异质盐胁迫对杜梨幼苗生长及光合特性的影响
2022-10-21李永涛王霞马丙尧胡丁猛王清华李宗泰囤兴建许景伟
李永涛王霞马丙尧胡丁猛王清华李宗泰囤兴建许景伟
(1.山东省林业科学研究院,山东 济南 250014;2.山东黄河三角洲森林生态系统国家定位观测研究站/盐碱地生态修复工程技术中心,山东 东营 257000)
土壤盐渍化是影响黄河三角洲地区植物生长和生物多样性的主要限制因素之一,该区盐碱地范围广、面积大、生产力低,大量长期处于撂荒状态,如何有效利用和改善这些资源成为该区域亟待解决的问题[1,2]。研究表明,由于受微地形、土壤质地、土壤结构、降水、蒸发、覆盖等因素的影响[3,4],土壤空间异质性被认为是自然生态系统的一个普遍特征,盐渍土空间分布的不均匀性是其中的一个重要表现特征[5,6]。植物能够在盐渍土中维持生长,土壤盐分的分布不均匀性起着重要作用。因此,了解土壤盐分异质性和植株生长的互馈关系对提高盐碱地使用率具有十分重要的意义。
盐胁迫对植株生长的抑制引起植株生长缓慢、叶片气孔关闭、叶细胞失水及胞间CO2浓度下降等,从而影响植株的生长发育[7]。异质盐胁迫条件下,植物受抑制的程度可得到部分缓解。研究表明,与均匀盐胁迫相比,根系异质盐胁迫减轻其对小麦地上部、根系生长及光合作用的抑制作用,缓解盐胁迫对小麦的伤害[8];局部低盐胁迫条件下绒毛白蜡生长高于盐分均匀胁迫,并通过降低叶片中Na+、Cl-和脯氨酸的积累增加K+含量,缓解盐胁迫的抑制[9,10];不均匀盐胁迫中大洋洲滨藜的生长量高于同等含盐量的均匀盐分胁迫,盐分的差异化分布促进植物生长[11];此外,根系异质盐胁迫对高粱[12]、紫花苜蓿[13]等植物生长的抑制也均低于均匀盐胁迫。
杜梨(Pyrus betulifoliaBunge)为蔷薇科(Rosaceae)梨属(Pyrus)落叶乔木,是我国原生梨属植物,具有较强的耐盐能力,也是黄河三角洲地区主要绿化观赏树种之一。基于异质盐胁迫下的植物耐盐性研究更接近于植物生长实际环境。本研究通过异质盐胁迫分根试验,对杜梨幼苗的生长状况、光合特征、叶片氮含量及叶面温度等进行分析,以揭示杜梨异质盐胁迫下的生长特征及光合特性,探索局部盐分淡化技术在盐碱地上应用的可行性,为盐碱地原土绿化提供理论依据。
1 材料与方法
1.1 供试材料
试验于2021年在山东省林业科学研究院东营林草试验站进行。供试材料为杜梨当年容器播种苗。无纺布容器规格为直径5.5 cm,高度7.5 cm,之中基质为草炭+珍珠岩+蛭石,其配比为7∶2∶1。4月中旬在温室内将层积好的杜梨种子点播到无纺布容器内,5月底将容器苗移到炼苗场备用。7月初选择生长一致、株高20 cm左右的杜梨容器苗,去除外层无纺布,栽植于具有中间隔板的分根容器中间位置,以保证后期根系均等向隔板两侧生长。
分根容器制作:采用PVC塑料管(直径20 cm,高度35 cm)和铝塑板(厚度3 mm)制作分根容器,结构示意图(图1a)与效果图(图1b)描述和制作过程参照Feng等[14]的研究,具体规格略有调整。为了便于移动,分根容器制作完成后放入塑料花盆中(上口直径23 cm)。将通用的建筑水洗沙过2 mm筛,等量装入分根容器两侧,之前容器底部放置少许粗大的石砾,以便于排水。

图1 杜梨幼苗沙培分根容器结构示意图(a)与效果图(b)
1.2 试验设计
试验共设置5个处理,包括对照非盐胁迫(0/0)、根系异质盐胁迫(0/100、0/200)、均匀盐胁迫(100/100、200/200),盐胁迫浓度分别为0、100、200 mmol·L-1。每处理重复3次。
为保证幼苗在分根容器两侧均匀生长,先配制1/3改良Hoagland’s营养液进行浇灌(每5 d浇灌1次),10 d后浇灌含有不同浓度NaCl的1/3改良Hoagland’s营养液。其中,分根容器内无盐胁迫一侧直接浇灌1/3改良Hoagland’s营养液,盐胁迫一侧分别浇灌相应NaCl浓度的1/3改良Hoagland’s营养液。采用过量浇灌方法,试验开始后每10 d浇灌1次,每次浇灌以营养液从底部渗透孔浸出为宜。
1.3 测定指标及方法
1.3.1 株高及生物量测定 盐胁迫处理60 d后,采用塔尺测定幼苗株高。分地上部分和地下部分收获整株。根系取样时采用自来水缓慢冲洗,冲走分根容器两侧内的沙子。地上和地下部生物量的测定采用烘干法,即于烘箱中105℃杀青0.5 h后70℃烘48 h至恒重,称重。
1.3.2 光合指标测定 幼苗盐胁迫处理15、30 d和45 d时,利用Li-6400便携式光合仪(LI-COR,USA)测定叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。测定时采用LED红/蓝光源,光强为1 000 μmol·m-2·s-1,测定时间为10∶00—11∶00,每处理测3株。
采用OS-30p+快速植物胁迫测量仪(美国)测定叶片叶绿素荧光参数,每株选取3片叶,用暗叶夹夹住叶片中间部位暗适应40 min后,测定其Fv/Fm值,每处理3个重复。
1.3.3 叶片叶绿素含量、氮含量及叶片温度测定 幼苗盐胁迫处理15、30 d和45 d时,利用TYS-4N植物营养测定仪(中国)同步测定幼苗叶片的SPAD值、氮含量及叶片温度的变化,每处理3个重复。
1.4 数据处理
采用Microsoft Excel 2010、DPS 15.10进行数据处理、作图与分析,采用单因素方差分析和最小显著差异法(LSD)比较不同数据组间的差异,显著性水平设定为P<0.05。
2 结果与分析
2.1 盐胁迫对杜梨幼苗生长的影响
由表1可知,盐胁迫下杜梨幼苗生长受到明显抑制,且随着盐胁迫浓度升高,均匀盐胁迫和异质盐胁迫处理均降低幼苗株高和总生物量,两者间差异显著。随着胁迫时间的延长,均匀盐胁迫100/100和200/200处理下,杜梨植株生长受阻,叶片变小,出现叶尖叶缘干枯或叶片萎蔫脱落等盐害症状,幼苗分别在盐胁迫41 d和26 d后死亡。其株高相对生长率分别为5.42%和0.05%,而异质盐胁迫0/100和0/200处理下相对生长率分别为26.96%和16.60%,较均匀盐胁迫显著增加。与对照(0/0,无盐胁迫)相比,盐胁迫显著降低植株的根冠比。异质盐胁迫(0/100、0/200)和均匀盐胁迫(100/100、200/200)处理下地上生物量分别较对照降低11.86%、17.70%和32.57%、60.88%,而根系则分别降低30.52%、43.03%和54.29%、73.56%,较地上部降幅大。
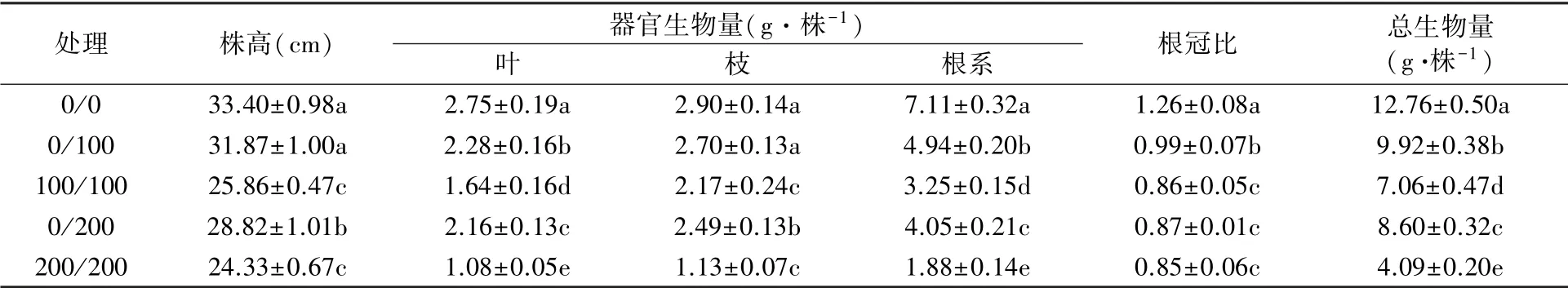
表1 不同盐胁迫处理下杜梨幼苗生长及生物量
2.2 盐胁迫对杜梨幼苗叶片SPAD值和荧光参数Fv/Fm的影响
由图2可知,盐胁迫下,叶片SPAD值随着盐胁迫浓度的增加而降低。与对照(0/0)相比,均匀盐胁迫100/100和200/200处理,15 d时SPAD值显著下降,分别降低11.67%和39.87%;盐胁迫30 d时,100/100处理植株的SPAD值显著下降达28.95%,而此时200/200处理植株已死亡。异质盐胁迫0/100和0/200处理下,植株未出现死亡,15 d和30 d时其SPAD值与对照相比无显著差异,45 d时则差异显著。
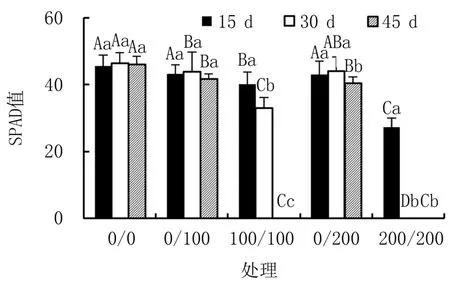
图2 不同盐胁迫下杜梨幼苗叶片的SPAD值
图3显示,盐胁迫下,叶片叶绿素荧光参数Fv/Fm随胁迫时间延长和浓度增加呈现下降趋势,其中,均匀盐胁迫处理下降显著,且降幅明显高于异质盐胁迫。盐胁迫15 d时,异质盐胁迫0/100处理叶片Fv/Fm较对照无显著差异,0/200处理较对照差异显著,但降幅较小,两者较同浓度均匀盐胁迫100/100和200/200处理相应多出24.14%和82.01%;30 d时,均匀盐胁迫200/200处理植株已死亡,0/100处理较均匀盐胁迫100/100多出73.77%,差异显著。45 d时,均匀盐胁迫处理下植株均已全部死亡。
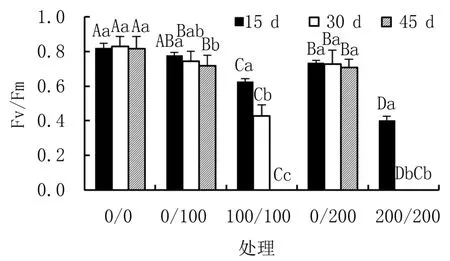
图3 不同盐胁迫下杜梨幼苗叶片的Fv/Fm
2.3 盐胁迫对杜梨幼苗叶片光合特性的影响
由图4可知,盐胁迫降低杜梨叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。随着盐胁迫浓度升高和时间延长,均匀盐胁迫下Pn、Gs、Tr与对照差异显著,而异质盐胁迫下0/100处理15 d时与对照差异不显著,且异质盐胁迫受抑制程度明显低于均匀盐胁迫。盐胁迫15 d时,异质盐胁迫0/100和0/200处理与同浓度均匀盐胁迫100/100和200/200处理相比,Pn、Gs、Ci及Tr指标分别多出26.55%、137.11%、28.18%、87.90%和93.36%、104.08%、0.97%、71.52%;30 d时,均匀盐胁迫200/200处理植株死亡,异质盐胁迫0/100与均匀盐胁迫100/100处理相比,Pn、Gs、Ci、Tr分别多出169.13%、471.74%、626.94%、625.51%;45 d时,均匀盐胁迫处理植株均已死亡,而异质盐胁迫下植株均正常生长。

图4 不同盐胁迫下杜梨叶片的光合特性
2.4 盐胁迫对杜梨幼苗叶片氮含量和叶面温度的影响
由图5可知,异质盐胁迫与均匀盐胁迫下杜梨幼苗叶片氮含量的变化规律并不一致。随着盐浓度升高,15 d时,异质盐胁迫下叶片氮含量略有增加;而30 d时,叶片氮含量先升高后降低;45 d时,则呈下降趋势。随着盐胁迫时间延长,15~45 d期间,异质盐胁迫下叶片氮含量均呈现出先升高后降低趋势,且差异显著;均匀盐胁迫则呈下降趋势,100/100和200/200处理下植株分别于41 d和26 d后陆续死亡。
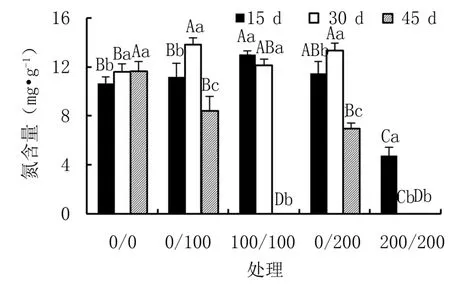
图5 不同盐胁迫下杜梨幼苗的叶片氮含量
由图6可知,异质和均匀盐胁迫下,植株叶面温度略高于对照,并随胁迫时间延长呈现“低-高-低”的单峰趋势。胁迫15~30 d,除均匀盐胁迫200/200处理植株死亡外,其它异质盐胁迫处理的叶面温度与均匀盐处理无显著差异;45 d时,均匀盐胁迫植株均已死亡。

图6 不同盐胁迫下杜梨幼苗叶片的叶面温度
3 讨论与结论
盐碱胁迫下植株的生长状态是耐盐碱能力强弱的最直接表现,其生长量和生物量会随胁迫程度的变化而受到不同程度的抑制[15,16]。本研究表明,不同盐胁迫处理方式均降低杜梨地上、地下部生物量和株高,而异质盐胁迫下植株的生物量则显著高于均匀盐胁迫,这与在橙子[17]、绒毛白蜡[9]上的研究结果一致。异质盐胁迫0/100和0/200处理下,杜梨植株能够保持正常生长,而均匀盐胁迫100/100和200/200处理下,植株分别在41 d和26 d后死亡,这说明该盐胁迫浓度高于其致死胁迫浓度,而异质盐胁迫可以有效缓解高盐胁迫对杜梨生长的抑制。有研究表明,植物生物量配置模式的自我调节是适应盐胁迫的可塑性机制之一,往往符合最优分配理论的预测[18,19]。本研究中,杜梨的根冠比随着盐胁迫浓度升高而下降的趋势明显,而异质盐胁迫下降有所缓解,这与小麦[8]、大洋洲滨藜[11]根冠比增大的结果不同。其原因主要与本试验设置的盐浓度高于杜梨的致死浓度有关,该浓度盐胁迫下根系生物量明显下降,而植株地上部生物量的下降趋势比根系更显著,这是导致根冠比下降和后期均匀胁迫处理下植株死亡的主要原因。由此看出这种生物量分配的差异性能够减轻土壤盐渍化对根系的危害[20]。
光合参数是评价植物盐胁迫能力的重要指标,主要表现为离子渗透和毒害作用导致的植株代谢紊乱、叶绿素含量降低,从而影响植物的光合作用[21]。本研究表明,随着盐胁迫浓度升高和胁迫时间延长,杜梨叶片SPAD值、Fv/Fm均降低,受到明显的光抑制影响,光合作用减弱,此时Pn、Gs、Ci和Tr也表现出一致的协同作用,且均匀盐胁迫处理45 d时植株全部死亡。而异质盐胁迫下,叶片光合特征参数较均匀盐胁迫有所增大,表明异质盐胁迫能缓解均匀盐胁迫对光反应的抑制,光合作用虽有所减弱,但仍能维持植株正常生长,这与异质盐胁迫对番茄[22]、葡萄[23]的影响结果类似。
植物氮素含量和叶面温度是表征植物生长的重要营养和生理指标,也是反映植物生命体征的重要参数。本研究盐处理15~45 d期间,无盐胁迫(对照)下叶片氮含量呈现逐渐升高趋势,而异质盐胁迫下叶片氮含量呈现出先升高后降低趋势,均匀盐胁迫则呈下降趋势。其主要原因为:本试验测定时间为夏季,对照植株随着生长和光合产物的积累及转化,叶片氮含量不断增加。异质盐胁迫与均匀盐胁迫相比显著减轻盐分对叶片的损伤,30 d时仍呈现升高趋势,但随着胁迫时间的延长,过多的Na+会抑制叶片中氮素的积累,从而导致氮含量下降。均匀盐胁迫由于浓度较高,光合作用受到严重抑制,导致植物体内一系列含氮化合物的代谢紊乱,叶片氮含量不断下降[24,25]。
植株的温度值是其能量代谢的结果,其叶温的变化受多种外界条件的影响,它常被用来反馈植物水分状态、气孔的关闭和蒸腾的衰减[26]。异质和均匀盐胁迫下,杜梨的叶面温度略高于无盐胁迫,其原因可能是盐胁迫扰乱了植株体内的水分平衡,水分亏缺和冷却水分的不足导致叶温升高,尤其是夏季炎热时蒸腾降温功能失调。随盐胁迫时间延长,对照和处理叶面温度均呈现“低-高-低”的单峰趋势,这主要与测定时气温的高低呈直接关联性,本研究均匀盐胁迫45 d时植株均已死亡。
杜梨作为黄河三角洲地区主要的耐盐乡土树种,已在轻中度盐碱地区得到广泛应用。本试验在全株水平上研究杜梨幼苗对异质性盐胁迫的响应,这为重盐碱地局部淡化原土栽植杜梨提供了理论基础。同时,本研究为方便控制试验的开展,采用了杜梨苗期盆栽试验,在时间和空间应用上具有一定的局限性。此后,随着微域土壤学的提出[27],异质性盐胁迫对大田环境条件下苗木整个生育周期的影响有待于进一步研究。
