持续性内切酶EG5C-1的改造及酶学性质
2022-10-24吕科旻周雨朦何冰芳
吕科旻,周雨朦,吴 斌,何冰芳
(1. 南京工业大学 生物与制药工程学院,江苏 南京 211800;2. 南京正大天晴制药有限公司,江苏 南京 222062;3. 南京工业大学 药学院,江苏 南京 211800 )
纤维素是地球上含量最丰富的可再生资源,因其具有替代生物燃料和化学工业中化石燃料的潜力,受到人们的广泛关注[1]。由于天然纤维素的结构和组织十分复杂坚固,通常需要至少3种酶的协同作用,包括内切葡聚糖酶(EC 3.2.1.4),随机地水解纤维素的非结晶部分并切断β-1,4-糖苷键,产生不同聚合度的纤维寡糖和新的游离末端;外切葡聚糖酶(EC 3.2.1.91),作用于纤维素链的还原端和非还原端并不断降解产生纤维二糖产物;β-葡萄糖苷酶(EC 3.2.1.21),将纤维二糖和寡糖最终水解为可溶性葡萄糖[2]。因此,酶混合物的高成本和低水解效率一直是纤维素转化领域的主要障碍[3]。
近年来,人们不断地从纤维素降解菌中发现具有双催化功能的纤维素酶,这类酶被称为持续性内切葡聚糖酶,因为这类酶同时具有内切酶和外切酶活性,与β-葡萄糖苷酶的相互协同作用可以更加高效地水解纤维素[4]。迄今为止,文献报道的持续性内切葡聚糖酶主要属于GH5和GH9家族的糖苷水解酶[4]。GH9家族的持续性内切葡聚糖酶主要来自梭菌属,如热线梭菌(Clostridiumthermocellum)[5]、解纤维梭菌(C.cellulolyticum)[6]、植物梭状芽孢杆菌(C.phytofermentans)[7]等,该家族的酶通常由催化结构域(GH)和一个或者多个碳水化合物结合模块(CBM)组成。而GH5家族的持续性内切葡聚糖酶有来源广泛及组成结构多样的特点,如:来源于草菇(Volvariellavolvacea)的纤维素酶EG1在C端含有1个GH5催化结构域和在N端还包含1个CBM1的碳水化合物结合模块,并且CBM1的缺失直接影响酶的持续性[8];而来源于Saccharophagusdegradans2-40的持续性内切酶CeI5H,其碳水化合物结合模块CBM6的缺失并不影响酶原有的催化特性[4]。虽然持续性内切葡聚糖酶广泛受到人们的关注,但是目前许多研究主要集中于不同碳水化合物结合模块对于酶持续性的机制方面,而在提高持续性内切葡聚糖酶的催化活性方面的研究报道很少[9]。
笔者所在课题组在前期的研究中从BacillussubtilisBS-5中筛选得到一个持续性内切葡聚糖酶EG5C-1,因此拟通过建立该酶的同源模型和使用分子对接的方法,基于酶活性架构的分析筛选得到23个关键氨基酸残基,利用丙氨酸扫描、饱和突变及组合突变等方法筛选酶催化特性最优的突变体并对其酶学性质和水解产物进行研究分析,为持续性内切酶催化活性的进一步提高提供基础。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
重组菌EG5C-1、大肠杆菌E.coliBL21(DE3),均保存于何冰芳教授实验室。
1.1.2 主要试剂
NaCl、胰蛋白胨、酵母粉、琼脂,生工生物工程(上海)股份有限公司;质粒提取试剂盒,Axygen公司;Phanta高保真酶、DNA marker、DpnⅠ、蛋白Marker,TaKaRa公司;纤维寡糖标品,Megazyme公司。
1.1.3 培养基
LB培养基(g/L):NaCl 10.0、酵母粉 5.0、胰蛋白胨 10.0。固体培养基中另加琼脂 20.0。
LB卡那固体培养基(g/L):NaCl 10.0、酵母粉 5.0、胰蛋白胨 10.0、琼脂 20.0。灭菌冷却后加入过滤无菌的硫酸卡那霉素(Kan),使其终质量浓度为50 mg/mL。
1.2 方法
1.2.1 同源建模与突变点的选择
以来源于B.subtilis168的纤维素酶BsCel5A催化模块的晶体结构(PDB:3PZT)为模板,使用BIOVIA公司的Discovery Studio(DS 3.5)软件构建原始酶EG5C-1的三维结构[10-11]。从来源于短小芽孢杆菌的内切葡聚糖酶GH48的晶体结构(PDB:5CVY)中提取纤维六糖结构,并使用Chemical Computing Group (CCG)公司的MOE2019.01软件进行分子对接,将其对接至原始酶EG5C-1的活性中心架构中。根据对接的自由结合能,选择纤维六糖周围0.5 nm以内的氨基酸作为丙氨酸突变的位点[12]。
1.2.2 突变体文库的构建
使用Overlap PCR方法[13]进行定点突变。根据前期实验获得的持续性内切葡聚糖酶EG5C-1的基因序列和目的突变位点设计相应的引物。以原始酶EG5C-1质粒为模板,Phanta高保真酶进行全质粒扩增,反应条件:95 ℃反应3 min使DNA预变性;95 ℃变性反应15 s;60 ℃退火15 s;72 ℃延伸反应8 min,30个循环,最后72 ℃延伸10 min。扩增产物使用DpnⅠ酶,于37 ℃恒温消化30 min,经0.8%的琼脂糖凝胶验证后导入感受态细胞E.coliBL21(DE3)中。挑取平板的单菌落并送至南京擎科生物科技公司进行序列比对,将正确突变的单菌落接种于终质量浓度为150 g/L的甘油中,于-80 ℃超低温冰箱保藏。
1.2.3 突变体酶的表达与纯化
将突变体接种于含Kan抗性的LB种子培养基中,37 ℃、180 r/min摇床培养12 h。以2%(体积分数)的接种量将种子培养基接种于LB发酵培养基中,37 ℃、180 r/min摇床培养至OD600为0.6左右,加入IPTG使其终浓度为1 mmol/L,25 ℃、180 r/min摇床诱导表达培养24 h。将发酵液离心,收集菌体并用适当的Na2HPO4-KH2PO4缓冲(pH 6.5)重悬,超声破碎菌体,4 ℃、12 000 r/min离心20 min,收集上清液即为粗酶液。粗酶液使用HisTrap FF色谱柱(GE Healthcare)通过亲和色谱法进行纯化,纯化方法见参照文献[14],取纯化酶液进行SDS-PAGE电泳分析,并测定酶活。
1.2.4 蛋白质浓度测定
取适当稀释的纯化酶液,使用Bradfoard法[15]测定蛋白质浓度,以牛血清白蛋白(BSA)为标准。
1.2.5 酶活测定
酶活力单位定义:一个酶活力单位即为在最适反应条件下,每分钟由底物产1 mmol还原糖所需的酶量。
1)羧甲基纤维素钠(CMC-Na)酶活。取0.5 mL适当稀释的酶液,与1.5 mL的CMC-Na(10 g/L)底物于60 ℃条件下反应10 min,立即加入3 mL的3,5-二硝基水杨酸(DNS)溶液终止反应,沸水浴5 min后,使用紫外分光光度计在540 nm处测定吸光度值A450,计算还原糖的生成量。
2)微晶纤维素(Avicel)酶活。取0.5 mL适当稀释的酶液,与1.5 mL的Avicel(100 g/L)底物于60 ℃条件下反应30 min,立即置于冰上冷却,12 000 r/min离心取上清液,加入3 mL的DNS溶液,沸水浴5 min后,使用紫外分光光度计在540 nm处测定吸光度值A450,计算还原糖的生成量。
1.2.6 突变体酶学性质分析
1)最适温度和温度稳定性。取0.5 mL适当稀释的酶液,与1.5 mL的CMC-Na(10 g/L) 底物在30、40、50、60、70、80和90 ℃条件下反应10 min后测定其相应的酶活。以最高的酶活为对照,依次计算相对酶活,分析温度对酶活的影响。将适当稀释的酶液在30、40、50、60、70、80和90 ℃条件下孵育2 h后,测定其相应的酶活。以最高的酶活为对照,依次计算相对酶活,分析温度对酶稳定性的影响。
2)最适pH和pH稳定性。用柠檬酸-柠檬酸钠(pH 3.0~6.0)、Na2HPO4-KH2PO4(pH 6.0~8.0)、Gly-NaOH(pH 8.0~9.0)配制10 g/L不同pH(3.0、4.0、5.0、6.0、7.0、8.0、9.0)的羧甲基纤维素钠,吸取1.5 mL作为底物,加入0.5 mL适当稀释的酶液,于60 ℃条件下反应10 min后,测定其相应的酶活。以最高的酶活为对照,依次计算相对酶活,分析pH对酶活的影响。将适当稀释的酶液分别在pH 3.0、4.0、5.0、6.0、7.0、8.0、9.0条件下孵育2 h后,测定其相应的酶活。以最高的酶活为对照,依次计算相对酶活,分析pH对酶稳定性的影响。
1.2.7 突变体酶水解产物分析
选择CMC-Na、Avicel、磷酸处理的微晶纤维素(PASC)为底物,突变体酶与底物于45 ℃下混合反应6 h,将酶反应混合物沸水浴加热10 min以终止反应,12 000 r/min离心10 min,收集上清液,使用薄层层析法(TLC)[14]分析酶水解产物。
2 结果与讨论
2.1 突变位点的确定
将EG5C-1的同源模型与底物配体分子进行分子对接分析,结果如图1所示。由图1可知:EG5C-1的活性架构是由8个平行的β链和8个α螺旋形成的一个开放式裂隙,该裂隙长约3.1 nm,可以容纳6个葡萄糖基单元。通常情况下,底物结合亚位点从-3到+3分布,其中-n端为非还原段,+n端为还原端,而产物的裂解发生在-1和+1位之间[12]。使用自动对接程序AutoDock 4.2将纤维六糖对接至活性位点结构中,分析酶与底物间起关键作用的氨基酸,对接结果如图2所示。由图2可知:共有23个氨基酸残基,包括H36、W40、Y67、D70、H102、L104、N105、E140、T177、W178、Q180、D181、Y202、T205、H206、F209、K213、E228、A234、S235、W262、K267和E269与对接的纤维六糖分子距离在0.5 nm以内,其中,E140位于β链Ⅳ上作为质子供体,E228位于β链Ⅶ上作为催化亲核基团,由于这2个氨基酸处于EG5C-1的活性中心,将它们突变会直接导致酶完全丧失活性。因此,选择其余的20个氨基酸残基(A234除外)作为突变研究的目标氨基酸。
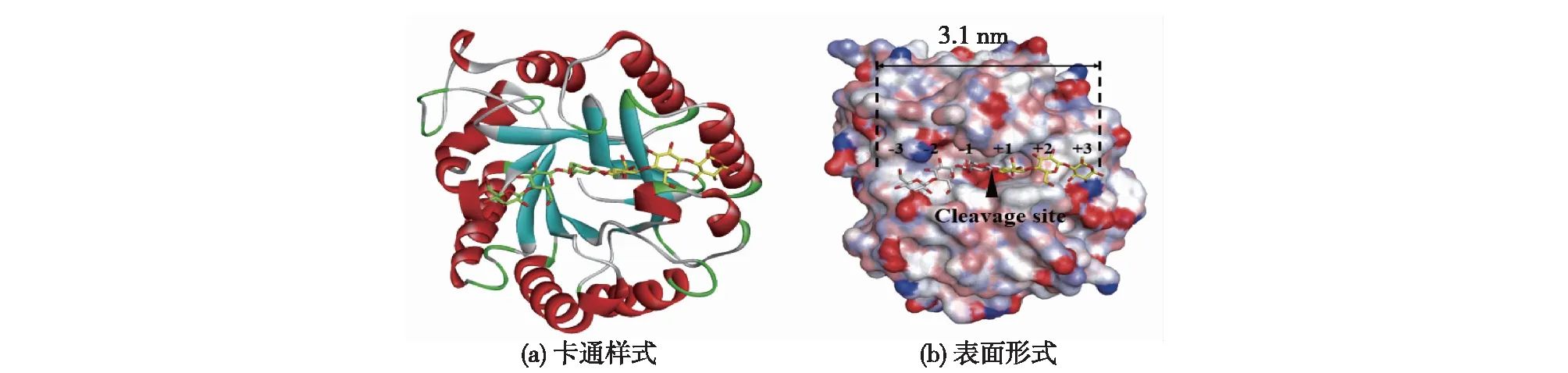
图1 EG5C-1与纤维六糖活性中心对接的卡通样式和表面形式Fig.1 Structures of the active site of EG5C-1 complexed with a cellohexaose molecular carton representation and surface representation
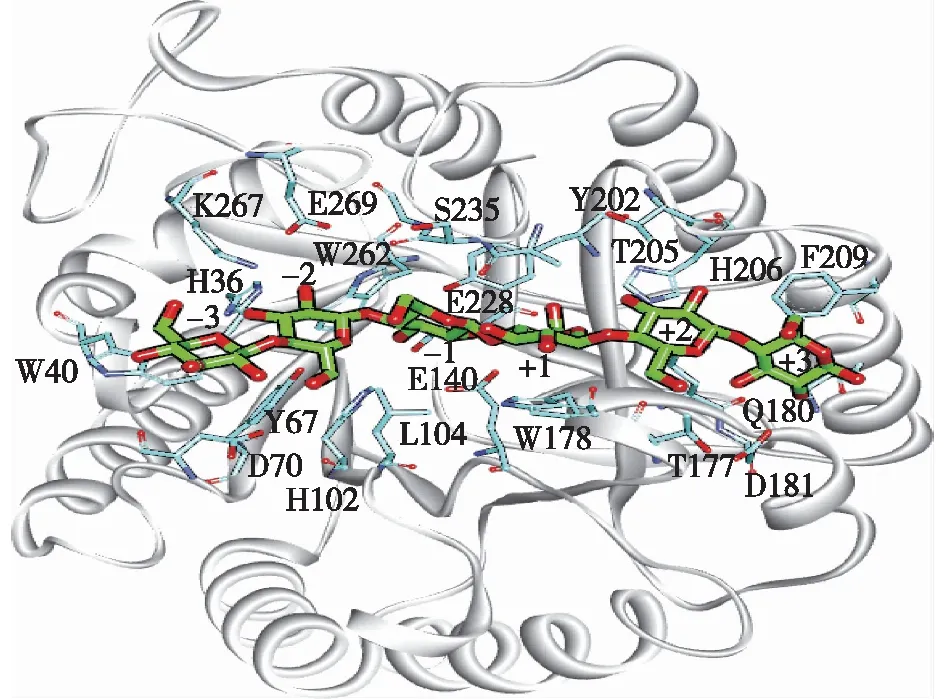
图2 纤维六糖分子对接至持续性内切纤维素酶的催化裂隙中Fig.2 Modeled structure of the processive endoglucanase EG5C-1 with a docked cellohexaose bound to the catalytic cleft
2.2 丙氨酸突变结果
以原始酶EG5C-1质粒pET-28a/EG5C-1为模板,通过PCR扩增,完成相应的突变H36A、W40A、Y67A、H102A、L104A、N105A、T177A、W178A、Q180A、D181A、Y202A、T205A、H206A、F209A、K213A、S235A、W262A、K267A和E269A(A234除外)。通过测序和序列比对确定突变位点与设计的目标位点一致。
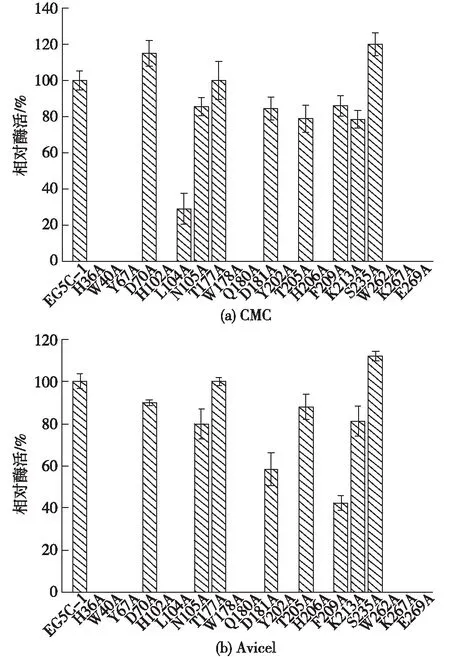
图3 EG5C-1的丙氨酸突变体对CMC和Avicel相对的水解酶活Fig.3 Relative hydrolytic activities of alanine-substituted mutants of EG5C-1 against CMC and Avicel
将丙氨酸突变体进行发酵表达后,以羧甲基纤维素钠和微晶纤维素为底物测定其相应的酶活,结果如图3。由图3可知:总体而言,大多数突变体对于CMC和Avicel底物的活性均显著下降,其中,芳香族氨基酸和极性氨基酸残基的突变尤其显著,如W40A、Y67A和W178A,当它们突变为丙氨酸后,其突变体对于2种底物几乎都失去了相应的活性。而突变体D70A与原始酶相比,其对于CMC底物的酶活提高了1.15倍;突变体S235A对于CMC和Avicel的酶活分别提高了约1.20倍和1.10倍。此外,突变体T177A仍具有与原始酶相似的酶活。为了探究D70、T177和S235位点上被其他氨基酸残基取代对酶活的影响,选择这3个位点与A234一同进行饱和突变。
2.3 饱和突变结果
分别使用在饱和突变位点D70、T177、A234和S235含简并碱基NNN的引物随机突变成任意氨基酸,通过测序和序列比对确定突变位点与特定氨基酸密码子一致。将饱和突变体进行发酵表达后,以羧甲基纤维素钠和微晶纤维素为底物测定其相应的酶活,结果如图4。由图4可知:将谷氨酰(D)置换为天冬氨酸(Q)对酶的活性影响最大,与原始酶相比,其对于CMC和Avicel的比酶活分别提高了1.5倍和1.3倍。同样,在S235饱和突变中,最优突变体对于CMC和Avicel 的比活性分别提高了1.4倍和1.2倍。然而,T177和A234位点的饱和突变体的酶活均没有显著的提升。
2.4 组合突变结果
将D70和S235饱和突变中的最优突变体进行组合突变,得到突变体D70Q/S235W,经发酵表达后,以羧甲基纤维素钠和微晶纤维素为底物测定其相应的酶活,结果如图5。由图5可知:与两个单一的突变体D70Q和S235W相比,组合突变体D70Q/S235W的酶活都有所提高,而与原始酶EG5C-1相比,D70Q/S235W对于CMC和Avicel的酶活分别提高了2.1倍和1.7倍,提高效果显著。
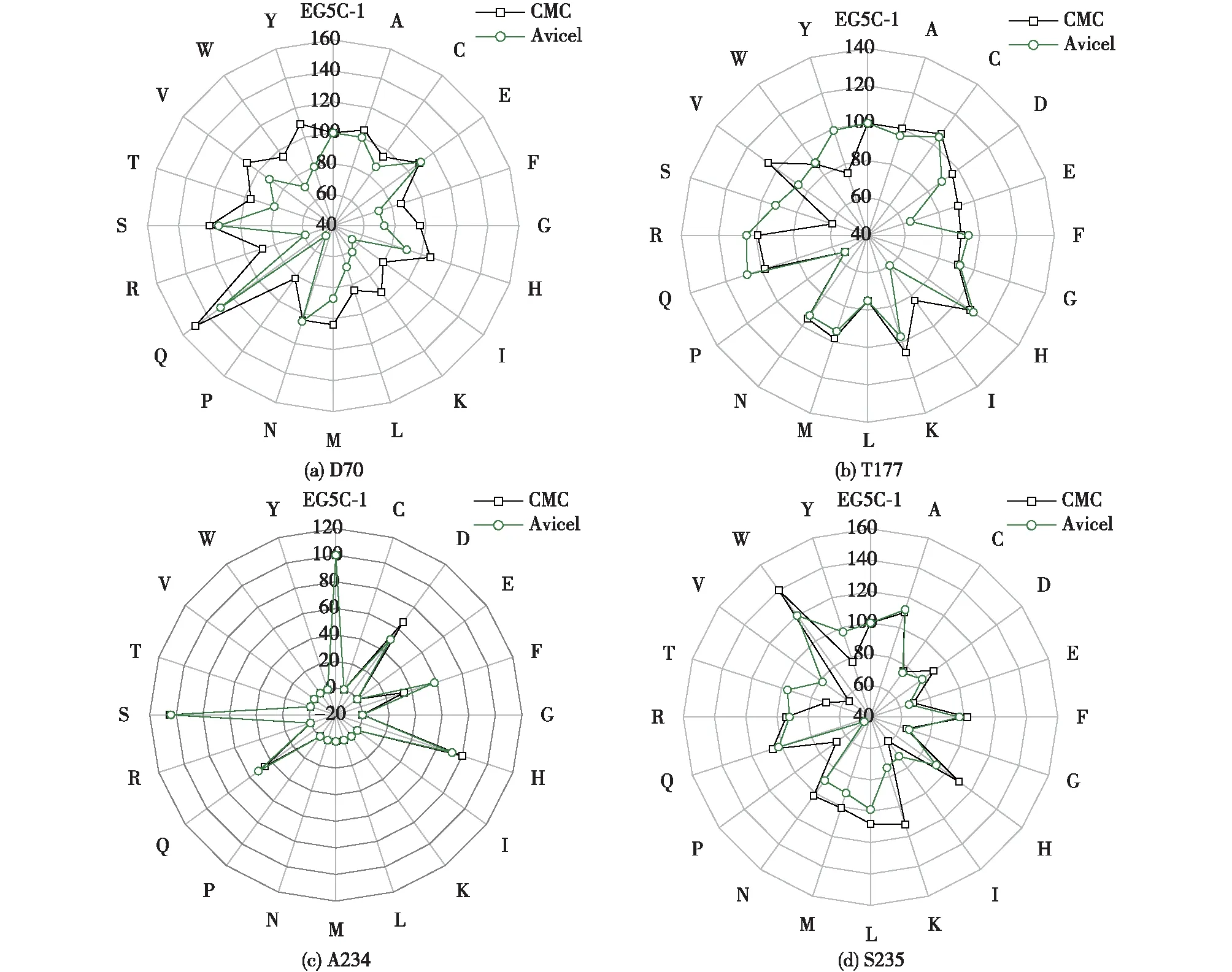
图4 D70、T177、A234和S235的饱和突变与原始酶EG5C-1对CMC和Avicel相对的水解酶活Fig.4 Relative hydrolytic activities of wild-type EG5C-1 and variants obtained through saturation mutagenesis of residues D70, T177, S234, and S235 against CMC and Avicel
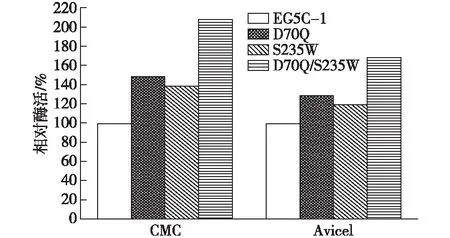
图5 组合突变体D70Q/S235W对CMC和Avicel相对的水解酶活Fig.5 Relative hydrolytic activities of combined mutants of D70Q/S235W against CMC and Avicel
2.5 D70Q/S235W酶学性质分析
2.5.1 D70Q/S235W的最适温度及热稳定性
以CMC作为底物测定不同温度下酶活的变化,结果如图6所示。由图6可知:原始酶EG5C-1的最适温度为60 ℃,而所有突变体D70Q、S235W和D70Q/S235W均显示出与EG5C-1相似的最适温度。当温度为30~60 ℃时,原始酶及突变体的酶活逐渐升高;当温度高于60 ℃时,原始酶及突变体的酶活显著下降。
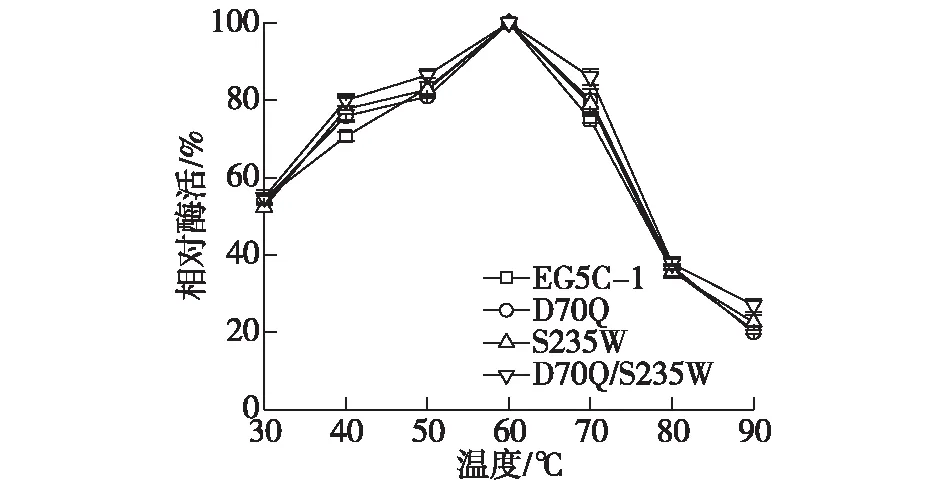
图6 原始酶EG5C-1及其突变体酶的最适反应温度Fig.6 Optimum temperature of the wild type EG5C-1 and its variants
酶的热稳定性结果见图7。由图7可知:原始酶EG5C-1在50 ℃下保温2 h后,仍然保留了80%以上的酶活,而所有突变体D70Q、S235W、D70Q/S235W均显示出与EG5C-1相似的温度稳定性。原始酶EG5C-1在60 ℃下保温2 h后,酶活保留了约70%左右,而突变体中D70Q/S235W仍然保留了80%的酶活,表明突变体酶的温度稳定性相比于原始酶有所提高,这对于工业生产具有很大的帮助。
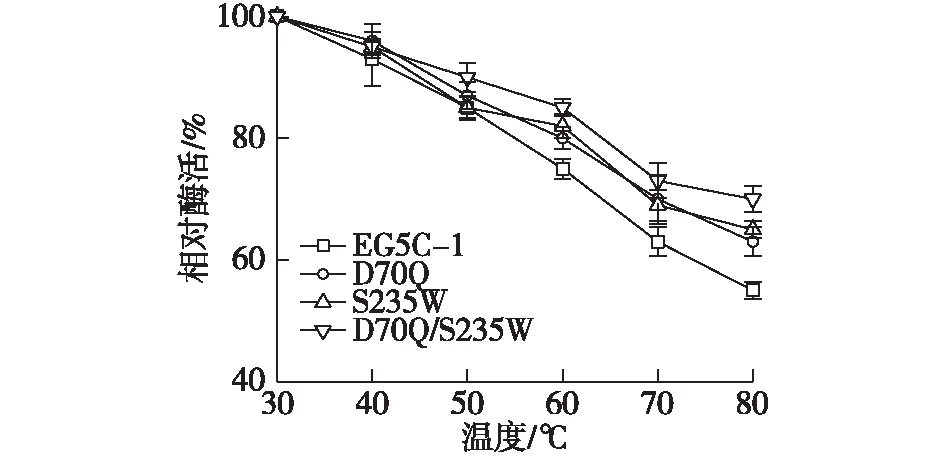
图7 原始酶EG5C-1及其突变体酶的温度稳定性Fig.7 Thermal-stability of the wild type EG5C-1 and its variants
2.5.2 D70Q/S235W的最适pH及pH稳定性
以CMC作为底物测定不同pH下酶活力的变化,结果如图8所示。由图8可知:原始酶EG5C-1与所有突变体D70Q、S235W、D70Q/S235W的最适pH均为6.0,随着pH的逐渐升高,酶活逐渐降低,表明EG5C-1与突变体酶都是偏酸性的酶。

图8 原始酶EG5C-1及其突变体酶的最适反应pHFig.8 Optimum pH of the wild type EG5C-1 and its variants
图9为原始酶EG5C-1及其突变体酶的pH稳定性。由图9可知:原始酶EG5C-1与突变体酶在pH 为5.0~ 9.0时的稳定性较好,孵育2 h后仍然保留了70%以上的酶活,而当pH为3.0~5.0时,原始和突变体酶的稳定性都较低,表明该酶不适于保存在强酸的条件下。同时,在不同pH条件下,原始酶EG5C-1 保留的酶活相比于其他所有突变体都较低,表明突变体的pH稳定性得到了提高。
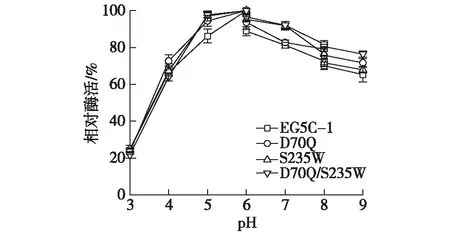
图9 原始酶EG5C-1及其突变体酶的pH稳定性Fig.9 pH stability of the wild type EG5C-1 and its variants
2.6 D70Q/S235W水解产物分析
以CMC、Avicel和PASC为底物,用TLC分析双突变体D70Q/S235W的水解作用模式,结果如图10所示。由图10可知:突变体D70Q/S235W对于不同的底物都具有很好的水解效果,且对于每一种底物的最终产物都主要为纤维二糖和纤维三糖;双突变体对于可溶性底物CMC和不溶性底物Avicel、PASC的水解作用模式与原始酶EG5C-1的作用模式相似[14],表明D70和S235位点上氨基酸残基的改变没有对产物的释放造成影响。

M为标准糖;G1—葡萄糖;G2—纤维二糖;G3—纤维三糖;G4—纤维四糖;1、4、7—对照;2、5、8—EG5C-1水解CMC、Avicel和PASC的产物;3、6、9—D70Q/S235W水解CMC、Avicel和PASC的产物图10 EG5C-1和突变体D70Q/S235W对不同纤维素基质的水解产物的薄层色谱分析Fig.10 TLC analysis of the hydrolysis products from various cellulosic substrates by EG5C-1 and mutant D70Q/S235W
3 结论
纤维素酶作为一种重要的工业催化剂,广泛应用于各个领域,然而由于纤维素复杂的组成结构,往往天然的单一纤维素酶催化效率很低。本文通过同源建模和分子对接对蛋白质结构分析,确定了持续性内切葡聚糖酶EG5C-1活性位点结构,并使用定点突变和饱和突变等方法设计得到了具有较高催化活性的突变体D70Q/S235W。
组合突变体D70Q/S235W在60 ℃和pH 6.0时表现出与原始酶EG5C-1相似的最适温度和最适pH,且与原始酶相比,组合突变体具有更好的温度和pH稳定性,为进一步研究和开发工业用酶奠定了基础。
