贵州娘娘山湿地两种藓类沼泽植物群落组成及其与环境因子的关系*
2022-10-26杨浚恒崔海军石珣珣郭应罗金祥王舒谭敏
杨浚恒,崔海军,石珣珣,郭应,罗金祥,王舒,谭敏
(1.西南林业大学 地理与生态旅游学院,云南 昆明 650224;2.西南林业大学 云南省高原湿地保护修复与生态服务重点实验室,云南 昆明 650224;3.盘州市自然资源局,贵州 盘州 553537)
泥炭藓(Sphagnumpalustre)沼泽湿地具有强大的涵养水源、净化水质的功能,是多种生物优良的栖息地,同时也是重要的碳储库,对于环境演变和全球气候变化的区域响应研究具有十分重要的价值[1]。世界上泥炭藓沼泽湿地多分布于阿拉斯加、西伯利亚、加拿大、瑞典、新西兰和捷克等温带到副极地地区[2-3]。我国泥炭藓沼泽湿地主要分布于大兴安岭、小兴安岭及长白山等温带地区[4-6],亚热带地区也发现分布有亚高山泥炭藓沼泽斑块,如福建天宝岩和鄂西的七姊妹山、大九湖、二仙岩、太山庙及贵州娘娘山、独山、麻若平台均相继报道了泥炭藓沼泽的分布[2,7-11]。与温带的泥炭藓沼泽相比,亚热带泥炭藓沼泽具有稀有性、珍贵性特点,且具有重要的生态、科研、科普及旅游等价值。然而,泥炭藓沼泽具有脆弱性,且遭受采藓、火烧、泥炭开采、农垦、造林等对其造成的巨大威胁[12-14]。受破坏后的泥炭藓泥炭地的自然恢复需要很长时间,其往往要经历旱生藓类的演替阶段,如金发藓科(Polytrichaceae)植物阶段。长白山哈尼泥炭地和贵州娘娘山泥炭地均报道分布有大面积的泥炭藓沼泽和金发藓(Polytrichumcommune)沼泽[9,15]。
藓类沼泽植物群落往往具有分层现象,除藓类植物层外通常还有上层维管植物草本层或矮灌木层,藓类沼泽植被蕴藏着丰富的植物多样性[16-18]。我国亚热带亚高山藓类沼泽通常为草本型沼泽,藓类植物通常占据优势地位,或与草本维管植物共优生长。不同温度、水文及养分条件形成不同的藓类沼泽植物群落,群落水平多样性保育亦是藓类沼泽生物多样性保育的重要内涵[17,19-20]。因此,探索藓类沼泽植物群落组成与环境因子之间的关系,对于该类型湿地的保护与恢复以及未来气候变化或人为干扰条件下该类型湿地的演变预测均具有重要意义。
环境因子间往往具有显著的相关性,如土壤全碳含量与全氮含量通常呈显著正相关,土壤容重与土壤含水量通常呈显著负相关,土壤酸碱性可影响多种元素离子的存在形式而影响其浓度或有效性[21-22]。科研监测可以利用这一现象,在仅测定若干个重要因子或代表因子的情况下而快速获得环境的主要信息,进而降低监测繁度与成本。排序分析中的前向选择或后项选择可去除环境因子间的共线性而达到此目的[23]。
本研究选择云贵高原典型亚热带藓类泥炭沼泽——泥炭藓沼泽和金发藓沼泽为研究对象,通过植物群落调查与环境因子的测定,利用排序分析探索不同藓类植物群落类型物种组成差异及其与环境因子之间的关系,以期在全球变化条件下为该类型湿地的科学研究与监测提供参考。
1 材料和方法
1.1 研究区域概况
贵州娘娘山湿地位于云贵两省交界处,地理坐标为104°45′24″~104°51′41″E、26°04′25″~26°8′24″N,是云贵高原典型的亚热带喀斯特岩溶山地泥炭沼泽湿地,于2018年正式获批国家湿地公园。藓类沼泽在其生态系统功能的维持和发挥中具有重要作用,藓类沼泽主要类型为泥炭藓沼泽和金发藓沼泽。藓类沼泽发育于喀斯特地区亚高山台地的玄武岩岩层的积水区域,其涵养的大量水资源为周边社区及流域经济社会发展提供了保障。贵州娘娘山湿地气候属典型的亚热带季风气候。气温变化小,历年极端最高气温36.7 ℃,极端最低气温-7.9 ℃,年平均气温为15.2 ℃。年平均日照时数1 615 h,日照百分率37%。年平均降雨量1 413.6 mm,平均蒸发量1 526.7 mm。年平均相对湿度76%,最大相对湿度100%,最小相对湿度2%。年平均无霜期271 d。贵州娘娘山湿地保育区主要土壤类型为泥炭土和灌丛草甸土。
1.2 样地设置
2018年8月下旬在湿地公园保育区分别选择面积较大的泥炭藓沼泽斑块3个和金发藓沼泽斑块2个,每个斑块植被组成均较均匀。在每个斑块中分别设置一个有代表性的、面积为20 m×20 m的样地,在各样地随机设置3个1 m×1 m的样方,因此共设置样地5个,样方15个。5个样地分别命名为NTX1、NTX2、NTX3和JFX1、JFX2。记录每个样方的海拔(Alt)、经纬度、坡度(Gra)、坡向(Asp),调查其植物群落组成,采集0~10 cm泥炭土测定土壤理化性质。
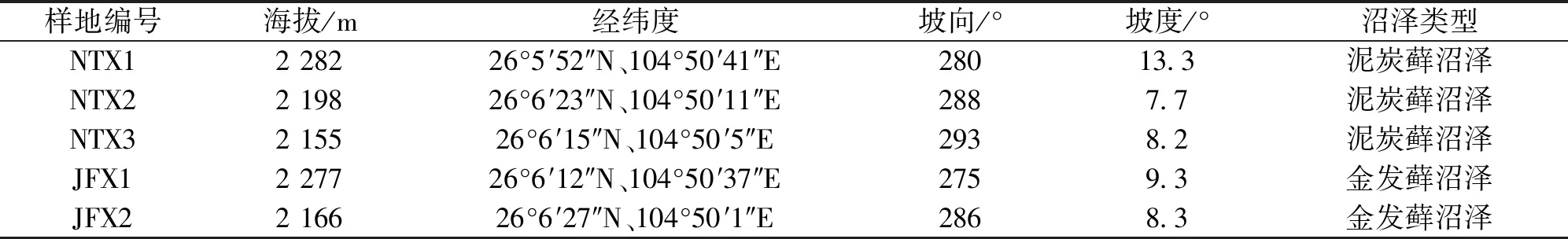
表1 样地基本信息Tab.1 Site basic information
1.3 植物群落调查
用内径为1 m×1 m且等分成25个20 cm×20 cm的小方格的样方框调查植物群落组成。记录每个小方格中出现的物种,并记录是否占优势,计算物种在样方中出现的频度,估计每个物种在样方中的盖度,物种的高度通过测定5株高度求平均值获得。计算每个样方出现物种的相对盖度、相对高度、相对频度,进而计算物种的重要值。
物种重要值=(相对盖度+相对高度+相对频度)/3。
每个样地出现物种的重要值等于物种在3个样方重要值的均值。依据物种在样地中重要值的大小,确定物种的优势程度,进而确定优势种。把每个样地所含物种重要值从大到小排列,把排在前列且重要值相加之和刚刚达80%的物种定义为植物群落主要物种,进行排序分析。
1.4 群落类型的命名
采用优势种命名原则,并按各优势种在群落中所处层次的高低,将各优势种的名称自上而下地排列,同一层次的优势种之间用“+”号联结,不同层次的优势种之间用“-”号联结。
1.5 土壤理化性质测定
植物群落调查完毕后,去除地上绿色植被层及藓类死亡部分和凋落物层,采用随机3点取样法,用泥炭钻取0~10 cm泥炭层测定土壤理化性质。
采用烘干法测定土壤容重(VW)及重量含水量(Wg)。利用风干土,采用玻璃电极(STARTER 300,上海)测定土壤pH值(土水比1︰5,W/V)。土壤全碳含量(TC)采用总有机碳分析仪(Vario,德国)测定。土壤全氮含量(TN)采用H2SO4-H2O2消煮,消煮液采用流动分析仪测定(SKALAR SAN++,荷兰)。土壤铵态氮含量(NH4)及硝态氮含量(NO3)采用1 mol/L KCL浸提法(土水比1︰10,W/V),浸提液使用连续流动分析仪测定。土壤全磷含量(TP)采用HClO4-H2SO4消煮,消煮液采用流动分析仪测定。土壤有效磷含量(AP)采用NH4F-HCL溶液浸提,浸提液使用连续流动分析仪测定。土壤全钾含量(TK)采用HF-HClO4(氢氟酸-高氯酸)溶解,然后用电感耦合等离子体发射光谱仪(inductively coupled plasma-optical emission spectrometer,ICP-OES)测定。土壤速效钾含量(AK)采用中性乙酸铵浸提,然后ICP-OES测定。
1.6 数据分析
以植物群落主要物种的重要值作为物种数据,采用主成分分析(PCA)对不同样地群落进行非约束排序,排序结果通过样地样方的相对位置展示植物群落组成的差异。对样地土壤理化性质指标进行方差分析,多重比较采用Duncan法(α=0.05)。以土壤特征指标(pH、TC、TN、 TP、TK、AK、AP、NO3、NH4、Wg、VW)和地形因子(Alt、Gra、Asp)作为环境数据,同时使用物种数据和环境数据对样地进行约束性排序冗余分析(RDA),结果通过样地、物种和环境因子的相对位置展示植物群落组成差异与环境因子的关系。环境因子与植物群落组成相关的显著性通过蒙特卡罗置换检验来实现(通过R软件中的permutest函数实现)。对土壤因子进行Pearson相关分析,且进行显著性检验。采用排序中的前向选择分析去除环境因子中的共线性问题(通过函数forward.sel实现),进而保留显著且有代表性的环境因子。方差分析和相关分析在SPSS 19.0中实现,排序分析主要通过R软件中Vegan软件包实现。
2 结果与分析
2.1 植物群落组成与命名
根据植物群落组成物种的重要值排序结果见表2。

表2 样地物种重要值及群落命名Tab.2 Species importance values and community naming in sites
NTX1、NTX2、NTX3、JFX1、JFX2样地植被可分别命名为条穗薹草-泥炭藓群落(CommunityCarexnemostachys-Sphagnumpalustre)、箭竹+柳叶箬-泥炭藓群落(CommunityFargesiaspathacea+Isachneglobosa-Sphagnumpalustre)、柳叶箬-泥炭藓群落(CommunityIsachneglobosa-Sphagnumpalustre)、毛轴蕨-金发藓群落(CommunityPteridiumrevolutum-Polytrichumcommune)、毛轴蕨-金发藓群落。两种藓类沼泽植物群落均由上层的草本层或草本小灌木层和下层的藓类植物层构成。5种植物群落下层藓类盖度均达100%,且除JFX1样地植物群落藓类植物层既包括金发藓又出现少许泥炭藓外,其他样地植物群落下层藓类植物层均由1种藓类植物构成。
条穗薹草-泥炭藓群落中主要物种按照重要值降序依次包括泥炭藓(Sphagnumpalustre)、条穗苔草(Carexnemostachys)、线叶水芹(Oenanthelinearis)、薄叶新耳草(Neanotishirsuta)、柳叶箬(Isachneglobosa)。箭竹+柳叶箬-泥炭藓群落主要物种依次包括箭竹(Fargesiaspathacea)、柳叶箬、泥炭藓、丝叶薹草(C.capilliformis)。柳叶箬-泥炭藓群落主要物种依次包括泥炭藓、柳叶箬、笄石菖(Juncusprismatocarpus)、薄叶新耳草、箭竹、百球藨草(Scirpusrosthornii)、藏黄花茅(Anthoxanthumhookeri)。毛轴蕨-金发藓群落JFX1样地主要物种依次包括毛轴蕨(Pteridiumrevolutum)、金发藓(Polytrichumcommune)、红粉白珠(Gaultheriahookeri)、菝葜(Smilaxchina)。毛轴蕨-金发藓群落JFX2样地毛轴蕨、金发藓、藏黄花茅、丝叶薹草、四脉金茅(Eulaliaquadrinervis)。
基于植物群落主要物种重要值的主成分分析结果表明,两个主成分轴PC1、PC2共解释了植物群落差异总方差的73.71%,其中PC1解释了56.55%,PC2解释了17.16%,说明保留PC1和PC2的主成分分析可以反映样地间植物群落主要物种组成的总体差异(图1)。由排序图可以看出,泥炭藓沼泽植物群落与金发藓沼泽植物群落具有明显分异,泥炭藓沼泽植物群落分布于PC1<0的一侧,金发藓沼泽植物群落分布于PC1>0的一侧。泥炭藓沼泽植物群落间也具有明显分异,条穗薹草-泥炭藓群落(NTX1)分布于PC2<0的一侧,箭竹+柳叶箬-泥炭藓群落(NTX2)和柳叶箬-泥炭藓群落(NTX3)分布于PC2>0的一侧。金发藓沼泽植物群落间(JFX1和JFX2)的分异较小,箭竹+柳叶箬-泥炭藓群落(NTX2)和柳叶箬-泥炭藓群落(NTX3)间分异也较小。从排序图还可以看出各样地内的样方点均分布比较集中,说明各样地内植物群落主要物种组成的异质性较小(图1)。主要物种中,泥炭藓、毛轴蕨和金发藓在PC1上的因子载荷较大(|PC1|>0.5),说明这些物种主要影响泥炭藓沼泽植物群落和金发藓沼泽植物群落间的分异;主要物种中条穗薹草和箭竹在PC2上的因子载荷较大(|PC2|>0.5),说明这两个物种主要影响泥炭藓沼泽植物群落间的分异(图1,表3)。
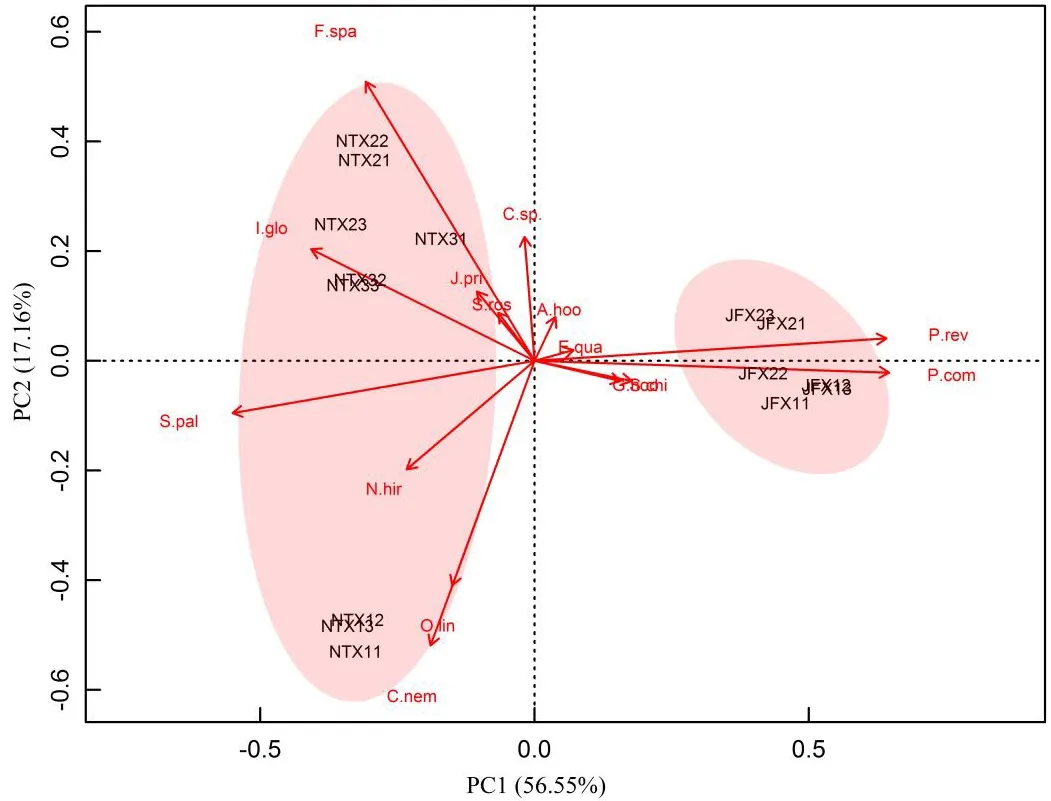
图1 基于主要物种重要值的植物群落组成的主成分分析注:NTX11、NTX12、NTX13分别为样地NTX1样地的重复样方,其他样方名称以此类推;物种拉丁缩写名同表3。Fig.1 Principal component analysis(PCA)of plant community coposition based on major species importance value
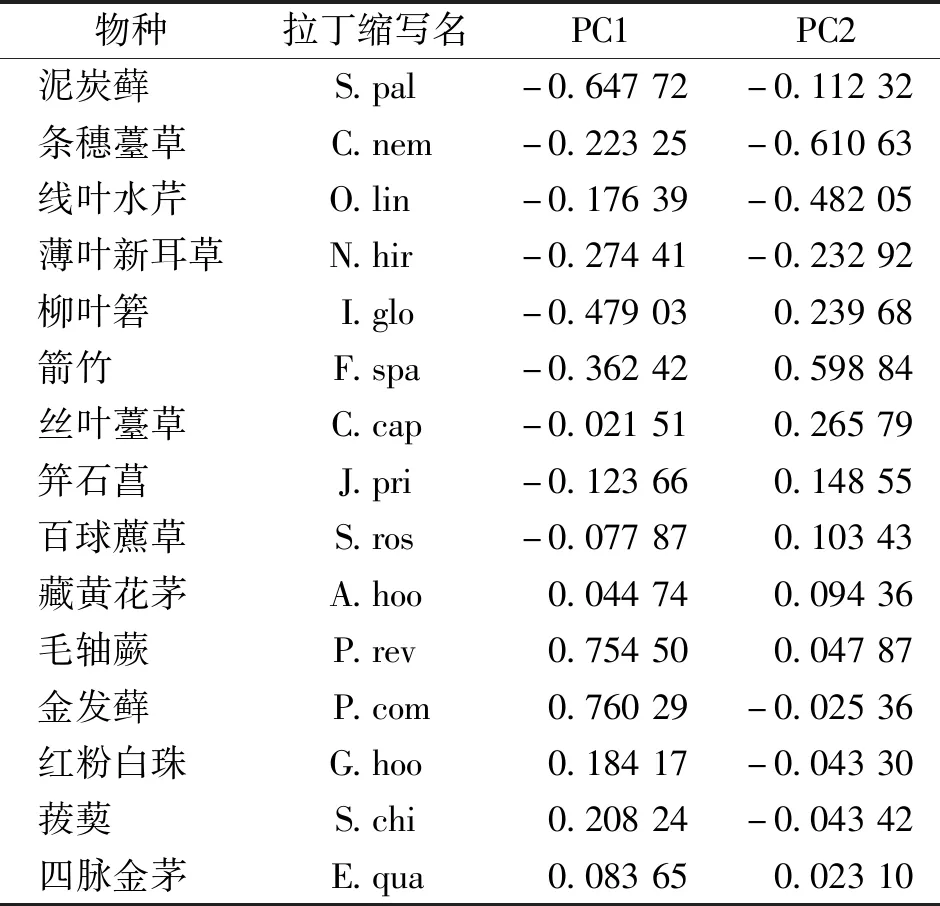
表3 主成分分析中PC1、PC2的植物群落主要物种载荷因子Tab.3 Plant community major species loading factors of PC1 and PC2 in PCA
2.2 土壤因子在样地间的差异
土壤因子速效钾含量、铵态氮含量、pH、全钾含量、重量含水量在泥炭藓沼泽样地的均值均大于或等于在金发藓沼泽样地的均值;土壤有效磷含量、全氮含量和容重在泥炭藓沼泽样地的均值均小于在金发藓沼泽样地的均值(表4)。其中,泥炭藓沼泽土壤pH值均显著高于金发藓沼泽的值。在泥炭藓沼泽中,泥炭藓样地NTX2的速效钾含量、硝态氮含量、全碳含量均值均显著高于泥炭藓样地NTX1与NTX3的值。
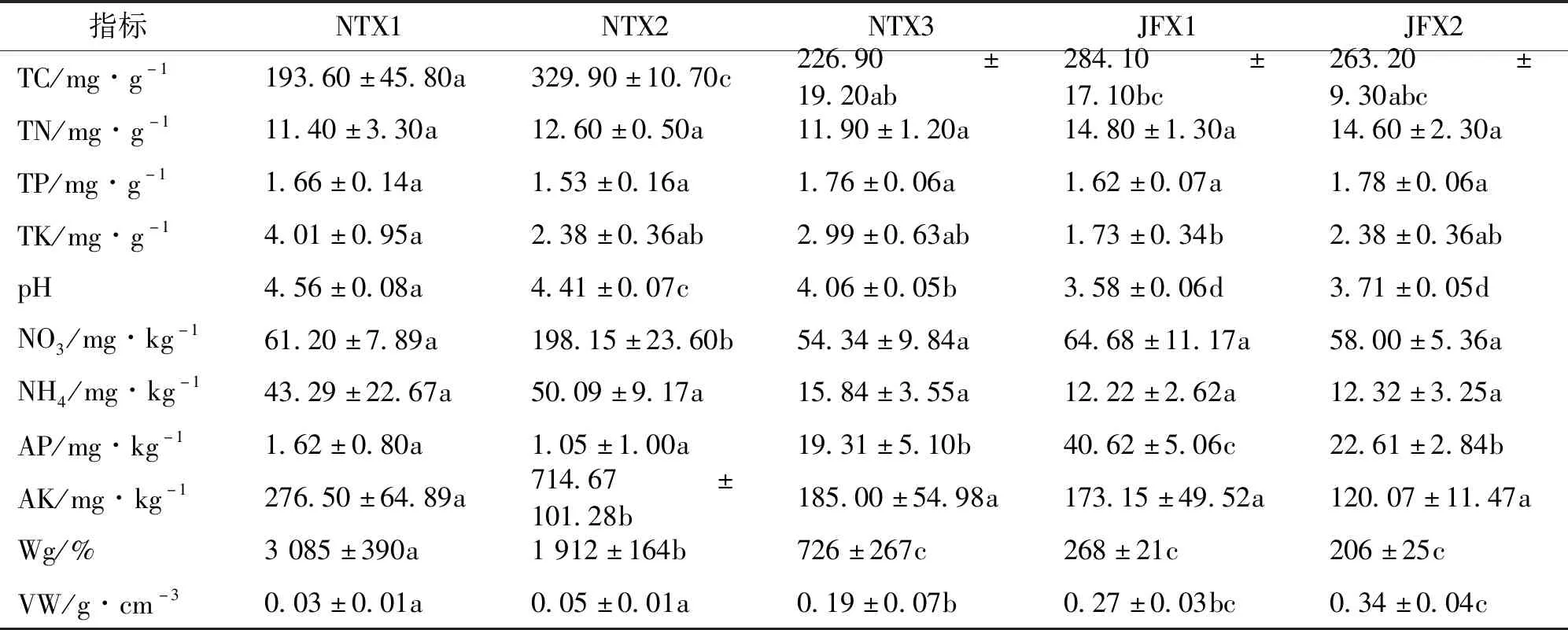
表4 土壤因子在样地间的差异Tab.4 Differences in soil factors between sites
2.3 植物群落主要物种组成与环境因子之间的关系
约束性排序RDA分析结果表明,环境因子对植物群落主要物种组成差异的方差解释度达100%,RDA1和RDA2两个轴共解释总变异方差的73.71%(图2),因此保留前两个排序轴对于分析植物群落主要物种组成与环境因子之间的关系具有一定的代表性。
RDA1轴主要解释泥炭藓沼泽类型和金发藓沼泽类型间的植物群落主要物种组成差异;RDA2轴主要解释泥炭藓沼泽类型内部植物群落主要物种组成差异(图2)。与RDA1轴相关性高的环境因子包括全氮含量、全磷含量、pH、铵态氮含量、有效磷含量、重量含水量、容重(|r|>0.8),说明这些因子主要与泥炭藓沼泽类型和金发藓沼泽类型间植物群落主要物种组成差异相关;与RDA2轴相关性高的环境因子包括全碳含量、硝态氮含量、海拔、坡度,说明这些因子主要与泥炭藓沼泽类型内部植物群落主要物种组成差异相关(表5)。对植物群落主要物种组成的解释总体达到显著水平的因子包括pH、硝态氮含量、有效磷含量、速效钾含量、重量含水量、容重、海拔(P<0.05),其中达极显著水平的因子包括pH、有效磷含量、重量含水量、容重(P<0.01)(表5)。
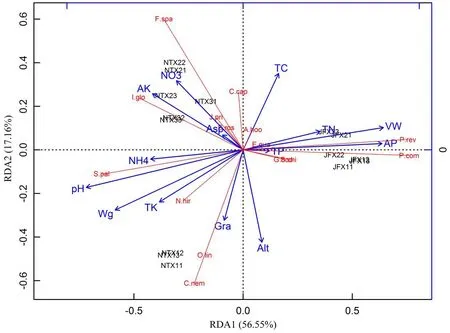
图2 以环境因子(土壤和地形)作为解释变量的植物群落主要物种组成的冗余分析排序图Fig.2 Redundancy analysis(RDA)ordination diagrams of the plant community major species compostion with environment factors(soil and topography factoes) as explained varibles
RDA排序图中可以通过解释变量(环境因子)和响应变量(物种)之间的夹角,判断它们之间的相关性。对植物群落主要物种组成解释达极显著水平的pH、重量含水量分别与主要种泥炭藓重要值呈高度正相关,分别与金发藓和毛轴蕨重要值呈高度负相关,有效磷含量和容重则分别与这三物种之间的相关关系相反(图2)。硝态氮含量与箭竹重要值呈高度正相关,速效钾含量与柳叶箬重要值呈高度正相关。条穗薹草重要值与海拔正相关,与土壤全碳含量高度负相关(图2)。
多个土壤因子间存在共线性关系,pH与全钾含量、铵态氮含量、重量含水量间存在显著或极显著正相关;pH与全氮含量、有效磷含量、容重间存在显著或极显著负相关(图2,表6)。硝态氮含量与全碳含量、铵态氮含量、速效钾含量间存在显著或极显著正相关。容重与重量含水量呈极显著负相关。
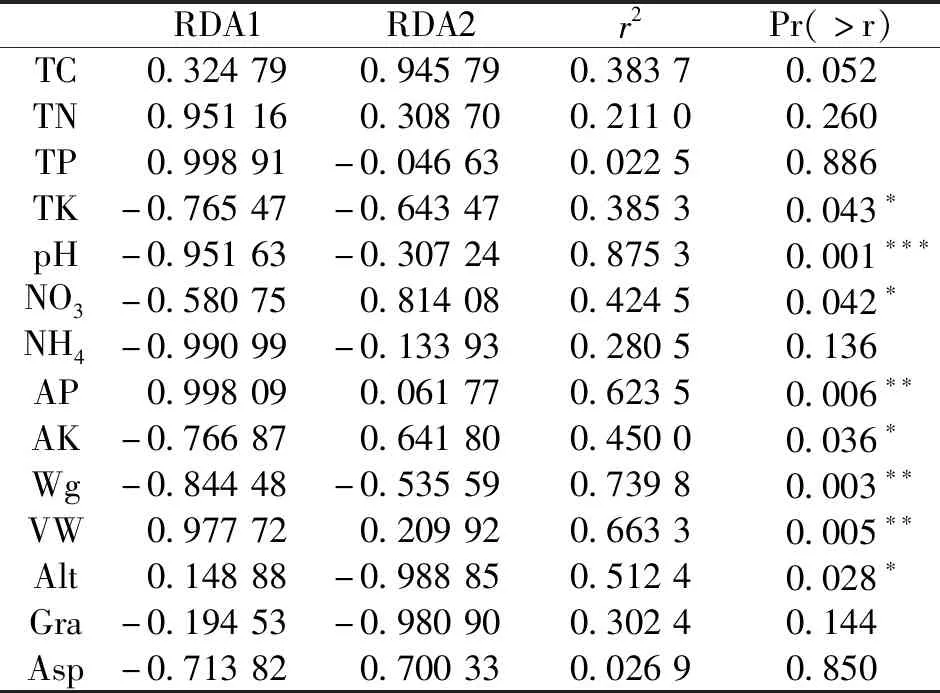
表5 环境因子与排序轴的相关系数及其对植物群落主要物种组成影响的显著性检验Tab.5 Correlation coefficients between environmental factors and the ordination axes,and significance testing to the effect of environmental factors on plant community major species composition
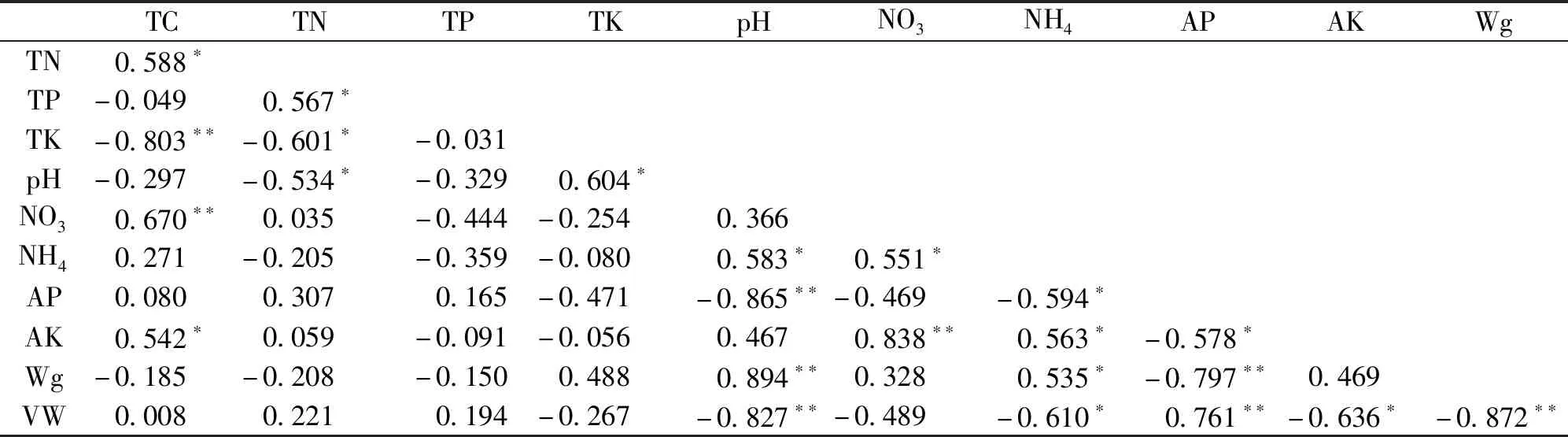
表6 土壤因子相关性分析Tab.6 Correlation analysis between soil variables
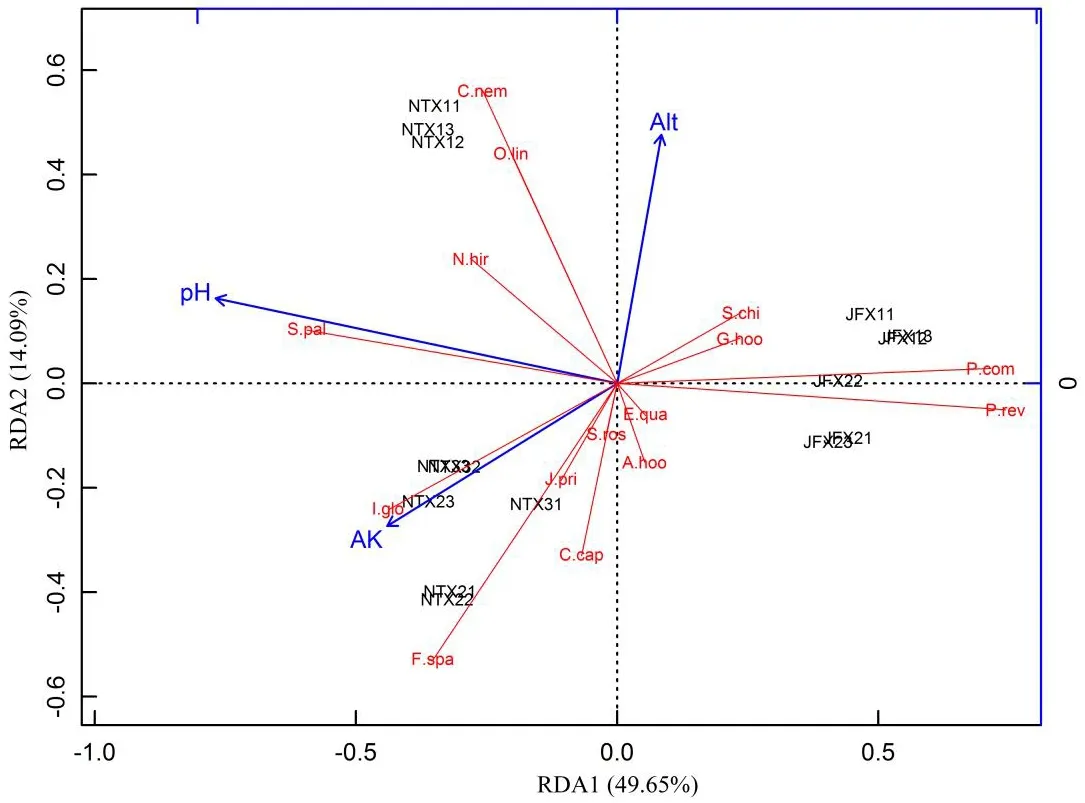
图3 环境因子前向选择后的植物群落主要物种组成的冗余分析(RDA)Fig.3 Redundancy analysis(RDA)of plant community majorspecies composition after forward selection of environmental factors
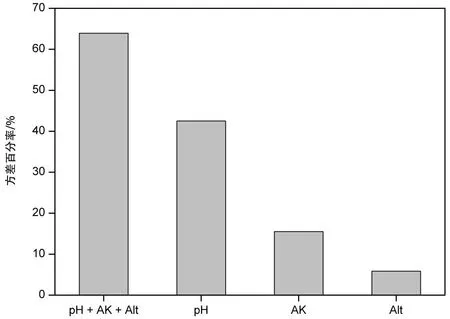
图4 经前向选择后保留的环境因子对植物群落分异方差的解释量Fig.4 The explained variance percentage for plant community differentiation of environmnetal factors retained after forward selection
共线性可能会造成解释方差膨胀,使环境因子对总方差的解释度高于真实值,前向选择可解决共线性问题。经前向选择分析后,保留了3个环境因子,分别为土壤pH、速效钾含量、海拔,RDA1和RDA2解释了总方差的64.55%(图3),与全部环境因子RDA前两轴的解释度73.71%接近(图2)。土壤pH、速效钾含量、海拔对总方差的校正解释度共为63.93%,分别为42.52%、15.50%、5.86%(图4)。因此,可以判断土壤pH与植物群落主要物种组成紧密相关。
3 讨论与结论
3.1 两种藓类沼泽植物群落组成
本研究调查了贵州娘娘山国家湿地公园保育区内较大斑块藓类沼泽植物群落类型,所记录的3种泥炭藓群落类型均为草本型沼泽,在鄂西和福建天宝岩均发现有类似的泥炭藓群落类型。鄂西发现的大理苔草-泥炭藓群落、川东薹草-泥炭藓群落与本研究命名的条穗薹草-泥炭藓群落相类似,优势维管植物均为莎草科植物;鄂西和福建天宝岩均为水竹-泥炭藓群落与本研究命名的箭竹+柳叶箬-泥炭藓群落相类似,优势维管植物均存在竹亚科植物;鄂西的紫羊茅-泥炭藓群落与本研究命名的柳叶诺-泥炭藓群落相类似,优势维管植物均为禾本科植物[2,24]。鄂西属于中亚热带季风湿润气候,天宝岩属于中亚热带海洋性季风气候,本研究区域属于典型亚热带季风气候[11,24]。不同区域及气候类型分布有相似植物群落类型,也间接说明藓类泥炭沼泽作为一种重要的湿地类型具有隐域植被的特征[21]。
本研究区分布着大面积的金发藓沼泽,而鄂西和福建天宝岩未见有该类型藓类沼泽大面积分布的报道,只在报道的物种名录中有金发藓科植物的存在[2]。这可能与本研究区域20世纪90年代末大面积商业采集泥炭藓及排水造林有关系。人为干扰后泥炭沼泽将发生次生演替,裸露的泥炭层自然恢复到泥炭藓群落要经历漫长的金发藓群落阶段。卜兆君等[12]对长白山哈尼泥炭藓沼泽研究发现,采藓等人为干扰后金发藓快速占领裸露的泥炭地,可稳定占领数十年后才会演替到泥炭藓群落,体现出泥炭藓沼泽生态系统的脆弱性。在自然恢复过程中,金发藓群落可能为泥炭藓群落的恢复创造一定的条件,可能通过改善泥炭地的水文条件及基质的稳定性,而有利于泥炭藓孢子的萌发与克隆生长[25]。本研究中JFX1样地金发藓沼泽植物群落藓层中出现少许的泥炭藓可能正是上述现象的反映(表2)。
3.1.2 植物群落组成与环境因子之间关系
本研究发现土壤pH、重量含水量、容重、有效磷含量对藓类沼泽植物群落的分异具有极显著影响,速效钾含量、硝态氮含量及海拔具有显著影响。陈旭等[4]研究发现水位埋深、沼泽水pH以及乔木郁闭度和灌木盖度是影响泥炭地苔藓植物生态位分异的主要环境因子,泥炭全磷含量对苔藓植物分布影响较大。郭水良等[26]研究发现金发藓科植物在海拔、土壤含砂量、含水量、酸度、光照条件等资源维度上存在明显的生态分化现象。以上研究均支持土壤pH、含水量为影响藓类沼泽植物群落分异的重要环境因子。
水位埋深或土壤含水量不同反映苔藓耐旱能力差异,而耐旱能力不同是导致苔藓植被演替的原因之一。在泥炭地环境趋于变干的条件下,耐旱能力强的苔藓会逐渐取代耐旱能力差的苔藓。金发藓属植物的地下根茎系统具有很高的能量利用效率,植物体有良好的水传导组织和复杂的叶片结构,能够在其他苔藓无法进行光合作用的干旱条件下生长[27]。金发藓属植物常见于人为扰动和植被原生演替的早期阶段的生境中,表现出较强的耐环境胁迫的能力[28-29]。Clymo[30]研究发现沼泽水pH主要与苔藓离子交换能力有关,耐旱性苔藓由于脱离地下水位,需要有更强的阳离子交换能力以获取足够的养分,苔藓植物个体阳离子交换能力的差异导致生境中pH不同。本研究中泥炭藓沼泽土壤pH显著高于金发藓沼泽土壤pH,这是否为2种藓类植物离子交换能力的差异所致,尚待今后进一步深入研究验证。
本研究发现多个土壤因子间存在共线性关系。陈旭等[4]研究发现TN与TP之间存在显著正相关,与本研究的结果一致。pH通过影响元素主要存在形式,从而间接影响它们之间的相关性[21]。本研究发现pH与有效磷成极显著负相关,这可能是因为低的pH条件下H+浓度高,利于形成磷酸根,进而有效磷的浓度较高。对北方雨养型和矿养型泥炭沼泽的研究揭示泥炭地初级生产力和微生物生长普遍存在磷限制,尤其在泥炭藓出现的泥炭地更为明显[31-32]。本研究金发藓沼泽样地有效磷含量均值均高于泥炭藓沼泽样地,且大多达显著水平,进一步说明泥炭藓沼泽植物群落具有更强的磷限制。本研究发现pH与铵态氮含量成显著正相关,这可能与泥炭藓沼泽土壤溶液中矿化氮(氨)的含量较高有关。Kaštovská等[32]认为泥炭藓泥炭中相对较高的氮可用性可能与泥炭藓的某些特定特性有关:这可能与泥炭藓相关的蓝细菌和甲烷氧化菌对大气中N2的生物固定作用及衰老泥炭藓对氮的再吸收效率低有关[33-34],这也可能是土壤重量含水量与铵态氮含量呈显著正相关的间接因素。
本研究结果显示泥炭藓沼泽植物群落间存在明显分异,全碳含量、硝态氮含量在其分异中起重要作用。维管植物可以通过生理、养分循环特性及凋落物质量而直接影响泥炭的养分循环及碳储存。硝态氮含量与箭竹重要值呈高度正相关,这与箭竹的生理结构特性有关,箭竹中空的地上茎及地下茎有利于氧气往土壤的输送,箭竹根际高浓度的氧有利于增强土壤硝化作用,进而硝态氮含量增加是硝态氮含量对泥炭藓沼泽植物群落间分异起重要作用的原因。维管植物可以通过根系分泌物释放活性碳塑造土壤微生物群落结构,维管植物的存在可能影响微生物生物量及其活性[35-37],进而间接地影响泥炭性质及碳储存功能。Kaštovská等[32]研究发现维管植物的存在对泥炭养分循环和碳储存的影响具有物种特异性。在本研究中,土壤全碳含量在泥炭藓群落类型分异中的作用可能与维管植物对系统碳储存功能影响的物种特异性有关。
3.2 结论
金发藓沼泽可以看作泥炭藓沼泽受采藓或排水等干扰后的次生演替阶段,其可为泥炭藓的恢复创造有利条件。土壤水分、pH、有效磷含量与两种藓类沼泽植物群落组成分异紧密相关,是否可通过改善土壤水分条件与磷的可利用性以促进金发藓沼泽向泥炭藓沼泽的演替进程,进而加快泥炭藓沼泽恢复,有待今后进行深入研究。维管植物在泥炭藓沼泽土壤碳储存和养分循环中具有重要作用,且具有物种特异性, 研究气候变化或人类干扰条件下泥炭藓沼泽维管植物的变化对于深入认识该类型湿地生态系统功能的改变具有重要意义。
