遮阴对紫楠幼苗生长和生理特性的影响
2022-11-03姜宗庆梁俊捷
姜宗庆,梁俊捷,李 豪,曾 理
(江苏农牧科技职业学院,泰州 225300)
光照对植物的更新分布、形态建成和生长发育起着非常重要的影响[1]。如果生长在光照不足的条件下,植物的净光合速率一般都会降低,植物正常的生长发育受到阻碍。相反,夏季少数地区光照强度太高,导致植物出现光抑制乃至光破坏现象,这可能使植物光合机构所接受的光能超过叶片光合作用所能利用的量,最终使光合速率下降,表明光并非越多越好[2]。为了避免受到光抑制或光破坏,植物大多可以通过自身形态和生理的调节来提高光能利用效率,来应对不同的光照条件[3]。前人研究表明,弱光条件下,植株地径增长量较小,而植株为获取充足的光照,向上生长较快,最终株高增长量较大[4]。
紫楠[Phoebe sheareris(Hemsl.)Gamble]为樟科楠属常绿乔木,不仅以材质优良而著称,而且也是优良的园林绿化树种[5-6]。近年来,我国积极倡导发展珍稀乡土树种和珍贵用材树种。目前,关于紫楠的研究主要集中在种子的繁殖特性、育苗技术、天然群落等方面[7-10],而关于遮阴对紫楠幼苗的影响研究鲜见报道。本研究通过不同程度遮阴处理,分析紫楠幼苗的生长指标和生理特性参数对不同处理的响应差异,探讨紫楠幼苗对不同光照处理的适应能力,探求最适宜紫楠幼苗生长发育的光照条件,旨在为紫楠苗木的培育及优化提供理论参考。
1 材料与方法
1.1 试验地概况
试验地设在江苏农牧科技职业学院苗木基地(32°27′N,119°56′E)。该地属亚热带季风气候,温暖湿润,雨水充沛。年平均气温15.3℃,年平均积温5 365.6℃,年平均降雨日113 d,年平均降水量为1 055 mm,年平均日照时数2 205.9 h,无霜期215 d。
1.2 试验材料
选取生长基本一致的当年生紫楠实生苗,初始苗高(6.83±0.08)cm,地径(1.28±0.03)mm。2018年4月20日移栽于规格为24 cm×23 cm×28 cm(上口径×下口径×高)的花盆中,每盆栽植1株。每盆装土为10 kg,为防止水肥流失,每个花盆配有塑料托盘。盆土质地为沙壤土,pH 6.8,有机质1.31 g∕kg,速效钾5.49 mg∕kg,速效磷2.28 mg∕kg,速效氮4.65 mg∕kg。
1.3 试验设计
2018年5月10日将紫楠幼苗移到棚中,采用黑色遮阳网遮阴,设置不同光照强度处理:不遮阴全光照处理(CK)、遮阴率40%处理(T1)、遮阴率60%处理(T2)和遮阴率80%处理(T3)4个处理,采用完全随机区组排列,每个处理重复3次,每个重复选取苗木10盆,株行距20 cm。试验期间进行常规水肥管理,每周随机调整每盆位置,确保各处理光照条件一致。
1.4 测定指标及方法
1.4.1 生长指标测定
试验末期于2018年11月10日,采用直尺测量苗高,地径采用游标卡尺测定。
1.4.2 光合色素含量测定
2018年10月初,选取无病虫害、长势一致的紫楠幼苗中上部的完整功能叶,擦净后剪碎,称取0.2 g,用体积比(丙酮)∶(无水乙醇)∶(水)=4∶5∶1的提取液研磨,过滤并定容至25 mL,分别用分光光度计在波长665 nm、649 nm、470 nm下测定,根据吸光度值,计算类胡萝卜素含量、叶绿素a含量和叶绿素b含量[11]。
1.4.3 光合速率测定
于2018年10月初选择晴天9:00—11:00,使用Li-6400便携式光合测定仪测定。采用自然光源,设定空气流速为0.5 L∕min,CO2浓度为400 μmol∕mol,叶室温度为30℃,叶室相对湿度为50%,测定时选取植株中上部的功能叶,每处理测定5片叶子的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),每个紫楠幼苗叶片重复测定6次,计算平均值。
1.4.4 其他生理指标测定
于2018年10月初采摘成熟功能叶,取部分鲜叶采用电解质外渗量法测定相对电导率。其余叶片液氮冷冻后置于-80℃冰箱中保存待用。参考李合生[12]的方法测定生理指标,各处理重复3次。丙二醛含量测定采用硫代巴比妥酸法;POD活性测定采用愈创木酚法;SOD活性测定采用NBT光化还原法;游离脯氨酸含量测定采用酸性茚三酮法;可溶性蛋白质含量测定采用紫外吸收法;可溶性糖含量测定采用蒽酮比色法。
1.5 数据处理
采用Excel 2013对试验数据进行处理,SPSS 20.0进行LSD多重比较与方差分析。
2 结果与分析
2.1 遮阴对紫楠幼苗生长指标的影响
由表1可知,随着遮阴程度的增加,紫楠幼苗的苗高生长以T1处理为最高,比对照(CK)高6.4%(P<0.05);苗高生长最低的为T3处理,比对照(CK)处理低29.2%。随着遮阴程度的不断增加,紫楠幼苗的地径生长呈下降趋势,T1处理与对照(CK)差异不显著(P>0.05),而中度遮阴的T2处理和重度遮阴的T3处理地径均显著低于CK,降幅分别达13.5%和23.4%。与全光照CK相比,轻度遮阴(T1)紫楠幼苗的地径生长影响不显著,但苗高生长显著增加。

表1 遮阴对紫楠幼苗生长指标的影响Table 1 Effects of shading on growth index of Phoebe sheareris seedlings
2.2 遮阴对紫楠幼苗光合色素含量的影响
由表2可知,随着遮阴程度的增大,叶绿素a、b含量及叶绿素总量呈上升趋势,且各处理差异显著(P<0.05)。T1、T2和T3处理叶绿素a含量分别是CK对照处理的2.22倍、2.79倍和2.97倍,叶绿素b含量分别是CK对照处理的2.23倍、2.96倍和5.58倍,叶绿素总量分别是CK对照处理的2.22倍、2.84倍和3.69倍,说明遮阴下的紫楠幼苗通过增加叶绿素含量,提高对光的吸收能力来适应弱光环境。相反,叶绿素a∕b值随遮阴程度的增大呈下降趋势,T2、T3处理与CK差异达到显著水平,说明叶绿素a含量的增加幅度低于叶绿素b含量。随着遮阴程度的不断增加,类胡萝卜素含量逐渐提高,但其增加幅度小于叶绿素含量,与CK处理相比,T1、T2和T3处理分别高出44.30%、63.61%和67.41%(P<0.05)。

表2 遮阴对紫楠幼苗光合色素含量的影响Table 2 Effects of shading on contents of photosynthetic pigments of Phoebe Sheareris seedlings
2.3 遮阴对紫楠幼苗光合速率的影响
由表3可知,随遮阴程度的增大,T1处理净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)升高,而T2和T3处理净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)则显著下降(P<0.05)。各遮阴处理的Pn依次为CK的101.10%、59.20%和56.46%,Tr依次为CK的102.03%、75.00%和59.30%,Gs依次为CK的106.09%、72.17%和36.52%。胞间CO2浓度(Ci)与遮阴程度正相关,遮阴处理Ci显著高于CK(P<0.05)。
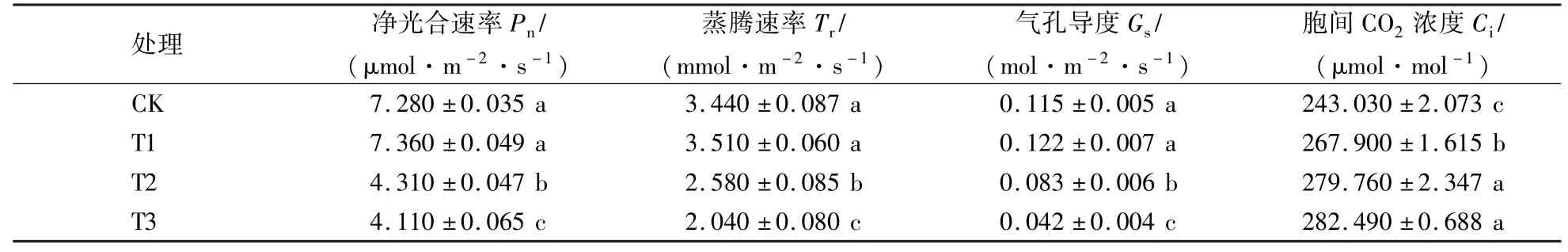
表3 遮阴对紫楠幼苗光合速率的影响Table 3 Effects of shading on photosynthetic rate of Phoebe sheareris seedlings
2.4 遮阴对相对电导率、丙二醛和渗透物质的影响
由表4可知,可溶性糖含量随遮阴程度的增大而不断下降,T1处理与CK差异不显著,而T2和T3处理显著低于CK(P<0.05)。随着遮阴程度的增大,紫楠幼苗叶片的相对电导率、丙二醛、游离脯氨酸和可溶性蛋白含量先下降后上升,处理间差异显著,T1处理最低。T1处理的相对电导率分别较CK和T3处理下降了53.33%和32.26%,丙二醛含量较CK和T3处理下降了47.51%和34.68%,游离脯氨酸含量较CK和T3处理下降了60.91%和27.99%,可溶性蛋白含量较CK和T3处理下降了73.26%和47.07%。
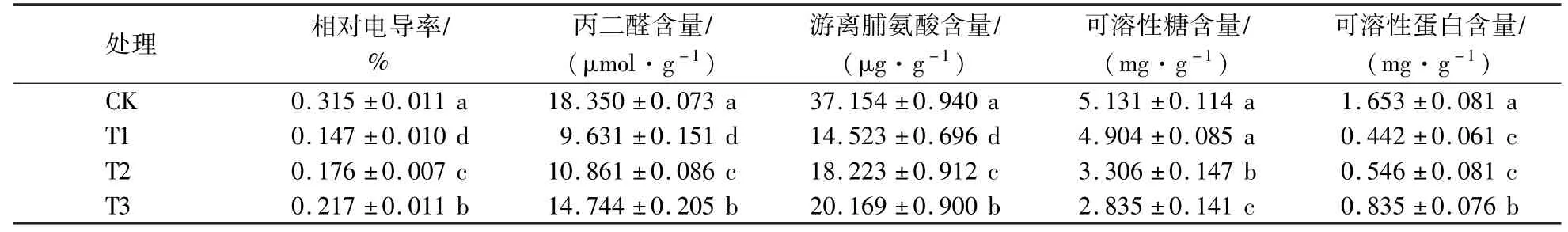
表4 遮阴对相对电导率、丙二醛和渗透物质的影响Table 4 Effects of shading on the relative conductivity,MDA and osmotic substance of Phoebe sheareris
2.5 遮阴对超氧化物歧化酶、过氧化物酶活性的影响
由表5可知,随着遮阴程度的增大,紫楠幼苗叶片的超氧化物歧化酶、过氧化物酶活性均呈先下降后上升趋势,T1处理最低,T1处理与CK和T3处理差异显著(P<0.05)。T1处理的超氧化物歧化酶活性分别较CK和T3处理下降了14.48%和10.58%,过氧化物酶活性则分别下降了51.03%和49.46%。
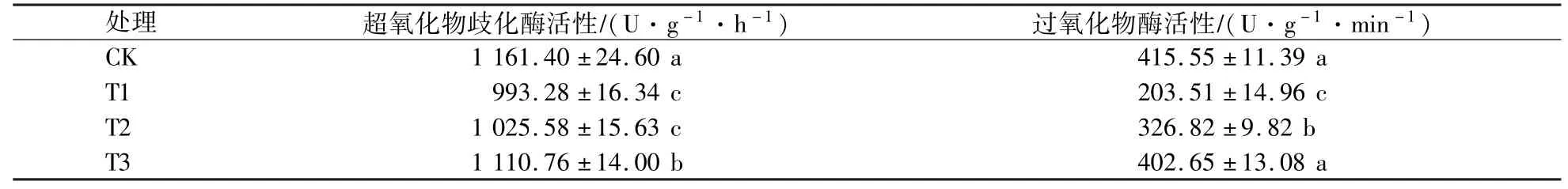
表5 遮阴对超氧化物歧化酶、过氧化物酶活性的影响Table 5 Effects of shading on the SOD and POD activity of Phoebe Sheareris
3 讨论与结论
梁俊林等[13]研究表明,遮荫处理下的桢楠、红椿和香樟的地径和苗高大部分均小于全光照处理。王慷林等[14]则认为,小省藤苗对遮阴的需求较高,遮阴90%生长最优。本试验中,与全光照相比,适度遮阴促进紫楠幼苗的苗高生长,遮阴度过高起抑制作用,而地径则随遮阴程度的增加而减小,这与代大川等[15]研究结果相似。适度遮阴能促进紫楠幼苗生长,究其原因,可能一方面是紫楠幼苗光能利用系统尚不完善,全光照超出了幼苗所能利用的光照强度范围;另一方面遮阴过度则导致光照不足,引起植物吸收光能转化为化学能减少,最终影响有机物的合成。
前人研究认为叶绿素a对自然光中的红光吸收较强,叶绿素b对植物吸收蓝紫光有促进作用,叶绿素a∕b是评价植物耐阴性的重要指标[16-17]。类胡萝卜素可以提高植物光能捕获能力,同时防御光破坏[18]。本研究发现,随着遮阴程度的增大,叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量均显著增加,而叶绿素a∕b比值下降,可能是遮阴阻挡了自然光中的红光部分,从而使得红光比例下降,较短波长的蓝光比例增加;紫楠幼苗通过提高自身光合色素含量,尤其是吸收蓝紫光的叶绿素b,充分利用光能,与张云等[19]对堇叶紫金牛的研究结果基本一致。
梁俊林等[13]认为,不同树种叶片光合速率对遮阴的反应不一致,红椿在全光照下的光合速率低于遮荫50%,而香樟和桢楠的光合速率下降。本研究中,随遮阴程度的增大,Ci不断上升,Gs呈先升高后显著下降趋势,各处理间Tr和Pn的趋势与Gs类似,表明中、重度遮阴导致叶片气孔迅速闭合,造成蒸腾速率与净光合速率均下降,而轻度遮阴则促进叶肉细胞光合作用,这与邓明明等[20]对米槁研究基本一致。
相对电导率、游离脯氨酸、可溶性糖、可溶性蛋白含量、SOD和POD活性是评价树种在各种胁迫条件下的重要指标[21-23];光胁迫条件下,MDA含量的增加幅度反映出膜系统的损伤程度[24];本研究中,随着遮阴程度的增加,相对电导率、丙二醛、游离脯氨酸、可溶性蛋白含量、SOD和POD活性均先降后升,T1处理最低,表明过强或过弱的光照都使紫楠幼苗受到胁迫,导致了相关生理指标的上升,这与郭品湘等[25]对双色木番茄的遮阴研究结果基本一致。随遮阴强度的不断增大,可溶性糖含量逐步下降,可能是幼苗光合直接产物的积累受遮阴影响导致,这与陈艺群等[26]对茄子的光胁迫结果一致。
综上所述,轻度遮阴(遮阴率40%)处理幼苗的生长优于全光照(CK)、中、重度遮阴(遮阴率60%、80%)处理,表明紫楠幼苗的生长在全光照(CK)、中重度遮阴条件下均受到了不同程度的胁迫。因此,为培育出更加优良健壮的紫楠苗木,建议采用适度的遮阴处理(以遮阴率40%为宜)。
