侧柏人工林林分空间结构对林下草本多样性的差异性影响及其关联度
2022-11-04张林周飘齐实张岱伍冰晨崔冉冉
张林,周飘,齐实,张岱,伍冰晨,崔冉冉
北京林业大学水土保持学院/水土保持国家林业局重点实验室,北京 100083
林下草本植物是森林群落的重要组成部分,在维护森林生态系统多功能的正常发挥中扮演着至关重要的角色作用(Kuiters,2013;Ahmad et al.,2018;郭彩云等,2021;王娜等,2021)。然而,林下草本植物的组成和分布受林分结构(胡文杰等,2019)、气候(张蕊等,2019)、土壤(闫玮明等,2019)和立地条件(张晓龙等,2020)等多因素综合影响。其中,乔木层的林分结构在局部小尺度范围内被认为是制约林下草本植物多样性的主要因素(Cook,2015),这是因为上层乔木的林分结构直接决定着林下水、热、光照和土壤养分等基本条件,进而使得不同林分结构下的草本物种多样性产生差异(Sabatini et al.,2014;Capellesso et al.,2016)。但影响林下草本物种多样性的主导林分结构因子尚不清楚,应值得进一步研究。因此明确影响林下草本物种多样性的主导林分结构因子对森林的可持续经营具有重要的理论和实际意义。
近年来,多数的研究倾向于分析海拔(刘秉儒,2021)、坡向(姜倪皓等,2021)、土壤(杨胜香等,2021)等环境因子对林下草本植物多样性的影响。例如,邓清月等(2019)指出,海拔是影响林下草本物种多样性的主要因子;姜倪皓等(2021)研究了林下草本多样性与环境因子的关系,认为坡向是限制林下草本物种多样性的主导因子;闫玮明等(2019)认为林下草本植物多样性主要受土壤有机质和氮磷含量的影响。事实上,林下草本物种的分布格局不仅归因于海拔、土壤等环境因子,还受到上层林分结构的显著影响(Chen et al.,2014),但专门的理论研究少有报道;王媚臻等(2019)研究了柏木(Cupressus funebris)林下草本植物多样性对林分密度的响应,认为低密度造林对林下草本植物多样性具有促进作用;张洋洋等(2021)研究表明,林龄通过改变土壤养分等基本条件来调控林下草本植物的生长和发育;陈丝露等(2018)以不同改造模式的柏木林为研究对象,认为混交林改造模式对林下草本物种多样性具有抑制作用。然而,以上研究只是单一阐述一个或两个林分结构因子对林下草本物种多样性的影响,而针对多林分结构因子对林下草本物种多样性的相对贡献研究鲜有报道,这难以充分揭示林分结构影响林下草本多样性的真实机理,并且关于如何提升林下草本植物多样性,一直没有提出相应的优化策略。因此,明确影响林下草本物种多样性的主导林分结构并提出相应的优化配置策略已成为目前生态学研究领域的核心问题。
侧柏(Platycladus orientalis)作为北京市京津风沙源二期林业工程的主要造林树种,其工程目的在于改善林分结构,提高林下植物多样性。因此,如何定量分析影响侧柏林下物种多样性的主导林分结构因子,并采取有效措施提高其林下物种多样性是北京市京津风沙源低效林改造工程亟待解决的问题。基于此,本研究以北京山区侧柏人工林为研究对象,基于27个标准样地的实测数据,采用多元逐步回归法筛选出了影响侧柏林下草本植物多样性的主导林分结构因子,并用响应面分析法将主导林分结构因子进行优化,进而提出相对应的林分结构优化调整策略,以期为北京山区林地森林质量的提升提供理论和技术支撑。
1 材料与方法
1.1 研究区概况
研究区为北京市西部山区京津风沙源治理二期林业工程区,包括房山、门头沟区、昌平,地理位置是:115°23′—115°44′E,39°15′—40°55′N。气候类型以暖温带半湿润季风大陆性气候为主,年均降水量为490—630 mm,年均气温10—13 ℃。山区地貌类型复杂多样性,平均海拔200—450 m,土壤类型有山地褐土和沙壤土,土层薄。工程造林主要以侧柏(Platycladus orientalis)为优势树种,黄栌(Cotinus coggygria)、山杏(Armeniaca sibirica)、元宝枫(Acer truncatum)等也有少量分布,林下草本植物则以白莲蒿(Artemisia stechmanniana)、丛生隐子草(Cleistogenes caespitosa)、小红菊(Chrysanthemum chanetii)、细叶薹草(Carex rigescens)、委陵菜(Potentilla chinensis)、中华卷柏(Selaginella sinensis)等为主。
1.2 样地选取与样品采集
2021年7月,根据森林二类资料调查,依据侧柏林地分布的代表性和均匀性,在研究区内选择立地条件和土壤因子基本一致,人为干扰较少的以侧柏为优势树种的林分为研究对象,共设置27块20 m×20 m的标准侧柏林样地,测量样地内乔木树种的树高、胸径、冠幅、林分密度等基本林分结构参数,并记录标准样地的坡向、海拔、土层厚度等立地因子。同时在每个标准样地的对角线上中下随机布设5个1 m×1 m的草本样方,并记录各样方内草本植物的名称以及每个物种的株数,用于计算其林下草本植物的多样性。统计整理后标准样地的林分结构和立地因子见表1。
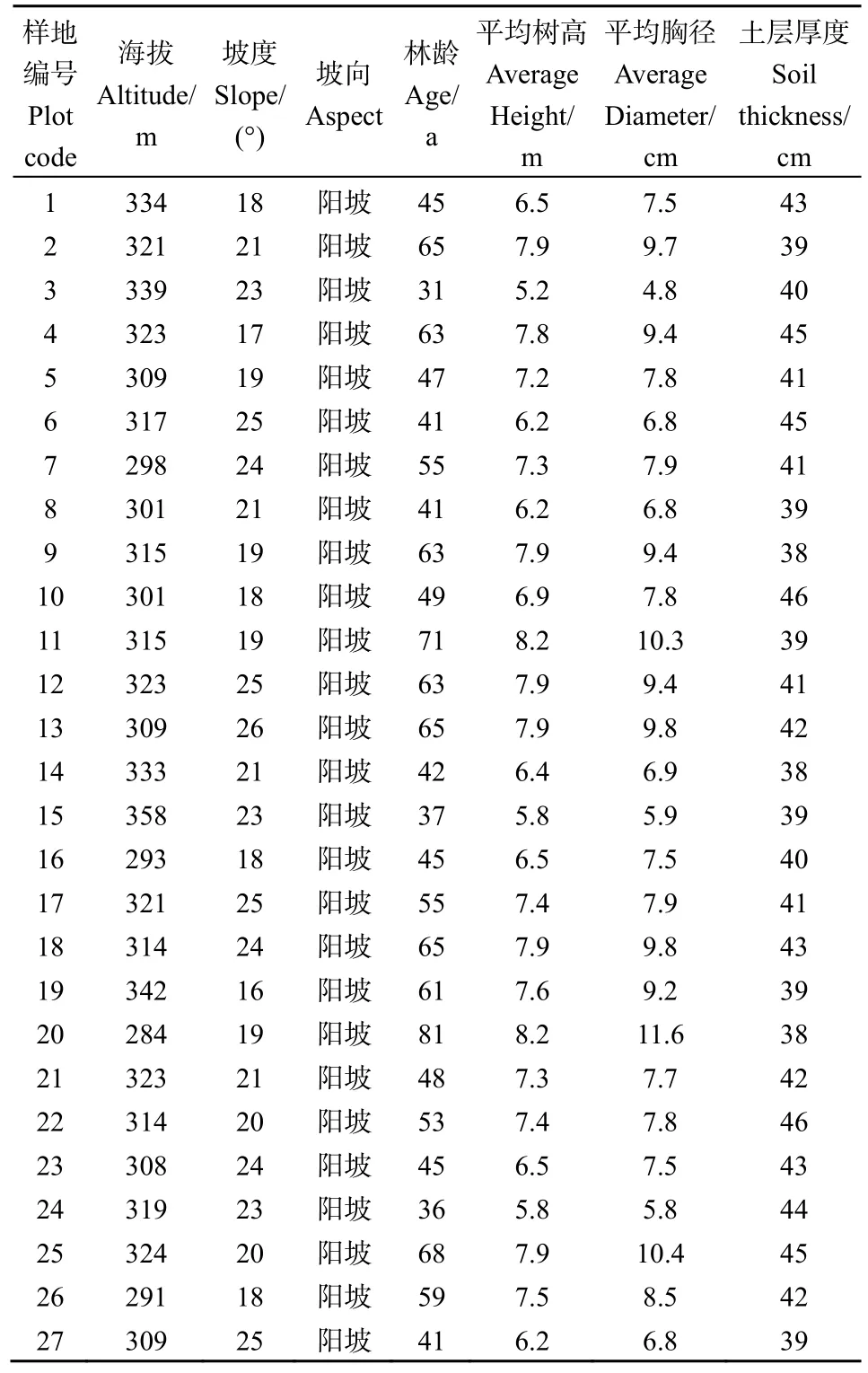
表1 样地基本特征Table1 Basic characteristics of the sample plot
1.3 研究方法
1.3.1 林分结构基本参数确定
本研究选取林分密度、郁闭度、林龄、平均树高和平均胸径等表示林分非空间结构,选取角尺度、混交度和大小比等表示林分空间结构。其中,角尺度、混交度、大小比根据每木检尺的相对坐标计算,具体计算参考惠刚盈等(2001)计算空间结构指数方法,林分密度通过计算单位面积的林木数量获得,乔木层郁闭度采用样线法调查,按对角线总长与树冠冠幅总长之比计算郁闭度大小。
1.3.2 多样性指数计算
本研究选用Simpson多样性指数、Pielou均匀度指数、Margalef丰富度指数来表示林下草本物种多样性水平,计算公式如下:
Simpson多样性指数:

Pielou均匀度指数:

Margalef丰富度指数:

式中:
S——林下草本种类总数;
pi——第i种个体数占所有种个体总数的比列;
N——林下草本个体总数。
1.3.3 林下草本物种多样性综合指数计算
本研究采用均方差决策分析法计算各样地林下草本物种多样性综合指数。由于物种多样性指标多种多样,且量纲不同,因此,首先对各样地多样性指标进行标准化处理,并计算每个指标的均方差,从而得到每项多样性指标的权重系数,最后将不同量纲多样性指标统一计算并进行比较,以最终计算出的总分值大小来衡量不同样地类型下的草本物种多样性整体水平,其计算公式为:
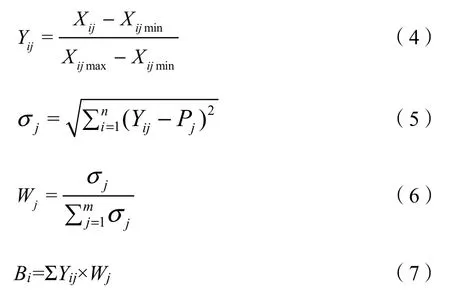
式中:
Bi——多样性综合值;
Wj——第j项多样性指标的权重;
σ——第j项指标的均方差。
1.3.4 多元回归分析
由于林下草本物种多样性受多林分结构因子综合影响,因此,可将各多样性指数作为因变量,将林分密度、郁闭度、混交度、角尺度等林分结构因子作为自变量进行多元线性逐步回归分析,筛选出影响林下草本物种多样性的主导林分结构因子。
1.3.5 响应面分析
响应面分析法是一种快速分析和构建多指标与响应值之间的模型方程的分析法,该模型方程可通过给定响应值的目标条件进而对所有参评指标进行优化,确定评价指标与之相对应的优化组合值。本文选用中心复合试验设计,通过构建多元二次回归方程模型对影响侧柏人工林林下草本植物多样性的主导林分因子进行优化(即优化目标),方程模型如(8)示:
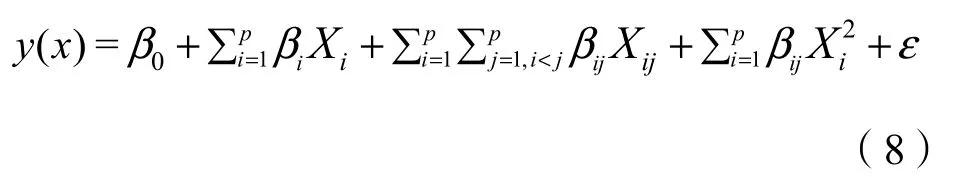
式中:
Xi——自变量;
y(x)——响应值;
β0——常数项;
βi——线性影响;
βij——交互影响;
ε——误差项。
1.4 数据处理与分析
本研究使用Excel 2018和SPSS Statistics 20进行数据整理、统计、分析和绘图,利用Pearson相关系数法将各林分结构指标与林下草本物种多样性指标进行相关性分析,采用Minitab 19软件中的响应面方法(RSM)构建主导林分结构因子与林下草本多样性综合指数的三维曲面方程和等值线图。
2 结果与分析
2.1 林分结构及多样性基本特征
由表2可知,研究区内侧柏林的林分密度均值为 1182.51,变异系数为 30.35%;郁闭度均值为0.71,变异系数为16.72%;角尺度均值为0.46,变异系数为27.07%,树木接近随机分布状态;混交度均值为0.29,变异系数为55.78%,隔离程度较好;大小比均值为0.49,变异系数为9.17%;树高和胸径的均值分别为 7.07和 8.12,变异系数分别为12.09%和19.29%。
研究区侧柏林下草本植物较为丰富,共有植物82种,分属34科75属。其中,侧柏林下草本植物以细叶薹草(Carex rigescens)、中华卷柏(Selaginellasinensis)、白莲蒿(Artemisia stechmanniana)、芨芨草(Achnatherum splendens)、黄背草(Themeda triandra)、马塘(Digitaria sanguinalis)、丛生隐子草(Cleistogenes caespitosa)为优势种,其重要值均值分别为17.61%、10.28%、9.56%、8.66%、8.60%、7.45%、5.73%。由表2可知,侧柏林下草本Simpson多样性指数、Pielou均匀度指数、Margalef丰富度指数和多样性综合值均值分别为0.59、0.78、1.25、0.57,其中Simpson多样性指数变异系数为21.07%,Pielou均匀度指数变异系数为19.05%,两者变异系数较小;而Margalef丰富度指数和多样性综合指数的变异系数较大,其值分别为43.63%、39.79%。上述研究结果表明各样地林下物种多样性水平相差较大,这也充分说明了各样地上层乔木的林分结构存在差异。
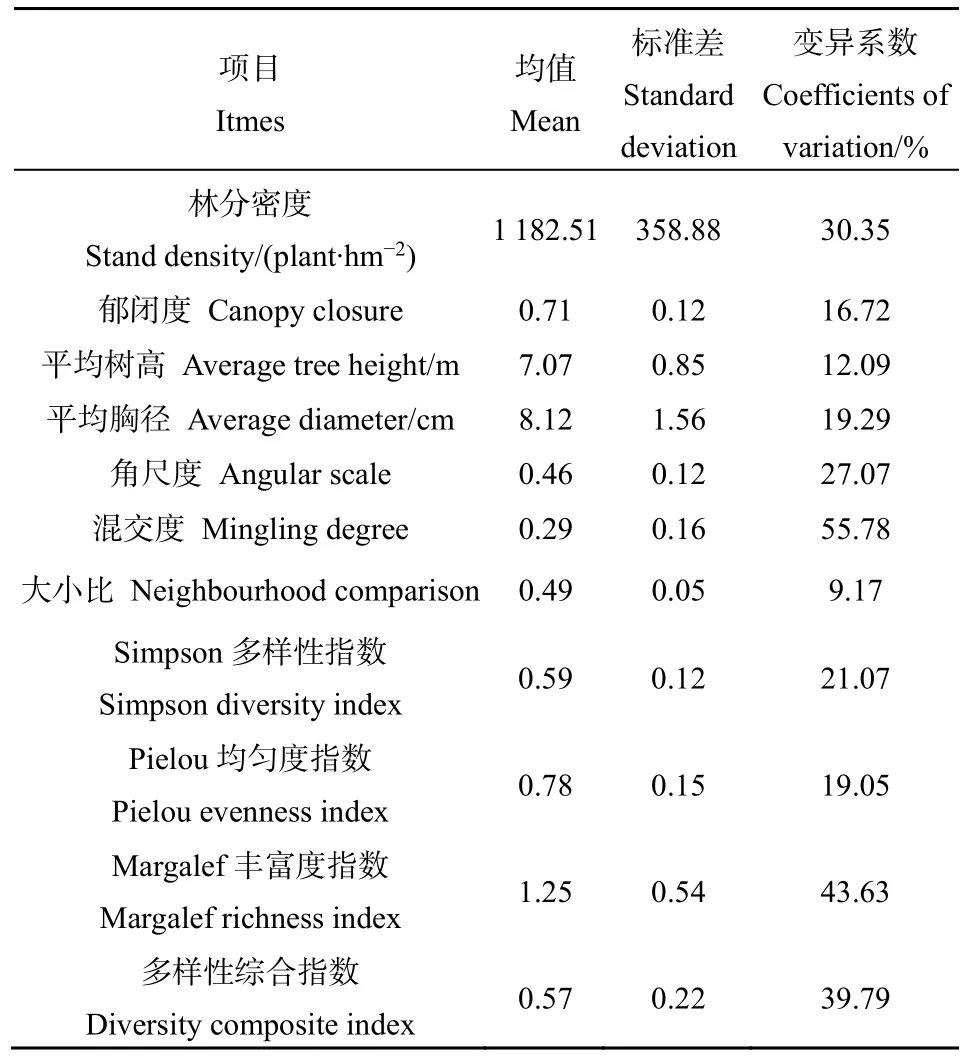
表2 林分结构及多样性指数Table 2 Stand structure and diversity index
2.2 林分结构因子与林下草本植物多样性的相关性分析
由表3可知,Simpson多样性指数与林分密度和郁闭度极显著相关(P<0.01),与混交度显著相关(P<0.05);Pielou均匀度指数与郁闭度和混交度极显著相关(P<0.01),与林分密度显著相关(P<0.05);Margalef丰富度指数与林分密度显著相关(P<0.05),与郁闭度和混交度无显著相关关系(P>0.05);而林龄、平均胸径、平均树高、角尺度和大小比与各指数均不相关(P>0.05)。

表3 林分结构因子与多样性指数的相关性分析Table 3 Correlation analysis between stand structure factors and diversity index
2.3 林下草本物种多样性与林分结构因子的多元逐步回归分析
将各多样性指标与林分结构因子进行多元逐步回归分析,由表4可知,最优回归方程系数拟合度良好,R2分别为0.401,0.406,0.141,0.411,说明该模型可用于检验影响林下草本物种多样性的主导林分结构因子,在8个林分结构因子中,影响林下 Simpson多样性指数的主导林分结构为郁闭度;影响Pielou均匀性指数的主导林分结构为郁闭度和混交度;影响Margalef丰富度指数的主导林分结构为林分密度;而对林下草本物种多样性整体水平起决定作用的林分结构为郁闭度和混交度。因此,预维持和提高林下草本植物多样性整体水平,可适当调整林分郁闭度和混交度。
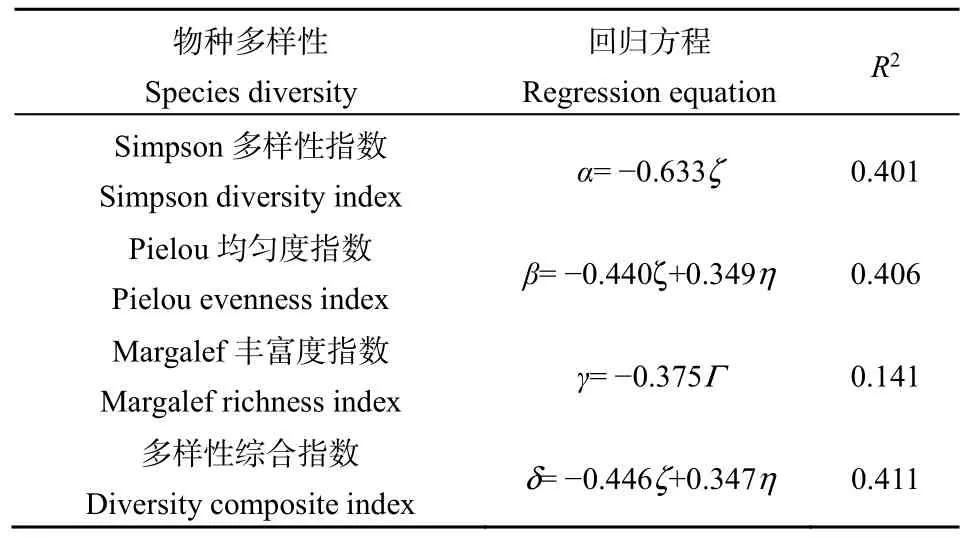
表4 林分结构因子与林下草本物种多样性的逐步回归方程Table 4 Stepwise regression equation between stand structure factors and understory herbaceous species diversity
2.4 以提升林下草本物种多样性为导向的主导林分结构优化配置
上述研究表明,林分郁闭度和混交度是影响林下草本物种多样性整体水平的主导林分结构因子,因此对侧柏林的郁闭度和混交度进行响应面分析,经过二次回归拟合分析可得等值线(图1),对应的回归方程公式(表5)所示,方程中δ表示多样性综合指数,ζ代表郁闭度,η代表混交度,响应面方程的相关系数和调整系数分别为0.82和0.76,说明分析得到的效应曲面方程是可接受的,可用于表达研究区侧柏林分结构优化理论分析。根据已有的实测数据进行以提升林下草本物种多样性整体水平为导向型的林分结构优化分析,结果表明响应面方程的预测相关系数为0.51,说明该响应面方程可用于预测侧柏林分结构的优化配置研究。研究样地林下草本物种多样性的综合值最大为0.92,因此,以多样性整体最大化(0.92)为目标导向,以侧柏林的郁闭度和混交度指标分布范围作为参照,将林分结构进行优化研究,优化结果表明林分结构的最优配置为林分郁闭度=0.58,混交度=0.60。根据预测,林下草本物种多样性综合指数与优化前的侧柏林下多样性综合指数平均水平(0.57)相比,预期值将提高约41.19%。
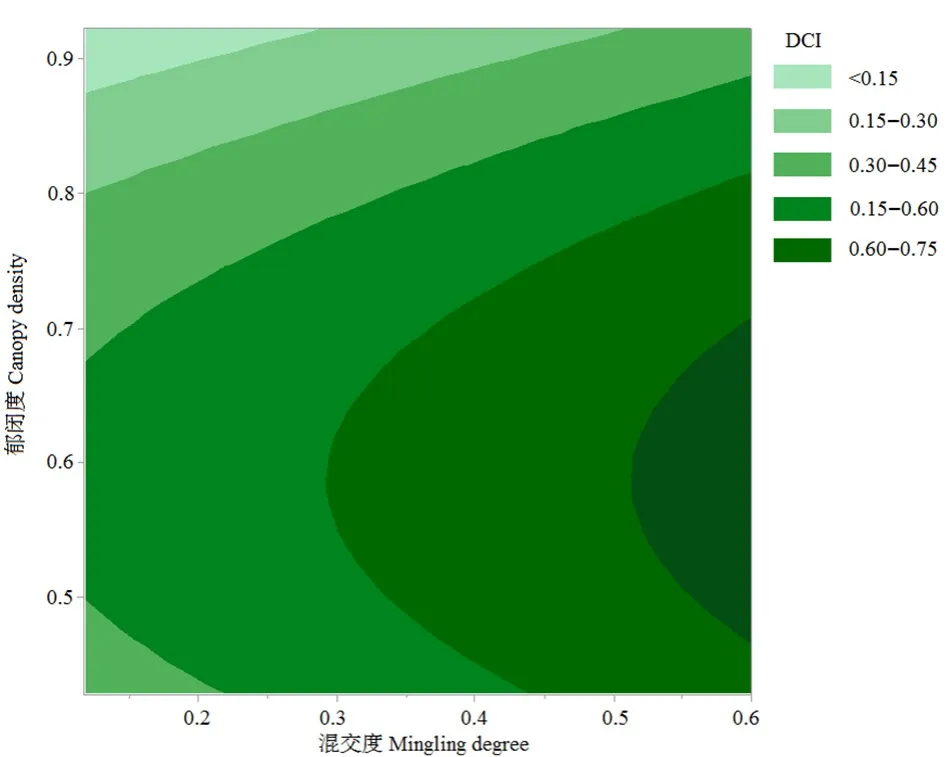
图1 郁闭度和混交度对物种多样性的等值线图Figure 1 Contour map of canopy density and mingling density on species diversity

表5 多样性综合指数与郁闭度、混交度的非线性回归方程Table 5 Nonlinear regression equation of comprehensive diversity index, canopy density and mingling degree
3 讨论
林下草本植物多样性与上层林分结构的关系始终是生态学研究的核心和热点问题之一,明确影响林下草本物种多样性的主导林分结构对森林的可持续经营具有重要意义(Crous-Duran et al.,2019;Lin et al.,2021)。本研究选取了林分密度、郁闭度、平均树高、平均胸径和林龄表征为林分非空间结构因子,选取了混交度、角尺度、大小比表征为林分空间结构因子,采用Pearson相关性分析和多元回归分析探讨了各林分结构因子对林下草本物种多样性的相对重要性。结果表明,林分郁闭度、林分密度、混交度对林下草本物种多样性影响程度相对较高,而大小比、角尺度和平均树高等其他林分结构因子对林下草本物种多样性无显著影响(P>0.05)。
本研究认为,乔木层的郁闭度是制约林下草本物种多样性的主导林分结构因子(P<0.01),这与Wagner et al.(2011)研究认为郁闭度是决定林下草本植物组成和分布的最主要因素研究结论基本一致,这可能是因为乔木层的郁闭度直接决定林内的有效光强度,对林内光、热、水等资源具有再分配作用,从而影响了林下草本植物的分布和生长,特别是对一些喜阳草本植物,随着光照强度的变化,其种群发育会发生改变,进而改变了林下草本的物种多样性(Chávez et al.,2012)。另一方面,在低郁闭度的林内,光照强度的增加促进了林下枯落物的分解和土壤微生物活动,加速了土壤有机质的形成,进而有利于草本物种吸收更多的土壤养分,间接的改变林下草本的组成和分布(Chastain et al.,2006;Wagner et al.,2011)。
研究发现,林分密度对林下草本物种多样性具有显著影响(P<0.05),且林分密度与林下草本物种多样性呈负相关关系,这与王玲(2020)研究认为林分密度是决定林下物种多样性的最主要因素,且随着林分密度的增加,林下草本物种多样性逐渐降低结论基本一致,这可能是因为林分密度直接决定着林内光、热、温度、湿度、土壤养分等一系列草本植物生长的环境(董威等,2019),高密度的侧柏优势树种抑制了林下草本对光的吸收效率,使得林下喜阳草本植物逐渐消失,且高密度的优势树种充分吸收了土壤养分,不利于处于劣势的草本植物生长(Machado et al.2019;张柳桦等,2019)。但有些学者(张勇强等,2020)认为,林下草本植物多样性与林分密度呈二次函数关系,与本研究的结论产生一定的差异,这可能是研究样地的气候类型、土壤类型、林分结构、立地因子等因素存在差异。因此,今后的研究可注重解析林分结构和立地条件对林下物种多样性的耦合影响机制。
研究同时发现,林分混交度也是影响林下草本物种多样性的主导林分结构因子,且混交度与林下草本多样性呈正相关,这与曹小玉等(2019)认为混交度是影响林下草本植物多样性最主要的林分结构因子,且随着混交度的增加,林下草本物种多样性逐渐增加研究结果基本一致,这可能是因为林分混交度的高低决定着林内乔木树种的竞争程度,混交度越高,加剧了乔木树种之间的相互竞争,从而为林下草本植物的生长释放了营养空间(Zhang et al.,2011;He et al.,2013;孙宇等,2020),且多种混交树种产生的枯枝落叶为土壤提供了丰富的有机质来源(Wang et al,2020;曹小玉等,2020;祁雪连等,2021),从而更有利于林下草本植物的生长。但许多学者(崔宁洁等,2014;张沛健等,2021)认为,林龄也是影响林下草本物种多样性的主导林分结构,与本研究产生一定的差异,这可能是本研究的侧柏林基本接近中成熟林,对于幼林龄侧柏林下草本植物多样性尚未研究,因此,在今后的研究中,可注重研究整个侧柏生长周期的林下物种多样性变化,从而为森林的可持续经营提供更多的科学理论依据。
合理的林分结构是提高森林生态系统服务功能的关键,而北京山区地形地貌条件较为恶劣,如何提高侧柏林下草本植物多样性已成为北京市京津风沙源低效林改造工程亟待解决的问题。本研究发现影响林下草本物种多样性整体水平的主导林分结构因子为林分郁闭度和混交度。因此,为提高林下草本物种多样性,可将郁闭度调整为0.58,林下物种多样性较为稳定,适当补植阔叶树种将混交度调整为0.60,形成结构复杂的复层针阔混交林。本研究在探讨林分结构与林下物种多样性的关系时,未充分考虑土壤、海拔等立地因子是否会对侧柏林下草本植物多样性产生影响,因此,在今后的研究中,应考虑将林分结构和环境因子相结合,且增加样本的数量,全面系统地解析影响侧柏林下草本植物多样性的维持机制。
4 结论
郁闭度和混交度是影响北京山区侧柏林下草本植物多样性整体水平的主导林分结构。影响多样性指数的主导林分结构因各指数差异而异,其中,影响林下草本Simpson多样性的主导林分结构因子为郁闭度,影响林下草本Pielou均匀度指数的主导林分结构因子为郁闭度和混交度,影响林下草本Margalef丰富度指数的主导林分结构因子林分密度。因此,欲维持和提高林下草本物种多样性整体水平,可适当将郁闭度和混交度调整为0.58和0.60,林下草本植物多样性整体水平预期值约提高41.19%。
