甘肃省临夏盆地渐新世—中中新世爪兽科和马科(奇蹄目,哺乳纲)化石*
2022-11-02沙卓菲苟诗皓刘傲然陈少坤
李 岩 沙卓菲 苟诗皓 张 彤 刘傲然,3 陈少坤
(1.甘肃省博物馆 兰州 730050;2.河北地质大学地球科学学院 石家庄 050031;3.河北省战略性关键矿产资源重点实验室 石家庄 050031;4.河北地质大学古生物研究院 石家庄 050031)
爪兽和安琪马化石在我国新生代堆积中都是很少见的。甘肃省博物馆与和政古动物化石博物馆在临夏盆地征集了一批此两类动物的化石,为临夏盆地渐新世和中中新世地层年代辨别提供了更多的依据。爪兽是地史时期出现过的最为奇特的哺乳动物之一,以爪状的末端指(趾)骨而有别于其他有蹄类动物,爪状的蹄骨用以抓取树枝,从而取食嫩枝和树叶(Coombs,1983)。安琪马的颊齿不同于晚期的马科动物,它的牙齿为低冠脊形齿,且无白垩质覆盖,是典型的食叶动物(McFadden,1992)。这两类动物都是重要的森林环境指示物种,在生物地层和古环境鉴别方面具有重要的意义。
1 地质背景
甘肃省临夏盆地从构造上讲是由青藏高原东北缘雷积山深大断裂、秦岭北深大断裂和祁连山东延余脉马衔山所围成的一个山间拗陷盆地(方小敏等,1997);盆地内保留了约30 Ma 以来连续的巨厚陆相沉积,从下往上依次为下渐新统他拉组、上渐新统椒子沟组、下中新统上庄组、中中新统东乡组和虎家梁组、上中新统柳树组、上新统何王家组和积石组以及更新统的午城黄土和马兰黄土(邓涛,2004;Deng et al.,2013)。
临夏盆地巨厚的沉积中蕴含了大量的哺乳动物化石,包括晚渐新世巨犀动物群、中中新世铲齿象动物群、晚中新世三趾马动物群和早更新世真马动物群,其中前两者中化石标本和种类相对较少。本文所述之河套裂爪兽(Schizotherium ordosium)产自椒子沟一带的砂岩和砂砾岩中,属椒子沟组下部。椒子沟组的命名地点位于东乡族自治县椒子沟,该地点的岩性地层已被详细描述(邱占祥等,1990),但因化石采集的原因,最初命名的椒子沟动物群中混入了来源不明的嵌齿象(Gomphotheriumsp.)化石,而被定为早中新世(邱占祥等,1990),后因牙沟动物群的发现和深入细致的地质调查而将之修订为晚渐新世(邱占祥等,2004)。虽然被修订为晚渐新世,椒子沟地点仍缺少可与牙沟相对比的时代标志性物种,此处描述的S.ordosium正可弥补这一缺陷。
裂爪兽(Schizotheriinae gen. et sp. indet.)和戈壁安琪马(Anchitherium gobiense)化石产自祖祖村上沟地点,异齿爪兽(Anisodonsp.)产自老沟地点,这两个地点都发现了铲齿象动物群化石,化石层位是典型的河流相砂砾岩,属虎家梁组。老沟地点的地层参考邓涛等(2004)文献,祖祖村上沟地点的地层可与老沟地点进行对比(未发表)。
文中所用爪兽类牙齿术语参考Chen et al.(2016),安琪马牙齿术语参考Sánchez et al.(1998)。缩略语(ZR,甘肃省博物馆自然部化石标本编号;HMV,和政古动物化石博物馆化石标本编号)。测量数据单位均为mm。
2 系统描述
(1)哺乳纲 MAMMALIA Linnaeus,1875;奇蹄目 PERISSODACTYLA Owen,1848;爪兽科 CHALICOTHERIIDAE Gill,1872;裂爪兽亚科 SCHIZOTHERIINAE Holland and Peterson,1914;裂爪兽属SchizotheriumGervais,1876;河套裂爪兽Schizotherium ordosiumHu,1959(图 1,表1)。

图1 椒子沟地点的Schizotherium ordosium 化石Fig.1 Fossils of Schizotherium ordosium from the Jiaozigou

表1 椒子沟的Schizotherium 下臼齿尺寸/mm 及与相关属种的比较Table 1 Measurements of lower molars of Schizotherium/mm from Jiaozigou and comparisons with related species
材料:ZR000513,残破左p3 一件、残破右p4 一件、右m1 两件、右m2 一件、残破左m3 一件、右m3 一件、残破左M1 一件,另有不能鉴定位置的牙齿碎块两件;ZR000473,右m1 一件、右m2 两件、右m3 两件、右P4 两件、右M2/3 残块一件。
产地与层位:甘肃省东乡族自治县椒子沟,椒子沟组底部的厚层砂砾岩,晚渐新世塔本布鲁克期。
描述:P4 两件,均有较深的磨蚀,且有不同程度的破损。冠面轮廓近正方形。前、后附尖破损,无中附尖发育的痕迹,因此外脊应不具有W 形态。前尖较后尖更高一些,原尖呈圆锥形,位于舌侧中间位置,无原小尖的痕迹。原脊比后脊更窄一些,二者分别与原尖的前后缘相连接,使中谷封闭。中谷内无小刺等结构。齿带在前、后和舌侧存在,其中前、后齿带非常发育,并在舌侧的前、后角处形成圆角,在舌侧连续,无中断。
M1 一件,仅保留了外脊和部分后脊,外脊长24.5 mm,可见较为发育的前尖肋和较弱的后尖肋,前附尖较中附尖更弱,后附尖不发育。后脊起自中附尖和后尖之间外脊的中间部位。后凹较深,呈V 型。唇侧齿带较弱,后齿带发育,在后凹开口处形成棚架状。
M2/3 一件,仅保留前舌侧部分。原小尖较高,原脊起自原尖顶端的前舌侧。原尖孤锥状,舌侧陡直。前齿带发育,呈棚架状。
p3 一件,保留中段部分。可见下后附尖附于下后尖的后唇侧,下后附尖较下后尖更为矮小。
p4 一件,下三角座和部分下后脊保存。下三角座V 型。下前尖部分亦有破损,但仍可看出其较低,中等磨蚀的下原尖较下后尖更低,下后尖和下后附尖并排紧靠在一起,下后尖较下后附尖更粗壮一些。可见微弱的前唇侧齿带存在。
m1 三件,均有较深的磨蚀。双V 型,下三角座较下跟座稍宽或相当,但长度不及后者。在各尖之中,下前尖位置较低,下后尖最高,下后附尖靠在下后尖的后唇侧,比下后尖更低一些。下前脊在前舌侧有明显的内弯,下前脊与下后脊走向接近平行,下原脊与下次脊走向接近平行,后两者的走向都与牙齿长轴走向接近垂直。齿带仅在前唇侧和后唇侧存在,且不甚发育。
m2 三件,下三角座前部均有不同程度的破损。下跟座V 型。在中等磨蚀的标本上,下后尖和下后附尖并排紧靠在一起,二者粗壮程度相当,下后附尖稍低于下后尖,舌侧齿尖的磨蚀速度远低于唇侧齿尖。后谷和外谷均为V 型,前者宽而浅,后者窄而深。齿带仅在后侧发育,在其中一件标本上还形成类似于下次小尖的结构。
m3 四件,磨蚀程度不等。下三角座接近U 型,下跟座为V 型,前者的长度和宽度均不及后者。舌侧齿尖的磨蚀速度远低于唇侧齿尖,下前尖位置相对较高,下后尖和下后附尖结合紧密,发育程度相当,二者紧靠在一起,在磨蚀后无显著的分隔。下前脊端部有明显的内弯,前半部分走向与牙齿长轴方向垂直,向后呈圆角状延伸至下原尖。下原脊和下次脊走向接近平行,都与牙齿长轴方向斜交。前谷、后谷和外谷均为较开阔的V 型,前谷深度不及后谷。齿带仅在前唇侧和后侧有比较弱的存在,无明显的下次小尖。
比较与讨论:从牙齿的尺寸、冠高、原小尖以及下后附尖的形态来看,这些标本属小型的裂爪兽亚科(Schizotheriinae),即渐新世的裂爪兽属(Schizotherium)或鲍氏爪兽(Borissiakia)。Coombs(1978)对Schizotherium的属征做出了明确的定义,在牙齿上表现为: 臼齿加长趋势不及本亚科其它属; 上臼齿近方形, 外脊较爪兽亚科(Chalicotheriinae)更高更陡直,但不及裂爪兽亚科其它属;上臼齿有小刺而无前刺;m3有下次小尖存在。Borissiakia比Schizotherium更大,但比裂爪兽亚科其它属更小,臼齿加长趋势更明显,m3 无下次小尖。椒子沟的这些爪兽m3 虽无明显的下次小尖,但臼齿的加长趋势并不明显,尺寸也显著的小于Borissiakia。因此,本文将椒子沟的爪兽化石鉴定为Schizotherium。
Schizotherium中有6 个种:S.priscum、S.turgicum、S.avitum、S.chucuae、S.ordosium和S. nabanense。Coombs(1978)详细总结了Schizotherium各种的牙齿特征,椒子沟爪兽m3 下三角座稍窄于下跟座,与S. avitum相似;m2 和m3 的长度超过最大宽度的2 倍,与S. avitum、S. chucuae、S. ordosium和S. nabanense相似;下后附尖强壮,与下后尖有明显的分隔,与S. priscum、S. turgicum、S. ordosium正型标本和S. nabanense相似或一致;下次小尖不明显,即非常弱,与S. chucuae相似。从这些对比来看,椒子沟的爪兽似乎与所有已知的Schizotherium均有区别,但若从个体大小来看,能与椒子沟的爪兽化石相比的仅有S.ordosium,因它以个体显著较大为典型特征(邱占祥,1998)。
从表1 可以看出,椒子沟的爪兽m3 尺寸虽然大于内蒙古河套地区出土的该种正型标本(胡长康,1959),但与兰州盆地(邱占祥,1998)、党河下游(王伴月,2004)以及临夏盆地牙沟地点(邱占祥,2004)的S. ordosium相当。由于椒子沟的爪兽与兰州盆地党、河下游、牙沟的该类化石并无明显的区别,因此,本文将椒子沟的爪兽鉴定为S. ordosium。同时也应看到,椒子沟爪兽的m1 尺寸小于这些地点,说明它的前部臼齿变小,后部臼齿变大,这是一个进步的特征。
Schizotherium ordosium的时代为早渐新世晚期(?)至晚渐新世,而椒子沟地点的该类化石与其它地点相比,有些许进步之处,其时代更可能为晚渐新世。椒子沟地点最初因有长鼻类化石(混入)而被定为早中新世(邱占祥,1990),之后因为在相邻的牙沟发现了明确的晚渐新世动物群,又被修改为晚渐新世(邱占祥,2004),此次发现的S.ordosium化石进一步明确了其时代属性。
椒子沟和牙沟动物群中,除林栖型的Schizotherium,亦有擅长奔跑的阿尔丁犀(Ardynia)和大型的准噶尔巨犀(Dzungariotherium)、副巨犀(Paraceratherium)等,说明在渐新世晚期,临夏盆地是以疏林环境为主。
(2)裂爪兽亚科(未定属种)Schizotheriinae gen.et sp.indet.(图2A~图2B)。
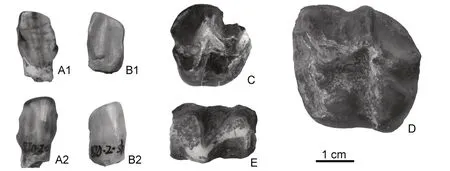
图2 临夏盆地中中新世的Schizotheriinae gen.et sp.indet(.A~B)和Anisodon sp(.C~E)Fig.2 Schizotheriinae gen.et sp.indet.(A~B)and Anisodon sp.(C~E)from the Middle Miocene of the Linxia Basin
材料:ZR000476,右下门齿两件,可能是i1 和i2。
产地及层位:甘肃省东乡族自治县祖祖村上沟,中中新统虎家梁组,通古尔期(约相当于欧洲的MN 6~8)。
描述:这两件门齿的尺寸有一定的差异,且磨蚀方向也有不同,说明它们应为同侧的不同位置,很可能是i1 和i2,表明这种爪兽至少有两对下门齿。铲型,前端平,几乎未磨蚀,两侧缘右不同程度的突起,其中近中侧的突起更为发育。舌面双凹,唇面圆隆。二者长宽分别为:9.7 mm×6.8 mm(?i2)和10.5 mm×7.7 mm(?i1)。
比较与讨论:爪兽类的门齿发现非常少,却是容易鉴别的。在爪兽亚科中,异齿爪兽(Anisodon)的下门齿很弱小,呈单尖状或者说豆状(Zapfe,1979),与上沟地点的爪兽差别巨大;裂爪兽亚科中,与后裂爪兽(Metaschizotherium)的猪型门齿(Coombs,2009)有显著区别,但Schizotherium以及石爪兽(Moropus)下门齿形态(Holland and Peterson,1914;邱占祥,1998)与上沟地点的这两件爪兽门齿均非常相似,仅尺寸稍偏更大,若再考虑时代差异,则当与Moropus接近。本文将之鉴定为裂爪兽亚科,不定属种,仅表明中中新世时期裂爪兽亚科在这一地区的存在。
我国中中新世的裂爪兽化石此前仅记录于内蒙古通古尔地区,以零星头后骨骼为代表(Colbert,1934;Xue and Coombs,1985;Fahlke and Coombs,2009;Chen et al.,2012),亦不能鉴定属种。
(3) 爪兽亚科 Chalicotheriinae Gill,1872;异齿爪兽属AnisodonLartet, 1851;异齿爪兽(未定种)Anisodonsp(.图2 C~图2E)。
材料:HMV 2050,右DP4;HMV 2051,左M2;HMV 2052,右m1。
产地与层位:甘肃省和政县老沟地点,中中新统虎家梁组,通古尔期晚期(约相当于欧洲的MN 8)。
描述:DP4 低冠,中等磨蚀,从其较薄的牙釉质和尺寸来看,可将之鉴定为乳齿。前附尖破损,中附尖发育强烈,后附尖很弱;前尖顶端破损,原小尖很小,位于前尖的前舌侧,后尖向舌侧倾斜程度较大,原尖呈圆锥形,较次尖位低,次尖为三角锥形。外脊呈W 型,后部的V 型较前面者更深,原脊起自前尖前部,向舌侧延伸至原尖的前唇侧,后脊直,起自外脊近后尖处,止于次尖顶部。中谷宽阔,底部呈V 型;后凹呈U型。齿带在牙齿周围均存在,但发育程度很弱。L×W:19.7 mm×21.7 mm。
M2 冠面方形,低冠,稍有磨蚀。这件标本与上述DP4 多有相似之处,亦存在很多区别。前附尖发育程度不及中附尖;后附尖较DP4 更发育些。原小尖较大,其横截面为三角形,位于前尖的舌侧;原尖的舌侧壁陡直,其它各面则坡度较大。原脊很弱,止于原小尖。中谷开阔,原尖和原小尖之间的开口呈V 型;后凹收缩程度不及DP4。L×W:33.5 mm×31.8 mm。
m1 的前壁破损,磨损很重。下前尖和下内尖很小,下后尖的后部较宽阔,可能预示着下后附尖的存在。下前脊和下次脊很窄,二者的走向几乎都与牙齿长轴相垂直。下跟座较长,但宽度不及下三角座,下三角座呈V 型,下跟座则为U 型。前谷很小,后谷则开阔的多。牙齿四周的齿带均较发育。L×aW+pW:23.7 mm×14.4 mm+13.5 mm。
比较与讨论:从牙齿的齿冠轮廓及高度,可以判断这几件标本当属低冠的爪兽亚科(Chalicotheriinae), 该 亚 科 现 有 6 个 属 ,Butleria、Kalimantsia、Chalicotherium、Anisodon、Nestoritherium和Hesperotherium(陈少坤等,2012)。从大小来看,这些标本可与“Chalicotherium”pilgrimi和Anisodon相比较。“C”.pilgrimi在本亚科中是一个很原始的种类,目前仅发现于Siwaliks 早中新世的地层中(Forster-Cooper,1920),它的上臼齿唇侧齿尖内倾程度很低,老沟的爪兽与之差距甚远。在Anisodon属中,老沟标本的尺寸与青藏高原西南部Siwalik 的A. salinus、东南部云南等地的A. yuanmouensis都很接近,明显小于欧洲的A. grande和A. macedonicus,形态特征也在前两者的变异范围(Chen et al.,2016)之内。因此,此处将老沟的爪兽鉴定为Anisodonsp.。
在临夏盆地的老沟动物群中,谷氏铲齿象(Platybelodon grangeri)是优势的种类,它在青藏高原北部的多个地点有大量的发现,然而在南亚次大陆从未见其身影(Deng et al.,2007),这可能表明在中中新世,青藏高原已经隆升到一定高度,从而阻止了大型动物的南北迁徙。然而,在临夏盆地和云南、缅甸和南亚的西瓦利克(Siwalik)发现了Anisodon(Colbert,1935;Pickford,1982;Chavasseau et al.,2010;Chen et al.,2016),表明中中新世时期的青藏高原周边,可能存在某种程度的动物群交流或未知的动物迁徙路线。
在过去,我国北方中中新世的爪兽化石,除上文提及的通古尔地区的不能鉴定属种的裂爪兽化石外,均被鉴定为短吻爪兽(Chalicotherium brevirostris)或河北爪兽(C.hebeiense)(Colbert, 1934; 胡 长 康 , 1959; Wang and Wang, 2001; Liu and Zhang,2012;陈少坤等,2013;陈少坤等,2015),它们都显著较Anisodon粗壮。老沟Anisodon化石的发现说明,在我国北方中中新世时期,爪兽类的物种多样性比我们之前认识的要高。
(4)马科 Equidae Gray,1821;安琪马亚科Anchitheriinae Leidy,1869;安琪马属AnchitheriumMeyer,1834;戈壁安琪马Anchitherium gobienseColbert,1939(图 3,表 2)。
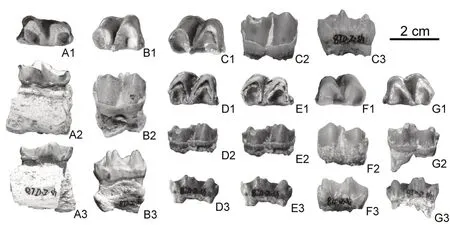
图3 临夏盆地上沟地点的Anchitherium gobiense 化石Fig.3 Fossils of Anchitherium gobiense from Shanggou locality in the Linxia Basin

表2 上沟地点的Anchitherium gobiense 尺寸/mm 及与相关属种的比较Table 2 Measurements of Anchitherium gobiense/mm from Shanggou and comparisons with related species
材料:ZR000476,左p2 一件、右p3 一件、右m1 一件、左m2 两件、右m2 两件。
产地与层位:甘肃省东乡族自治县祖祖村上沟,中中新统虎家梁组,通古尔期(约相当于欧洲的MN 6~8)。
描述:p2 一件,中等磨蚀。双叶型显著,下三角座较下跟座更长,但稍窄。下前尖显著,下后尖和下后附尖发育程度相当,唇侧齿尖的高度低于舌侧齿尖。下前脊窄,前端有内弯,下原脊走向与牙齿长轴方向垂直,下后脊较长,下次脊部位有破损致使下次小尖的状态不明。前谷接近U 型,后谷和外谷则为斜V 型。齿带在前、后和唇侧较为发育,舌侧齿带缺失。
p3 一件,基本未磨蚀。下三角座U 型,下跟座接近V 型,前者较后者更宽更短。下前尖低矮萎缩;下原尖稍低于下后尖而略高于下次尖;下后尖和下后附尖并排紧靠,中间有明显的纵沟分隔,二者发育程度相当;下次尖的位置较下原尖更靠近唇侧;下内尖为三棱椎形,在下内尖之后存在一个显著的下次小尖。下前脊前端有明显的内弯,后端走向与下后脊基本平行;下原脊和下次脊走向接近平行,均与牙齿长轴方向斜交。前谷浅U 型,后谷为更宽更深的V 型,外谷为斜向前的窄V 型。齿带在前、后和唇侧发育,在舌侧则完全缺失。
m1 一件,稍有磨蚀。下三角座U 型,下跟座接近V 型,前者较后者稍更宽更短。下前尖较低;下原尖与下后尖高度相当,二者均高于下前尖;下后尖和下后附尖接近愈合,下后附尖极弱;下次尖的顶部破损;下内尖为三棱椎形;下次小尖发育,稍低于下内尖。下前脊前端有明显的内弯,下前脊的后端和下原脊走向接近弧形;下后脊较长,起自下后尖的唇后侧,与下次脊呈圆角状相接。前、后谷和外谷,以及齿带的形态与前臼齿基本一致。
m2 四件,均完整保存。下三角座更接近V 型,下跟座更加的宽,但长度与下三角座相似。下后尖和下后附尖的形态存在变异,在三件标本上,二者并排紧靠,中间有明显分隔,下后附尖稍弱于下后尖,而在另外一件标本上,二者更接近愈合,与m1 一致。其余结构形态均与m1 接近。
比较与讨论:关于旧大陆安琪马亚科(Anchitheriinae)的分类已经进行过很多次的讨论(Abusch-Siewert,1983;Sànchez et al.,1998;孙博阳等,2021)。在我国,安琪马亚科的化石非常稀少,叶捷等将我国中中新世的安琪马类全部归入Anchitherium gobiense(除南京方山的A. aurelianense),而晚中新世三趾马动物群中的安琪马类化石全部归入中华马属(Sinohippus)(叶捷等,2005)。侯素宽等认真比较了Sinohippus和Anchitherium(尤其是A. aurelianense)的区别,认为它们在下颊齿上的区别主要表现在下次小尖的强弱程度、外谷向前倾斜与否、舌侧齿叶外壁圆隆与否、舌侧齿带的有无等方面(侯素宽等,2007)。从尺寸的大小来看,上沟地点标本显著较Sinohippus纤弱,而在A. gobiense和A. aurelianense的变异范围之内。再从下颊齿的形态来看,其下次小尖明显,外谷向前倾斜,舌侧齿叶外壁不及Sinohippus圆隆,舌侧齿带缺失,可与A. gobiense相比,而不同于Sinohippus和A. aurelianense。与临夏盆地老沟地点的A. gobiense相比,其形态非常接近(仅m1 标本可直接对比),但尺寸稍大于老沟地点者。
Anchitherium gobiense还发现于内蒙古通古尔(Colbert,1939)、南京方山(周明镇等,1956)、湖北房县(阎德发,1979)和新疆准格尔盆地(叶捷等,2005),时代为通古尔期,约等同于MN 6~8。
中中新世通古尔期正值全球气候适宜期,繁盛的铲齿象动物群指示了温暖湿润的森林环境(邓涛,2016)。老沟和上沟地点发现了Schizotheriinae gen. et sp. indet.、Anisodonsp.和Anchitherium gobiense,前两者奔跑能力弱,适合在树丛中用前肢勾取嫩枝或树叶来食用,Anchitherium牙齿低冠,适宜取食柔软的树叶,它们都是适宜温湿森林环境的动物。
3 结 论
甘肃省临夏盆地晚渐新世地层见于牙沟和椒子沟等地,椒子沟Schizotherium ordosium的发现增加了牙沟和椒子沟两个地点的共有属种,更证实了椒子沟组的时代为晚渐新世。
在中中新世的我国北方,爪兽类的生物多样性是比较高的,临夏盆地至少存在两种类型的爪兽,分属不同的亚科,其中裂爪兽亚科标本数量很少,不能鉴定至属,仅表明此类动物的存在;爪兽亚科的Anisodonsp.与我国北方同时期的爪兽有明显区别,而与青藏高原西南和东南部的A. salinus、A. yuanmouensis更接近,说明在中中新世,青藏高原周边存在某种程度的动物交流或未知的动物迁徙路线。
报导了临夏盆地中中新世的安琪马化石,根据其较为纤细、下次小尖明显和舌侧齿带缺失等特征,可以将其鉴定为Anchitherium gobiense,是通古尔期的代表物种之一。老沟地点和祖祖村上沟地点同属通古尔期虎家梁组。
爪兽类和安琪马类化石的发现说明,在临夏盆地晚渐新世以疏林环境为主,而中中新世温湿森林更加普遍。
