋势(黄小燕等,2015;李明等,2021)。我国西北地区属于典型的干旱半干旱气候区,区域内水资源匮乏,干旱事件频发(李明等,2021)。降水不仅是该区草原生态系统主要的水分来源,还是调节其生态系统功能的重要因素(Ma et al.,2020)。降水改变了分解者活动和有机物质分解速率等土壤养分动力学和生物有效性(李吉玫等,2015),进而对生态系统功能产生影响。因此,在该区草原生态系统开展降水量变化的相关研究,可为深入探究降水格局改变的生态效应提供数据支撑。
土壤酶是地下能量流动和物质循环的重要生物活化剂,在土壤C和养分转化中扮演着重要的角色(Avazpoorl et al.,2019)。氧化还原酶和水解酶是土壤酶学研究中涉及较多的酶类。其中,过氧化氢酶是一种重要的氧化还原酶,其活性大小可以表征土壤腐殖质化强度和有机质转化速率(王理德等,2016);蔗糖酶和脲酶是两种重要的水解酶,直接参与有机质矿化,对维持生物地球化学循环起着重要作用(刘红梅等,2018)。研究表明,降水不仅可以通过刺激植物生长和微生物活动影响酶分泌(Hewins et al.,2016),还可以通过改变土壤水分和养分有效性等影响酶和底物扩散,进而刺激或限制酶活性(陈敏玲等,2016),从而对凋落物分解和养分释放产生重要的影响(Taylor et al.,2017;Akinyemia et al.,2020)。凋落物分解可以调节土壤C固存和养分矿化,是维持生态系统功能的基本过程(Suseela et al.,2014;付琦等,2019)。因此,研究凋落物分解过程中降水量对草原土壤酶活性的影响,可为深入探讨气候变化下脆弱生态系统土壤有机质矿化等过程提供数据支撑。
荒漠草原是我国西北干旱半干旱区主要的草原生态系统类型,所处区域干旱少雨、蒸发量大,对气候变化十分敏感(白永飞等,2020)。宁夏荒漠草原位于毛乌素沙地西南缘,是我国典型的生态脆弱区。尽管该生态系统植被稀少,但在固C释氧、防风固沙等方面提供着不可替代的生态服务功能。目前国内在降水格局改变下草原土壤酶活性及其影响因素等方面已积累了丰富的研究成果,但这些研究主要集中在草甸草原和典型草原(如Ma et al.,2020;闫钟清等,2017;钞然等,2018;柴锦隆等,2019),尚缺乏针对荒漠草原的相关研究,尤其是极端降水量变化下。那么,极端降水量如何影响荒漠草原土壤酶活性?降水量变化下荒漠草原土壤酶活性是否会影响凋落物元素释放等问题都值得我们进行深入的思考。为此,本文基于2014年在宁夏荒漠草原设立的降水量变化野外试验,探讨了 480 d凋落物分解过程中土壤酶活性及其影响因素,分析了土壤酶活性与凋落物元素释放量的关系,以期为深入了解全球变化下干旱半干旱区草原生态系统生物地球化学循环提供基础数据。
1 材料与方法
1.1 研究区概况
野外试验样地设立于 1998年开始围封的宁夏盐池县柳杨堡乡杨寨子村草地内。该区地理位置为37.80°N,107.45°E,海拔约 1367 m,属于典型的黄土高原向鄂尔多斯台地过渡带,具有温带大陆性气候:年平均气温7.7 ℃,1月平均气温-8.9 ℃,7月平均气温 22.5 ℃;年平均降水量和蒸发量分别为289.4 mm和2131.8 mm。降水季节分配不均,主要集中在生长季5—8月(图1)(韩翠等,2022)。土壤结构松散,pH偏高,主要类型和质地分别为灰钙土和沙壤土。植被类型为荒漠草原,群落结构简单,物种组成以一年生或多年生草本为主,优势种为草木樨状黄芪(Astragalus melilotoides)和牛枝子(Lespedeza potaninii),常见种包括猪毛蒿(Artemisia scoparia)、白草(Pennisetum centrasiaticum)、苦豆子(Sophora alopecuroides)和针茅(Stipa capillata)等(黄菊莹等,2018)。
1.2 试验设计
于2014年4月下旬,选择围栏草地内地势平坦且植物组成均匀的地段作为长期降水量变化的野外模拟试验样地。降水量处理以近 50年我国西北地区湿润半湿润区降水量减少而干旱半干旱区降水量增加的趋势为主要依据(Wu et al.,2016),同时兼顾了野外试验的可操作性。降水时间设置以研究区降水的季节分布特征(60%以上集中在5—8月,图 1)和植物的生长规律为主要依据。降水频度参考国内同类研究的处理方法(Xu et al.,2018)。试验采用随机区组设计,设置了5个年降水量变化处理,分别为减少50%,即减雨144.7 mm(W1);减少30%,即减雨86.8 mm(W2);对照,即自然降水量(W3);增加30%,即增雨86.8 mm(W4);增加50%,即增雨144.7 mm(W5)。每个处理重复5次,共25个小区。每个小区面积为8 m×8 m。各小区之间设置2 m宽的缓冲带以减少小区将地表径流和地下渗漏等干扰。

图1 试验期间研究区域月降水量、平均气温和平均风速的变化Figure 1 Variations in monthly precipitation, average air temperature and average wind speed during the experiment in the studied area
于每年 5—8月,采用自制遮雨棚减雨和自制喷灌装置增雨相结合的方法进行降水量变化处理。降水量减少的处理中(W1和W2),每个小区搭建1个南北走向的U形遮雨棚(最高点离地约1.5 m)。雨天依据遮雨频率采用透光率>95%的聚氯乙烯塑料薄膜对遮雨棚遮雨。非雨天敞开塑料薄膜以保持遮雨棚通风、降低棚内温度。试验期间,采用雨量器收集降水量,记录每次降水量、统计总降水量和遮雨量;并依据每次降水情况,于每年8月中旬开始对设定的遮雨频率进行微调,以使实际遮雨量尽可能与理论遮雨量相符。其中,W1处理平均每3次自然降雨中遮雨两次,W2处理平均每5次自然降雨中遮雨两次(表 1)。降水量增加的处理中(W4和W5),将需要补给的降水量换算成喷水量,采用流量计的喷灌装置每两周喷水一次。其他处理方法见项目组前期研究(Na et al.,2019;黄菊莹等,2018)。

表1 2014—2016年各降水量处理实际改变的降水量和接受的总降水量Table 1 The actually altered precipitation and the received precipitation in each treatment during 2014-2016
1.3 样品收集与测定
土壤样品的收集依托项目组前期凋落物分解试验平台进行。即,分别于凋落物分解60 d(2017年1月10日)、120 d(2017年3月17日)、180 d(2017年5月21日)、240 d(2017年8月16日)、360 d(2017年11月11日)和480 d(2018年3月10日),在试验区采用内径为5 cm的土钻收集0—20 cm土壤样品。每个小区内随机取3钻,混匀作为一个样品,从中取出10 g左右装入铝盒中用于土壤含水量的测定。剩余部分过2 mm筛后快速分成两部分:一部分低温冰箱中4 ℃下冷藏保存,两周内完成酶活性、pH、电导率、NO3--N质量分数、NH4+-N质量分数和速效P质量分数的测定;另一部分自然风干后,用于有机C、全N和全P质量分数的测定。其中,过氧化氢酶活性采用高锰酸钾滴定法,蔗糖酶活性采用3, 5-二硝基水杨酸比色法,脲酶活性采用靛酚蓝比色法;pH和电导率分别采用酸度计法和便携式电导率仪;NO3--N和NH4+-N质量分数采用连续流动分析仪(Auto Analyzer 3,SEAL Analytical GmbH,Hanau,Germany),速效P质量分数采用 0.5mol·L-1NaHCO3法;有机 C、全N和全P质量分数分别采用重铬酸钾容量法-外加热法、凯氏定氮法和 HCLO4-H2SO4法(鲍士旦,2000)。
依据从全国温室数据系统中气象数据网站(http://data.sheshiyuanyi.com/WeatherData/)提取的2017年1月—2018年3月气象数据(月降水量、平均气温和平均风速),本文分析了土壤酶活性与气象因子的关系。此外,依据项目组前期收集的2017年1月—2018年3月植物凋落物样品,本文分析了土壤酶活性与植物种凋落物C和 N累积释放量的关系。植物凋落物样品收集、化学测定、元素累积释放量的计算方法见项目组前期研究(韩翠等,2022)。
1.4 数据处理
采用 Microsoft Excel 2016进行数据的初步整理。运用SPSS 17.0进行数据的统计分析:采用双因素方差分析(Two-way ANOVA)比较降水量、分解时间及其交互作用对土壤酶活性的影响;采用单因素方差分析(One-way ANOVA)比较相同分解时间下降水量处理间土壤酶活性及其他指标的差异,探讨相同降水量处理下分解时间间各指标的差异。若方差为齐性,选用最小显著性差异法(LSD),否则选用Tamhane’s T2法。在用Origin 2018中,采用线性回归方程拟合土壤酶活性与植物种凋落物元素累积释放量之间的关系。采用Origin 2018对以上分析结果作图。采用Canoco 5.0进行土壤酶活性与环境因子对应关系的冗余分析(Redundancy Analysis,RDA)。分析前,先对数据进行Log转换,以减少数据间差异。以全部环境因子为解释变量,以土壤酶活性为响应变量,依据解释变量前向选择,并通过蒙特卡洛置换检验(Monte Carlo Test)得出每个因子的条件效应。
2 结果与分析
2.1 降水量对土壤酶活性的影响
双因素方差分析结果表明(表2),降水量对过氧化氢酶活性有显著影响(P<0.05),对脲酶和蔗糖酶活性有极显著影响(P<0.01);分解时间对过氧化氢酶、蔗糖酶和脲酶活性均有极显著影响(P<0.01);降水量与分解时间的交互作用对3种酶活性均无显著影响(P>0.05)。

表2 降水量、分解时间及其交互作用对土壤酶活性的影响Table 2 Effect of precipitation, decomposition time and their interaction on soil enzyme activity
凋落物分解过程中,过氧化氢酶活性随分解时间呈上升趋势,蔗糖酶和脲酶活性均在240 d时酶活性最低,但无明显规律(图2)。相同分解时间下,与自然降水量相比,减少(W1和W2)和增加(W4和 W5)降水量整体上对过氧化氢酶活性影响较小(180 d除外);减少(W1和W2)和增加30%降水量对蔗糖酶活性无显著影响(P>0.05),增加50%降水量在180 d时显著提高了蔗糖酶活性(P<0.05);减少30%和50%降水量对脲酶活性影响较小(180 d除外),增加降水量在180 d(W4和W5)、240 d(W4)和360 d(W4和W5)时显著提高了脲酶活性(P<0.05)。
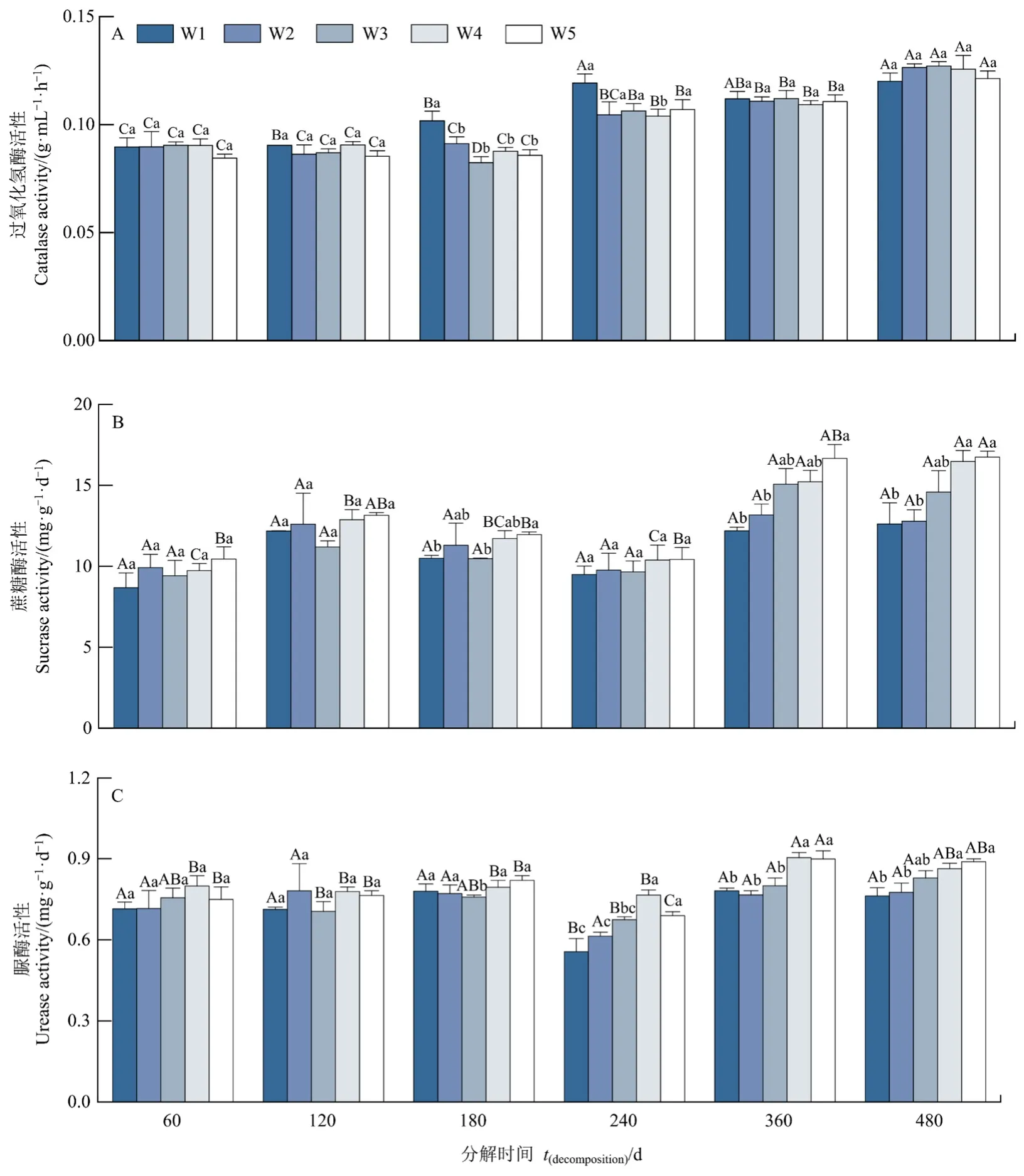
图2 降水量对土壤酶活性的影响Figure 2 Effects of precipitation on soil enzyme activities
此外,相同降水量处理下,酶活性在分解时间间存在差异,尤其过氧化氢酶活性(图2):减少50%降水量处理下,过氧化氢酶活性在240 d和480 d时显著高于除360 d外的其他分解时间(P<0.05)。蔗糖酶活性在分解时间间无显著差异(P>0.05)。脲酶活性在240 d时显著低于其他分解时间(P<0.05);减少30%降水量处理下,过氧化氢酶活性在480 d时显著高于其他分解时间(P<0.05)。蔗糖酶和脲酶活性在分解时间间无显著差异(P>0.05);自然降水量处理下,过氧化氢酶在480 d时显著高于其他分解时间(P<0.05)。蔗糖酶活性在分解时间间无显著性差异(P>0.05)。脲酶活性在120 d和240 d时显著低于360 d和480 d(P<0.05);增加30%降水量处理下,过氧化氢酶活性在240、360和480 d时显著高于其他分解时间(P<0.05)。蔗糖酶活性在360 d和480 d时显著高于其他分解时间(P<0.05)。脲酶活性在360 d时显著高于除480 d外的其他分解时间(P<0.05);增加50%降水量处理下,过氧化氢酶活性在240、360、480 d时显著高于其他分解时间(P<0.05)。蔗糖酶活性在480 d时显著高于60、180和240 d(P<0.05)。脲酶活性在360 d时显著高于除480 d外的其他分解时间(P<0.05)。
对6个分解时间的酶活性进行了整理汇总(图3)。与自然降水量相比,减少(W1和W2)和增加(W4和W5)降水量对过氧化氢酶和蔗糖酶活性无显著影响(P>0.05);减少50%降水量显著降低了脲酶活性(P<0.05),减少30%和增加(W4和W5)降水量对脲酶活性无显著影响(P>0.05)。
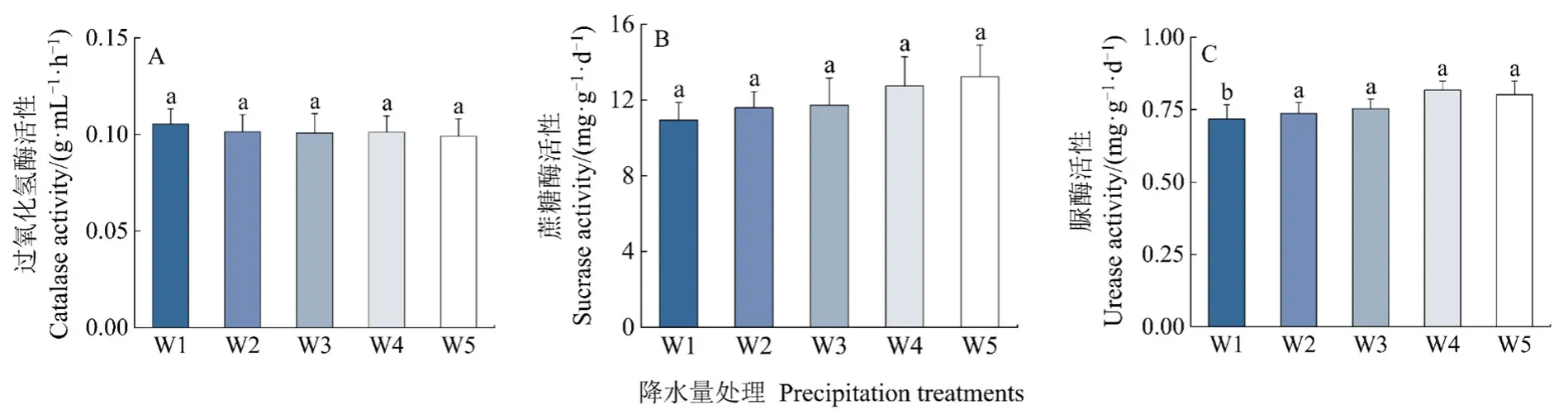
图3 降水量对土壤酶活性的影响Figure 3 Effect of precipitation on soil enzyme activity
2.2 降水量对土壤理化性质的影响
凋落物分解过程中,土壤含水量、电导率、NH4+-N和NO3--N无明显的变化规律,pH变化幅度较小,速效P呈上升趋势(图4)。与自然降水量相比,减少降水量在180 d(W1和W2)和240 d(W1)时显著降低了土壤含水量(P<0.05),增加30%和50%降水量在60、120和240 d时显著提高了含水量(P<0.05);多数情况下,减少和增加降水量对土壤 pH、电导率和 NH4+-N无显著影响(P>0.05);减少降水量在120 d(W1和W2)、180 d(W1)、240 d(W1 和 W2)、480 d(W1)显著提高了NO3--N(P<0.05),增加降水量在120 d(W4和 W5)、240 d(W4和 W5)、480 d(W5)显著提高了 NO3--N(P<0.05);减少降水量在 120 d(W1)、180 d(W1和W2)显著提高了速效P(P<0.05),增加降水量在 60 d(W5)、120 d(W5)、240 d(W4和 W5)、480 d(W4和 W5)显著降低了速效 P(P<0.05)。
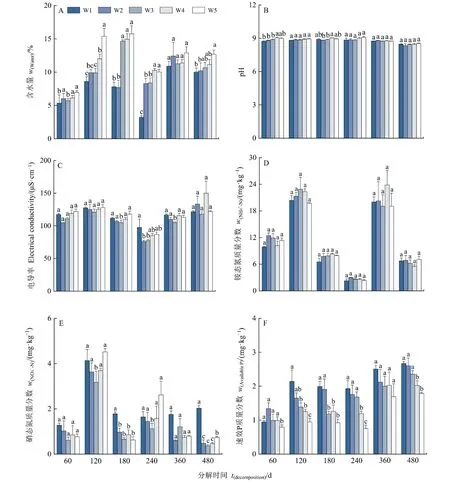
图4 降水量对土壤理化性质的影响Figure 4 Effects of precipitation on soil physicochemical properties
2.3 土壤酶活性与环境因子间的关系
RDA结果显示(图5),2个典范轴分别解释了69.56%和1.82%的土壤酶活性变异。对酶活性影响显著的环境因子依次为月平均风速、月平均气温、月降水量、土壤含水量、土壤 N∶P和土壤 NH4+-N(P<0.05,表3)。其中,月平均风速和土壤 N∶P的贡献率较高,月降水量和土壤NH4+-N的贡献率较低。3种酶活性均与月平均风速、土壤N∶P、土壤含水量和土壤NH4+-N呈较强的正相关关系(P<0.05),与月降水量呈较强的负相关关系(P<0.05);蔗糖酶和脲酶活性还与月平均气温呈较强的负相关关系(P<0.05)。
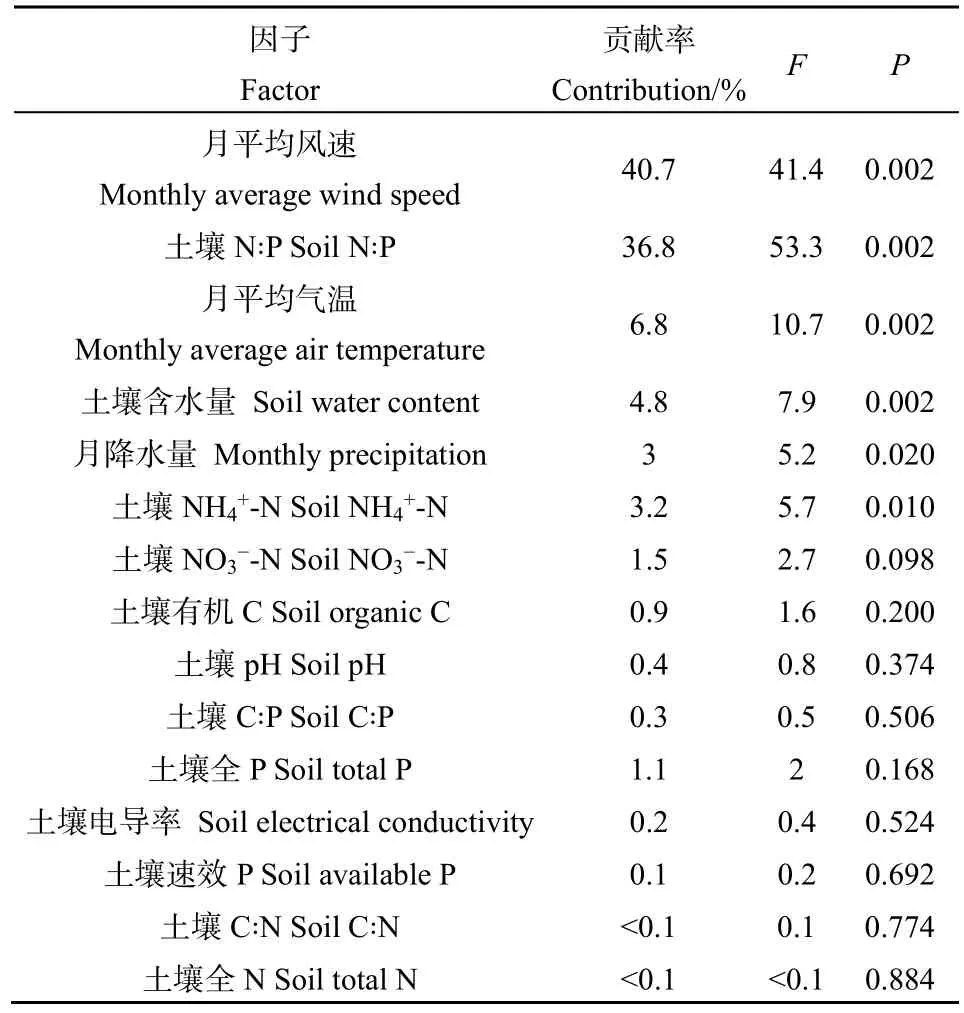
表3 土壤酶活性与环境因子RDA统计学分析Table 3 Statistics analysis in RDA of soil enzyme activities and environmental factors
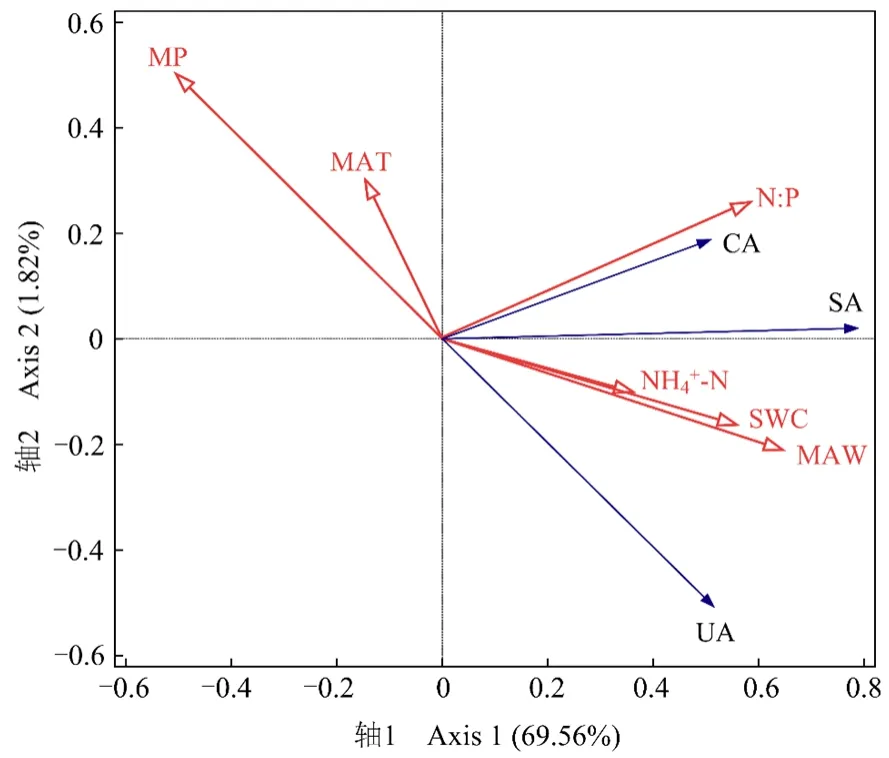
图5 土壤酶活性与环境因子的RDAFigure 5 RDA of soil enzyme activities and environmental factors
3种酶活性均与草木樨状黄芪、牛枝子、猪毛蒿和白草凋落物C累积释放量呈极显著正的线性关系(P<0.01,图6),即3种酶活性随植物种凋落物C累积释放量的增加而增加。过氧化氢酶和蔗糖酶活性与草木樨状黄芪凋落物 N累积释放量呈显著正的线性关系(P<0.05,图6B),脲酶活性与草木樨状黄芪无显著的线性关系(P>0.05)。3种酶活性均与牛枝子、猪毛蒿和白草凋落物N累积释放量呈极显著正的线性关系(P<0.01),即3种酶活性随牛枝子、猪毛蒿和白草凋落物C累积释放量的增加而增加。
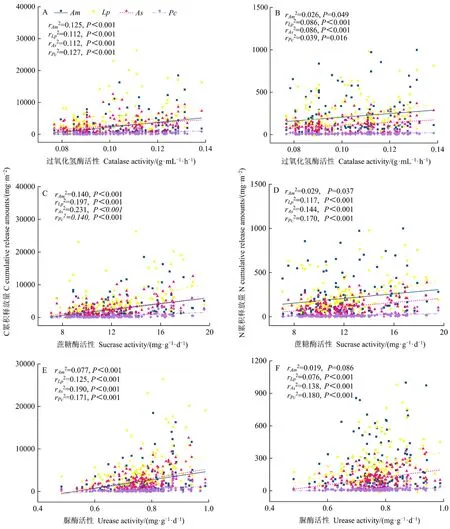
图6 土壤酶活性与植物种凋落物元素累积释放量的线性拟合关系Figure 6 Linear fitting relationships between soil enzyme activities and elemental cumulative release amounts from plant species litters
3 讨论
3.1 土壤过氧化氢酶活性随分解时间增加,蔗糖酶和脲酶活性无明显的时间动态
土壤酶活性的时间动态受土壤温度和含水量的调控(王理德等,2016)。本研究中,凋落物分解过程中各降水量处理下过氧化氢酶活性随分解时间呈上升趋势(图2)。这可能是因为,随着凋落物分解时间的推移(前360 d),土壤温度逐渐升高、新鲜凋落物输入增多,酶活性相应提高(曹聪等,2020);另外,过氧化氢酶参与土壤物质和能量的转化,其活性与凋落物中木质素降解和矿化有很大联系(Nannipieri et al.,2012)。凋落物分解后期纤维素、木质素等难分解物质占比增大(Zechmeister-Boltenstern et al.,2015),因此过氧化氢酶活性在凋落物分解后期(360 d和480 d)也呈上升趋势。蔗糖酶和脲酶活性缺乏明显的时间动态变化规律,但均在分解240 d时最低。此时为生长季旺盛期(2017年8月16日),较高的空气温度下(图1),过低的土壤含水量(图 4)和过高的土壤温度可能限制了微生物和植物根系活动(Nannipieri et al.,2012),从而抑制了蔗糖酶和脲酶的分泌。分解后期随着土壤含水量升高以及温度降低,两种酶活性也呈升高趋势(图2)。
3.2 减少降水量对土壤酶活性影响较小,增加降水量有助于提高蔗糖酶和脲酶活性
降水调控着植物地下部分生长和微生物活性,影响着土壤酶活性(Shi et al.,2014;Esch et al.,2017)。干旱条件下,植物地下部分生长慢、微生物活性低,不利于土壤酶的分泌。本研究中,与自然降水量相比,凋落物分解过程中减少降水量对3种酶活性影响较小。这可能是由于研究区植物和微生物长期生长于少雨的环境下,对干旱具有高的适应性,二者生长繁殖和酶的分泌受短期降水量减少的影响较小,与其他针对干旱荒漠区的研究结果一致(许华等,2018)。然而,综合6个分解时期的结果表明,降水量减少50%降低了脲酶活性(图3),证实极端干旱限制了酶活性(Gao et al.,2021);增加降水量不仅提高了土壤含水量,还会加速凋落物淋溶和物理破碎,促进有机C及养分释放归还给土壤(Allison et al.,2013),增加了微生物活性及其底物可获得性,促进了微生物向土壤中释放酶。增加降水量在凋落物分解中期提高了蔗糖酶和脲酶活性(图2)。与降水量增加30%相比,降水量增加50%对酶活性的影响更大,可能是因为影响酶活性的土壤水分含量存在阈值,在阈值范围内酶活性随土壤含水量的增加而升高(Kivlin et al.,2014)。
3.3 土壤酶活性主要受气象因子影响
自然环境下,土壤酶活性受气候、植物和土壤等因子综合作用的影响(Burns et al.,2013)。本研究中,3种酶活性与月平均风速正相关(图5和表3),原因可能是在一定的风速范围内较高的风速有利于凋落物的物理破碎,增加凋落物与分解者的接触面积,进而加快微生物在凋落物酶和土壤酶系统作用下的酶解过程(贾丙瑞,2019)。本研究发现,3种酶活性与月降水量负相关(P<0.05),且蔗糖酶和脲酶活性与月平均气温负相关(P<0.05)。可能是试验地位于干旱半干旱地区,该区蒸发量大、降水量高的月份平均气温也高(图1),水分受温度影响未及时渗入土壤而挥发,影响土壤养分溶解和扩散,限制了土壤酶活性(Jing et al.,2014)。本研究中3种酶活性均与土壤含水量正相关(P<0.05),表明土壤含水量增加会为各种酶促反应提供反应条件和场所,对酶活性升高起促进作用,与前人研究结果较为一致(Ladwig et al.,2015;Gao et al.,2021)。此外,3种酶活性与土壤 N∶P和 NH4+-N正相关(P<0.05),表明三者与土壤 N、P之间存在协同恢复关系,即当土壤全N增加时土壤酶积极参与二者的转化分解过程(王涛等,2018)。
3.4 土壤酶活性与植物种凋落物元素累积释放量密切相关
作为植物与土壤之间物质交换的枢纽,凋落物通过分解将物质和营养元素返还给土壤,其细胞中的酶也会释放进入土壤,驱动和调节着土壤酶活性(陈晓丽等,2015)。本研究中,3种酶活性与植物种凋落物元素累积释放量存在不同程度的线性关系,可能是因为凋落物分解会释放出大量可溶性和不溶性碳水化合物,这些碳水化合物通过刺激微生物多样性和生物量影响土壤酶活性(Kourtev et al.,2002;Kotroczo et al.,2014)。然而,不同植物凋落物会导致土壤微生物量、区系组成和代谢过程改变,其元素累积释放量对酶活性响应也会有所差异(Kourtev et al.,2002);草木樨状黄芪N累积释放量与脲酶活性无显著的线性关系(P>0.05),但其余植物种凋落物 N累积释放量均与 3种酶活性呈显著正的线性关系。进一步证实,虽然凋落物N是土壤-凋落物界面酶活性的重要影响因素(Ge et al.,2017),但不同植物种凋落物在养分利用效率(Waring,2013)和养分释放率(韩翠等,2022)等方面存在差异,使得土壤酶活性与凋落物N累积释放量之间的关系在不同植物种凋落物类型间表现不同。
4 结论
综上,(1)凋落物分解过程中,土壤过氧化氢酶活性呈上升趋势,蔗糖酶和脲酶活性缺乏明显的时间动态。(2)减少降水量对3种土壤酶活性影响较小。增加降水量有助于提高蔗糖酶和脲酶活性,但其影响程度在凋落物分解时期间存在差异。(3)凋落物分解过程中土壤酶活性较多地受气象因子调控,而非土壤因子。(4)随着降水量增加,土壤水分和N有效性提高,从而刺激了酶活性。随着土壤酶活性增加,研究区优势和常见植物种凋落物分解加快、元素释放增多,从而加快了植被—土壤系统生物地球化学循环。本文未同步监测土壤温度。土壤温度是调控土壤酶活性的主要因子之一,因此今后还需结合土壤温度等数据,通过长期的定位观测,深入探讨降水量变化下荒漠草原土壤酶活性的驱动因素。
