不同烟草品种上部烟叶的物质变化规律①
2022-10-29易美琴陈文文徐小洪冉茂秦平伟刘安洋高天科夏庆友王根洪赵敏
易美琴, 陈文文, 徐小洪, 冉茂, 秦平伟,刘安洋, 高天科, 夏庆友, 王根洪, 赵敏
1.西南大学 家蚕基因组生物学国家重点实验室,重庆 400715;2.西南大学 前沿交叉学科研究院生物学研究中心,重庆 400715;3.中国烟草总公司重庆市公司 烟草科学研究所,重庆 400715;4.中国烟草总公司重庆市公司 彭水分公司,重庆 彭水 409600
烟叶本身的烘烤难易程度(易烤性)及耐受程度(耐烤性)统称为烟叶的烘烤特性[1-3]. 烤烟的烘烤特性受到多方面因素的影响[4-8], 但最重要的影响因素是遗传基础也即品种差异[9], 且不同基因型烤烟烘烤特性差异有统计学意义[10].
在重庆地区, 存在“云烟87”和“云烟85”上部烟叶相对易烤且耐烤, 而“K326”和“红花大金元”难烘烤的现象. 本研究选定不同烘烤特性品种的烟草的上部烟叶, 对其在烘烤过程中烟叶的含水率、 颜色、 质体色素、 糖类物质、 含氮化合物的动态变化与叶绿素酶、 多酚氧化酶活性等常规化学指标的动态变化规律进行分析, 旨在为解决不同品种上部烟叶的难烘烤问题提供新的科学思路, 从而减少烘烤带来的损失, 为改善烟叶烘烤特性, 提高烤后烟叶品质提供理论支持.
1 材料与方法
1.1 试验材料
供试品种为“云烟87”(Y87)、 “云烟85”(Y85)、 “红花大金元”(HD)和“K326”, 种植于中国烟草总公司重庆市公司彭水分公司的润溪烟站基地. 该基地土壤肥力合适, 田间烟叶管理按照常规的烟叶生成模式进行种植管理. 烟叶发育正常, 长势良好. 供试烟叶为上部适熟烟叶. 4个品种烟叶置于同一烤房中, 采用适用于“云烟87”的三段六步式烘烤工艺进行烘烤, 在此工艺基础上根据烟叶状态做出适当调整. 主要设备为密集烤房控制器YJHK-01等.
1.2 试验设计
试验主要为烘烤试验和烤后烟叶生理生化测定.
1) 烘烤试验: 采收“云烟87”“云烟85”“红花大金元”和“K326”的上部第四位烟叶于密集烤房中进行烘烤, 试验采用常规的挂杆烘烤和三段六步式烘烤工艺(图1), 在烘烤过程中对烘烤前RT(Room Temperature)、 变黄期(38 ℃与42 ℃下烟叶8~9成变黄, 叶脉仍为绿色)、 定色期(48 ℃与56 ℃下黄片黄筋, 叶片干燥)、 干筋期(62 ℃下全炉烟叶, 主脉干燥)及烘烤后(68 ℃)5个时期不同时间点, 选取每个温度对应阶段的中间节点时间进行取样, 观察烟叶颜色变化.
2) 烟叶生理生化测定 : 采取随机取样原则, 选取烟叶叶中部位进行测定. 每3片叶为一份样本, 剪碎混匀并用锡纸包裹放入干冰中, 回到实验室后放于-80 ℃冰箱保存待用.
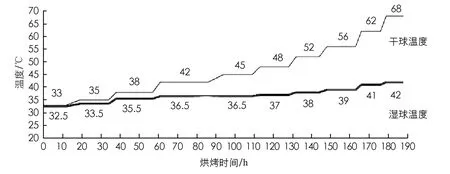
图1 三段六步式烘烤工艺温湿度设置条件
1.3 测定项目与方法
1.3.1 烟叶含水量测定
烟叶的含水量变化采用杀青烘干法[11]测定, 计算叶片及主叶脉的相对含水率, 公式如下:
W=(G1-G2)/G1×100%
其中:W为相对含水率,G1为湿烟草质量,G2为全干烟草质量.
1.3.2 化学成分含量测定
叶绿素、 类胡萝卜素含量的测定参照文献[12]的方法, 采用乙醇浸提与比色法来检测.
总氮含量: 采用硫酸-双氧水消解与凯氏定氮法测定植物全氮含量.
粗蛋白含量: 采用硫酸-催化剂消解与凯氏定氮法测定植物粗蛋白含量.
还原糖、 总糖含量: 分别采用G-CLONE还原糖含量、 总糖含量检测试剂盒(重庆康坭商贸有限公司)进行测定. 所用仪器主要为凯氏定氮仪、 梅特勒 ML204型万分之一天平、 分光光度计(T6-新世纪, 普析)、 Synergy H4全功能酶标仪(美国伯腾仪器有限公司).
1.3.3 相关酶活性测定
超氧化物歧化酶(SOD)、 过氧化物酶(POD)活性: 采用试剂盒测定超氧化物歧化酶(SOD)、 过氧化物酶(POD)活性(南京陌凡生物科技有限公司).
淀粉酶的活性: 采用淀粉酶试剂盒(南京建成生物工程研究所)测定.
多酚氧化酶(PPO)的活性: 采用多酚氧化酶(PPO)试剂盒(上海索桥生物科技有限公司).
叶绿素酶活性: 采用植物叶绿素酶ELISA试剂盒(重庆康坭商贸有限公司)测定.
1.4 数据分析
运用Microsoft Excel进行试验结果统计, 运用GraphPad Prism 6, Illustrator进行绘图.
2 结果与分析
2.1 烘烤过程中质体色素含量、 叶绿素酶的活性变化
随着时间和温度的增加, 烟叶由黄绿色逐渐变黄, 而烟叶的质体色素含量可以反映叶片的颜色变化, 图2是4个供试品种上部烟叶烘烤过程中的叶绿素a(图2a)、 类胡萝卜素(图2b)相关含量和叶绿素酶活性(图2c)变化图.
新鲜烟叶中“红花大金元”的叶绿素含量最高, 在烘烤过程中各品种烟叶叶绿素含量都呈现逐渐下降的趋势, 且在变黄前期(38 ℃前)下降速度较快, 此阶段烟叶从叶尖开始变黄, 变黄长度约10 cm, 烟叶总计变黄6~7成; 42 ℃时烟叶总计变黄9~10成, 主叶脉发软, 整体呈现为黄片青筋, 与“云烟87”“云烟85”相比, “红花大金元”“K326”的叶基部位叶片仍为黄青色且叶脉更绿, “云烟87”在42 ℃时叶绿素含量降到最低, 而“红花大金元”的叶绿素含量依旧高于其他3个品种; “云烟85”“K326”在48 ℃时, 叶绿素含量最低, 之后所有品种的叶绿素含量都有所上升, 这可能是有机试剂提取叶绿素产生的误差, 但总体上叶绿素含量在烘烤过后是显著降低的, 并且从变化趋势上来看, “红花大金元”和“K326”的叶绿素含量降解速率低于“云烟87”“云烟85”.
由图2b可以看出, 从烘烤阶段开始到定色期前期(48 ℃), 由叶绿素降解转变为类胡萝卜素, 且其含量逐渐增高, “红花大金元”的类胡萝卜素含量最高. 而从定色期后期到干筋期阶段, 类胡萝卜素含量有所下降, 可能是其进一步生成了芳香类化合物. 在整个烘烤过程中, “红花大金元”的类胡萝卜素含量比其他品种高, 但是其叶绿素含量下降相对更慢, 这可能是导致其烘烤后杂色烟占比较大的原因之一.

图2 烘烤过程中烟叶的质体色素含量、 叶绿素酶活性变化
我们重点关注了变黄阶段的叶绿素酶活性的变化. 在新鲜叶片中, “云烟87”的叶绿素酶活性最高, “红花大金元”其次, “云烟85”和“K326”最低, 二者差异不显著. 在烘烤过程中, “K326”的叶绿素酶活性表现出逐渐升高的趋势; “红花大金元”在42 ℃之前酶活性缓慢下降, 在42 ℃之后略微升高; 在38 ℃时, “云烟87”“云烟85”的叶绿素酶活性较低, 随后随着温度上升而升高, 进入变黄后期(42 ℃)时, “云烟87”“云烟85”的叶绿素酶活性显著高于“红花大金元”“K326”, 而“红花大金元”酶活性最低.
从叶绿素、 类胡萝卜素含量变化趋势可知, 在烘烤过程中叶绿素酶活性和烟叶的叶绿素含量成反比, 和叶片变黄程度成正比, 且不同品种间的叶绿素酶活性的高低也可能影响烟叶的变黄难易程度, 进而影响烟叶的易烤性及其烤后烟叶品质.
2.2 烘烤过程中糖类物质的含量及淀粉酶活性变化
糖类物质是烟草光合产生的重要化学物质, 在烟草的干重中占比25%~50%[13], 按结构可分为单糖、 低聚糖和多糖[14]. 烟草在生长过程中会积累多糖, 包括淀粉、 纤维素、 果胶等, 在烟叶收割后的调制发酵、 烘烤过程中, 会降解为单糖, 如葡萄糖、 果糖等还原糖. 烟叶中的糖含量显著影响烟叶的气味和品质[15].
在鲜烟叶中, “红花大金元”的还原糖(图3a)含量最低, 烘烤开始后, “红花大金元”和“云烟85”的还原糖含量在38 ℃前呈显著升高的趋势, 随着温度的变化, 两个品种的还原糖开始降解, “红花大金元”一直处于降低的变化过程, 而“云烟85”在48~56 ℃间又再一次发生了较为显著的上升随后急剧下降直到烘烤结束; “K326”烟叶中的还原糖含量在42 ℃前呈现出较为平缓的变化, 42~48 ℃间急剧增加, 随后还原糖含量随温度的升高而降低; “云烟87”的还原糖含量在整个烘烤阶段都处于略微降低或略为升高的变化中, 仅在62~68 ℃间呈明显下降趋势.
总糖(图3b)即还原性的葡萄糖, 包括果糖、 乳糖、 戊糖、 蔗糖、 麦芽糖以及能部分水解的淀粉等, 它们能在特定条件下被水解成还原性单糖[16]. 从图3b中可以发现, 4个品种新鲜烟叶的总糖含量由高到低依次为“云烟85”“云烟87”“K326”“红花大金元”. 烘烤过程中烟叶总糖的变化趋势表现为先下降后上升. 在42℃变黄阶段之前, 总糖含量整体处于250 mg/g以下, “云烟87”“云烟85”的总糖含量要显著高于“红花大金元”和“K326”. 到了定色期后, 由于淀粉等大分子糖类物质经过高温降解, 总糖含量逐渐增大, 而“红花大金元”的总糖含量总体上最低, 这可能是淀粉等大分子糖类分解不充分所致, 而其他3个品种的总糖含量差异不显著.
淀粉酶作用于α-1,4-糖苷键和α-1,6-糖苷键, 降解产物以麦芽糖为主, 还有少量的麦芽三糖、 葡萄糖、 α-极限糊精等[17].
从整体上看, 如图3c, 在烘烤过程中, 除38℃时“红花大金元”的总淀粉酶活性略微升高外, 其他3个品种的总淀粉酶活性均有所降低. 48 ℃后的酶活性表现从高到低依次为“红花大金元”“云烟85”“K326”“云烟87”. 淀粉酶将淀粉降解为单糖, 烟叶中的还原糖、 总糖含量升高. 烤后烟叶中“红花大金元”的糖含量最低. 淀粉等大分子糖类物质可能是影响烟叶烘烤特性的关键因素.

图3 烘烤过程中烟叶的糖类物质含量及淀粉酶活性变化
2.3 烘烤过程中粗蛋白、 总氮、 烟碱的含量变化
烟草中具有多种含氮类化合物, 包括蛋白质、 生物碱、 游离氨基酸、 硝酸盐等, 其含量会影响烟叶烘烤特性, 同时也影响烟叶的品质及吸烟者的健康[18].
根据烤烟化学指标评价标准, 烤后烟叶的总氮含量范围在1.5%~3.5%之间并且总氮含量小于1.5%时吃味平淡, 大于3.5%则燃烧时产生浓烈辛辣的烟气, 刺激性较大, 最适值为2.5%[19]. 品质良好的烟叶粗蛋白含量应小于10%, 含量在10%~15%之间的烟叶品质会变差, 如果大于15%的话, 烟叶品质会更差[20]. 烟碱含量范围宜在1.5%~3.5%之间, 在2%左右最好, 含量过低则吃味平淡劲头不足, 过高则劲头过强, 烟味发苦[21].
图4是烟叶烘烤过程中粗蛋白(图4a)、 总氮(图4b)、 烟碱(图4c)的含量变化示意图, 在烘烤前及烘烤整个过程中, 难烘烤品种“K326”和“红花大金元”的粗蛋白含量均整体高于易烘烤品种“云烟87”和“云烟85”. 新鲜烟叶中“红花大金元”和“K326”的总氮含量处于1.5%以上, 显著高于“云烟85”和“云烟87”. 在整个烘烤阶段, 4个品种的总氮含量以不同的变化趋势升高, 其中“K326”“红花大金元”的总氮含量高于“云烟87”“云烟85”. 图4c结果显示, 烤前4个品种的烟碱含量在1.5%~2%之间, 在烘烤过程中含量均逐渐增加, 烤后含量处于2%~2.5%之间, 整体上差异不显著.
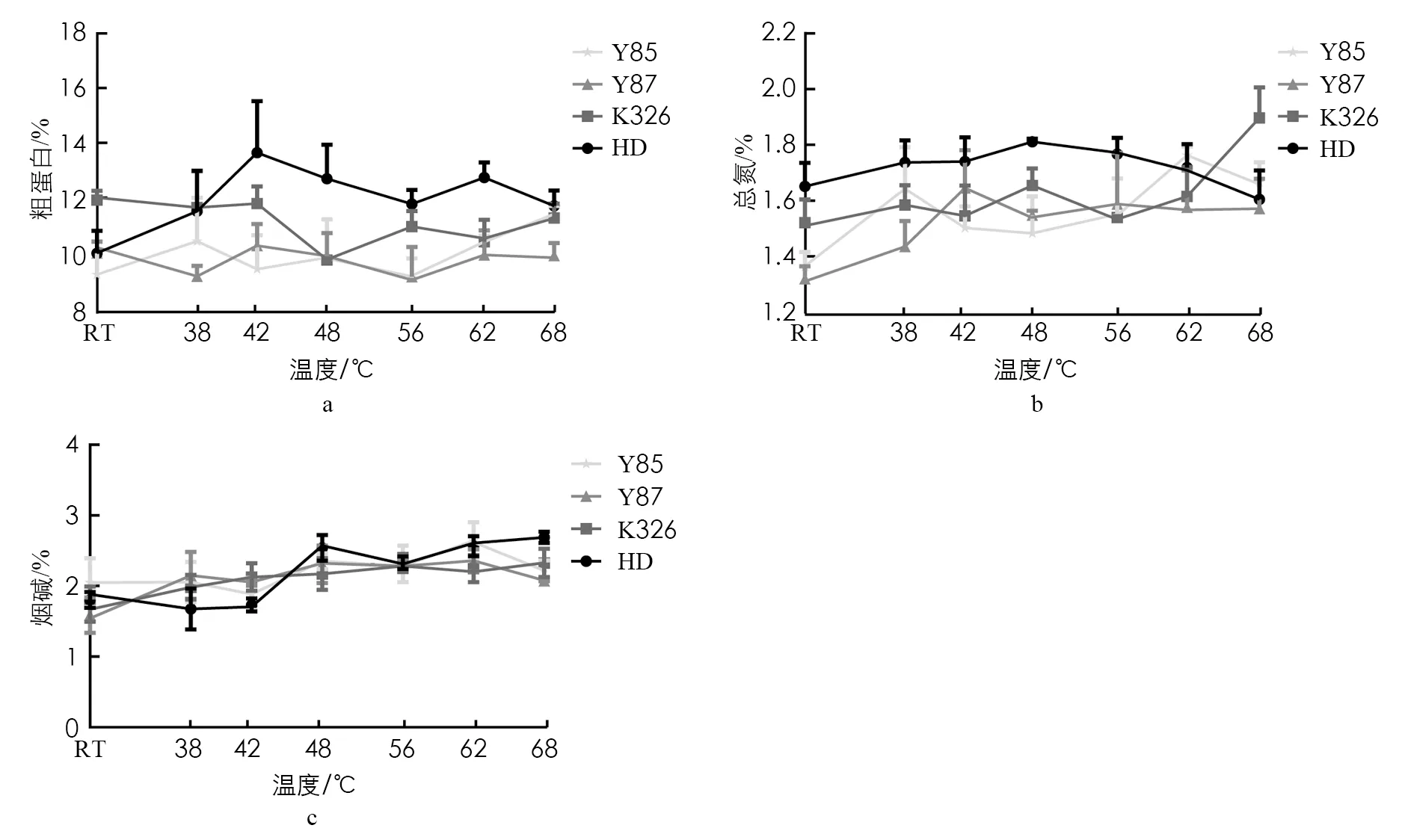
图4 烘烤过程中烟叶的粗蛋白、 总氮、 烟碱含量变化
2.4 烘烤过程中烟叶的含水量变化
供试4个品种的烟叶在烘烤前叶片、 主叶脉含水率相当, 烘烤过程中呈逐渐降低的趋势. 38℃变黄阶段前期, “红花大金元”的叶片含水率下降趋势最快, “云烟87”“云烟85”其次, “K326”最慢; “红花大金元”的主叶脉含水率下降也最快, 其他3个品种下降趋势相似. 38~42 ℃变黄中期, “红花大金元”叶片含水率虽然低但下降趋势与其他3个品种相比较为平缓; 42~48 ℃变黄中后期以及定色前期, “云烟87”“云烟85”叶片含水率下降更快; 而在42~48 ℃期间, 4个品种的主叶脉含水率又一次急速下降, 但“云烟87”“云烟85”下降趋势更快. 42~56 ℃定色期, 烟叶呈现出黄片黄筋的状态, 叶片勾尖卷边, 48 ℃之后叶片及主叶脉含水率下降速率开始降低, 4个品种的叶片含水率都处于20%以下, 而叶脉含水率在56 ℃时才下降到20%, 这个现象也正好对应了56℃之后的干筋期阶段, 烟叶的主叶脉逐步干燥、 变黄, 温度到达68℃时全炉烟叶主脉干燥(图5).
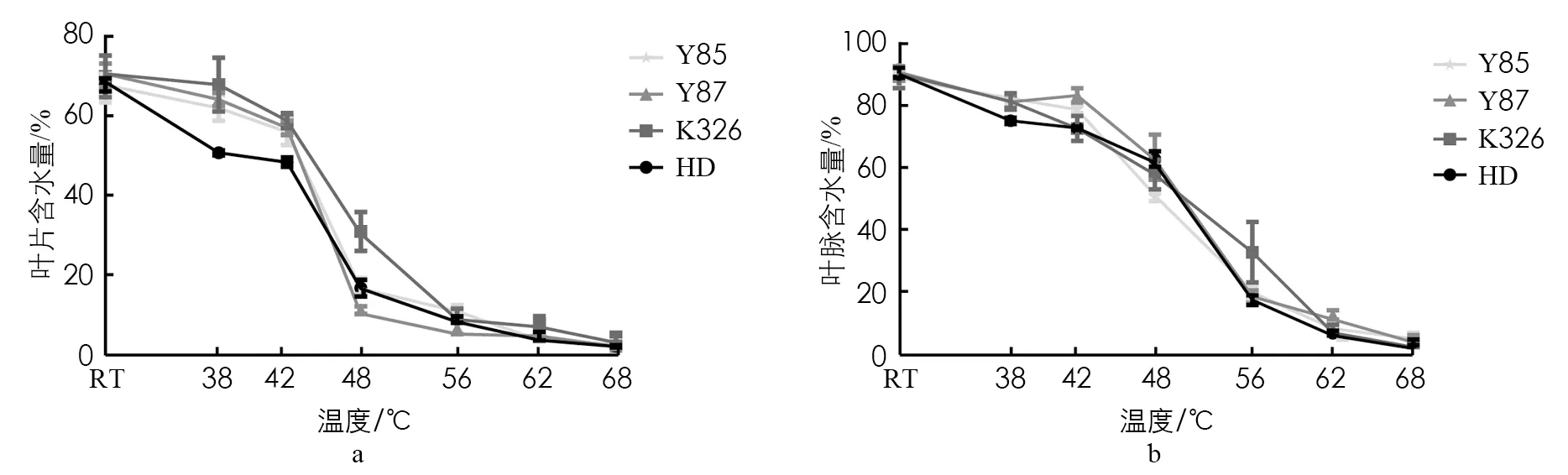
图5 烘烤过程中烟叶的含水量变化
结合先前的叶绿素含量变化来看, 上部烟叶烘烤过程中烟叶失水主要集中在变黄后期至定色期之间, 而“红花大金元”色素含量高, 叶片失水速率过快, 主叶脉失水速率低, 叶片和主叶脉的失水和变黄速度协调性差, 这也是其上部烟叶易烤性差, 烘烤后的烟叶青烟率和杂色烟率较高的原因.
2.5 烘烤过程中烟叶的抗氧化特性变化
在烘烤后期, “红花大金元”“K326”的叶片发生了褐变反应[22], 烟叶整体色泽呈现出黑褐色并且部分烟叶有糊片、 黑糟的现象, 这也是烘烤中常出现的现象. 而多酚氧化酶作为催化酶类, 催化产生酚类物质, 是烟叶转变为棕色并进一步褐变的主要原因[23]. 过氧化物酶是生物体内重要的清除剂, 用于降解酚类、 甲醛等[24]. 超氧化物歧化酶也是抗氧化酶之一, 是生物体内清除自由基和有害物质的重要酶类[25].
图6是上部烟叶烘烤过程中的多酚氧化酶PPO(图6a)、 过氧化物酶POD(图6b)、 超氧化物歧化酶SOD(图6c)活性变化示意图. 从烤前到42 ℃的变黄阶段中, “云烟87”“云烟85”的多酚氧化酶活性显著低于“红花大金元”, 而到了定色前期48~56 ℃, “红花大金元”“K326”的活性有所下降, “云烟87”“云烟85”的多酚氧化酶活性却显著升高. 4个品种鲜烟叶中的过氧化物酶活性无太大差异, 但随着烘烤时间的延续, 过氧化物酶的活性均逐渐增强, 但品种间具有显著差异, 表现为“云烟87”和“云烟85”高于“红花大金元”和“K326”. 烘烤前“云烟87”“云烟85”的超氧化物歧化酶活性相较于“红花大金元”“K326”更高一些, 随着烘烤时间及温度的变化, 酶活性逐渐增强. 4个品种的SOD平均活性表现为“云烟85”最高, “云烟87”次之, “红花大金元”相对于“K326”最低. 中高温胁迫导致烟叶PPO活性升高, 随之增加细胞内自由基和酚类等物质含量, 而这些物质的积累又会引起植物的褐变反应, 不过细胞内抗氧化酶活性也会随温度增强, 清除细胞体内的自由基和酚类等小分子物质[26]. 综合来看, 虽然在定色阶段“云烟87”“云烟85”的PPO活性要高于“K326”“红花大金元”, 但由于其POD与SOD活性远远高于“K326”“红花大金元”, 其细胞清除自由基和酚类等小分子物质的作用更强, 细胞环境变化较为稳定. 而随着温度增加, 细胞膜系统受损可能会进一步增加多酚类物质的释放, 而酶活性均逐渐降低, “红花大金元”“K326”产生的酚类物质更多, 从而产生棕色化反应及酶促褐变, 出现深浅不一的杂色烟、 黑糟等现象.

图6 烘烤过程中烟叶的抗氧化特性变化
3 结论
本试验采用三段六步式烘烤工艺对易烤品种“云烟87”“云烟85”和难烤品种“红花大金元”“K326”的上部烟叶进行烘烤. 选取烘烤前、 变黄期(38 ℃,42 ℃)、 定色期(48 ℃,56 ℃)、 干筋期(62 ℃)及烘烤后(68 ℃)5个时期不同时间点的烟叶样品, 探究了烘烤过程中变黄、 失水特性、 质体色素降解、 碳代谢、 氮代谢及相关酶活性等理化性质的变化规律.
研究表明, 烘烤过程中烟叶的颜色变化与叶片含水率及糖含量等化学成分变化密切相关. 在变黄期间, 烟叶颜色从绿色变为浅黄色, 烟叶叶片含水率由70%左右下降至58%左右, 此阶段叶绿素酶活性显著升高, 叶绿素含量大幅度降低, 还原糖、 总糖含量升高; 定色阶段, 烟叶黄色逐渐加深, 呈现出深橘黄色, 烟叶含水率由 58%左右降至10%左右, 多酚氧化酶及抗氧化酶活性增强.
在烘烤过程中“红花大金元”烟叶变黄速度较慢, 变黄期叶片失水速率又较快, 这与文献[27-28]的研究结果一致. 大分子糖类物质降解不充分导致总糖含量低, 到排湿定色阶段时, 其失水速率有所下降, PPO酶活性升高, 导致酚类物质积累, 而抗氧化的POD和SOD酶活性相对较低, 对烟叶变黄与定色不利, 难于烘烤且定色期易发生褐变, 不耐烤. 烤后烟叶粗蛋白含量大于12%, 黑糟、 糊片杂色烟率大, 品质较差. “K326”烟叶变黄速度稍快, 失水速率适中, 虽然其多酚氧化酶活性偏低, 但其定色期抗氧化酶活性较低, 清除体内自由基及酚类物质的能力较差, 烟叶易发生褐变, 耐烤性较差. 而“云烟87”“云烟85”在变黄期变黄速度适中, 叶片失水速率较快, 变黄与失水特性协调, 大分子糖类物质降解充分, 总糖含量累积多, 在定色期间虽然多酚氧化酶活性高, 但其POD和SOD酶活性相对更高, 清除细胞体内的自由基、 酚类物质的能力更强, 烤后烟叶粗蛋白含量10%左右, 烟碱、 总氮含量相对适中, 烟叶品质较好.
本研究系统分析了重庆烟区不同烘烤难易品种的上部烟叶物质变化规律, 研究结果初步表明烟叶变黄与失水的协调性、 粗蛋白含量及抗氧化酶活性差异是导致不同品种上部烟叶烘烤难易程度不同的主要因素, 研究工作为下一步通过基因工程手段改良烟草烘烤特性奠定了坚实的基础.
