非人灵长类动物肠道寄生虫与宿主相互作用的研究进展
2022-10-19陆媚静范鹏来周岐海
陆媚静, 范鹏来, 孙 涛*, 周岐海*
(1.珍稀濒危动植物生态与环境保护教育部重点实验室(广西师范大学), 广西 桂林 541006;2.广西珍稀濒危动物生态学重点实验室(广西师范大学), 广西 桂林 541006)
非人灵长类动物作为进化史上与人类亲缘关系最近的类群,在认知能力、繁殖和生育等方面与人类具有高度同源性[1],这使得非人灵长类动物体内共生生物类群和病害,例如寄生虫、致病性病毒和细菌等,往往相比其他动物具有更高的人畜共患风险。一直以来,非人灵长类动物携带的致病性病毒和细菌受到高度重视,目前已开展了众多关于非人灵长类动物病毒性感染模型的研究,例如艾滋病病毒感染模型、乙型肝炎病毒感染模型[1-2]。然而,肠道寄生虫作为非人灵长类动物的常见共生生物类群和病害之一,其对非人灵长类动物的生存和繁殖影响深远[3]。
肠道寄生虫主要通过直接的病理作用或者宿主的健康状况来影响动物的生存和繁殖[4-5]。例如,通过影响宿主的营养状况,增加其能量消耗,降低其对资源和配偶竞争的能力[6],严重情况下会造成宿主失血、组织损伤、自然流产、先天畸形和死亡[7-8]。在长期进化过程中,非人灵长类动物与寄生虫之间形成稳定的相互适应关系[9]。然而,近年来,随着人类与非人灵长类动物频繁接触(例如生态旅游)或侵入、破坏非人灵长类动物的栖息生境,使得非人灵长类动物与人类的接触机会增加,种间传染的风险也随之加大,从而不同程度地改变着非人灵长类动物与肠道寄生虫的交互关系。非人灵长类动物与肠道寄生虫交互作用及影响机制一直是动物生态学和保护生物学研究的重要领域之一,本文系统地总结非人灵长类动物与肠道寄生虫相互作用,旨在为非人灵长类动物肠道寄生虫研究和公共健康卫生提供一定的理论框架和新思路。
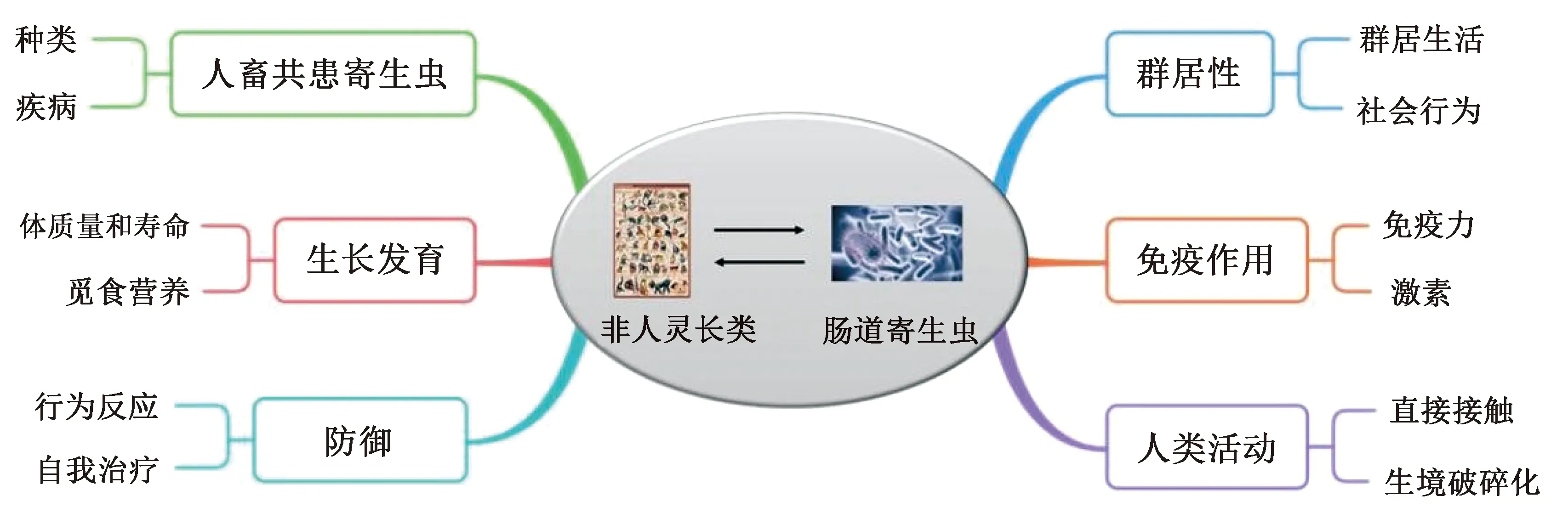
图1 肠道寄生虫在非人灵长类动物各方面的作用[10-15]Fig.1 Role of gastrointestinal parasites in various aspects of non-human primates
1 非人灵长类动物常见致病性人畜共患肠道寄生虫种类及其危害
1.1 种类
非人灵长类动物寄生着各种各样的寄生虫,其中许多通过胃肠道传播,且具有传播途径多样的特点。一项有关里约热内卢动物园圈养动物肠道寄生虫的研究指出,非人灵长类动物肠道寄生虫检出率是最高的,达到68.2%[16]。肠道寄生虫可以作为栖息地使用强度和非人灵长类动物健康程度的指示生物,在疾病预防和爆发、濒危动物保护方面发挥重要作用[4,17-18]。国内已有对野生或圈养非人灵长类动物肠道寄生虫的研究报道,例如:吕超超等[19]在太行野生猕猴Macacamulattatchelien中检测出阿米巴原虫Amoebasp.、鞭虫Trichurissp.、泡翼线虫Physalopterasp.、蛲虫Enterobiussp.和蛔虫Ascarissp.等肠道寄生虫;杨天友等[20]在贵州麻阳河保护区黑叶猴Trachypithecusfrancoisi中检测出蛔虫、短刺节口线虫Ancylostomatidae和蛲虫;吴伟等[21]在安徽黄山雄性短尾猴Macacaarctoides中检测出钩虫Ancylostomasp.、蛔虫、结肠小袋纤毛虫Balantidiumcoli、蓝氏贾第鞭毛虫Giardialamblia和溶组织内阿米巴Entamoebahistolytica等,这些发现对指导我国非人灵长类动物寄生虫病的防控和人类公共卫生安全具有重要意义。
非人灵长类动物中,致病性寄生虫主要有线虫、吸虫、绦虫和原虫。线虫是非人灵长类动物中最常见的肠道寄生蠕虫。对中非共和国德赞噶-桑哈国家公园(Dzanga-Ndoki National Park)大猩猩Gorillagorilla和黑猩猩Pantroglodytes的研究表明,它们线虫感染率达到92%~94%[22];此外,蠕形住肠线虫Enterobiusvermicularis、毛首鞭形线虫Trichuristrichiura、粪类圆线虫Strongyloidesstercoralis在所有非人灵长类动物中的检出率接近100%[23]。在非人灵长类动物中常见的吸虫有日本血吸虫Schistosomajaponicum、肝片吸虫Fasciolahepatica[24]。由棘球绦虫属绦虫引起的棘球蚴病是一种广泛分布的人畜共患病,在流行地区的非人灵长类动物中已有报道(汤基猕猴Macacatonkeana[25]、食蟹猕猴Macacafascicularis、猕猴Macacamulatta和狮尾猕猴Macacaslienus[26])。
非人灵长类动物体内致病性人畜肠道寄生原虫主要有阿米巴原虫、蓝氏贾第鞭毛虫和隐孢子虫Cryptosporidiumparvum[17,27]。阿米巴原虫广泛存在于野生哺乳动物,是感染率最多且宿主范围最广的人畜共患肠道寄生虫,常见的主要有结肠内阿米巴Entamoebacoli、溶组织阿米巴[27]。蓝氏贾第鞭毛虫主要通过受污染的食物、水等途径传播,可使人类和非人灵长类动物腹痛和腹泻[28]。隐孢子虫是水源性寄生虫,主要通过粪口传播途径在人猴之间交叉传播[29]。
1.2 危害
肠道寄生虫可引发宿主失血症、组织损伤、自然流产、先天性畸形,缩短寿命,甚至死亡等[7]。对日本猕猴Macacafuscata的研究发现,隐孢子虫引起的腹泻导致小肠病变,例如肠系黏膜增大,肠上皮细胞高度变异,单个肠上皮细胞坏死,还会出现急性支气管炎等[30]。由肠道寄生虫引起的传染病已经成为开展濒危野生非人灵长类动物保护工作的主要威胁,且其携带的寄生虫多为人畜共患肠道寄生虫,可以传播给共享同一栖息地的人类,威胁人体健康[31]。目前,非人灵长类动物常见的致病性人畜共患肠道寄生虫主要有溶组织内阿米巴、结肠小袋纤毛虫、隐孢子虫、钩虫、蛔虫、毛首鞭形线虫(表1),这些寄生虫常常使宿主患阿米巴病、结肠小袋虫病等,可引起宿主结肠炎、肠外肿脓、肠道黏膜破坏等症状,严重的甚至造成宿主死亡[32-33]。
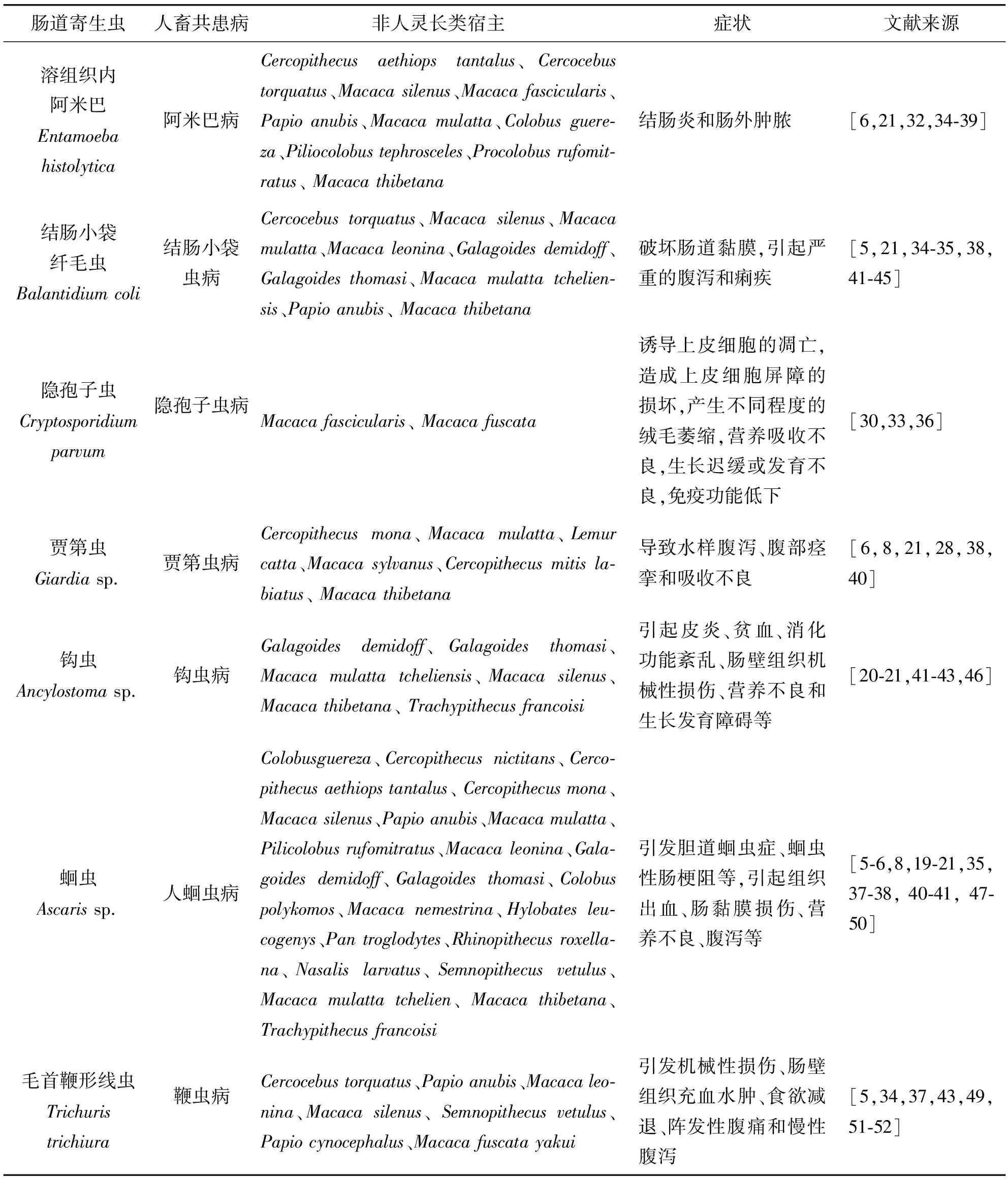
表1 非人灵长类动物常见的致病性肠道寄生虫及其危害
2 非人灵长类动物的社会行为与肠道寄生虫的相互作用
集群生活动物之间近距离接触或身体接触(例如梳理、抱团、咬人、交配等)的社会互动行为是必不可少的,对群体凝聚力和社会结构的建立和维持具有重要意义[53]。非人灵长类动物的群居生活可以增加资源的获取,减少捕食风险,提高觅食效率,但同时也增加了寄生虫传播的风险[54]。鉴于社会网络可以提供传染性寄生虫传播路径的相关信息,有学者采用社会网络分析法确定非人灵长类动物种群或群体中寄生虫的传播[55]。在非人灵长类动物中,寄生虫的丰富度与个体社会接触的频率呈正相关关系[56]。Rimbach等[57]对哥伦比亚圣胡安大庄园(Hacienda San Juan de Carare)棕蜘蛛猴Ateleshybridus社交网络的研究发现,高度联系的个体具有较高的肠道寄生虫丰富度;在白颈白眉猴Cercocebustorquatus中也同样发现,在邻近网络中,处于中心位置、联系密切并倾向于转换群体的个体,其感染蠕虫风险更高,但整个群体对感染原虫的风险无明显差别[34]。在群居性非人灵长类动物中,宿主行为和生理上的变化也会影响肠道寄生虫的传播,最终影响宿主健康。对日本猕猴梳理行为与寄生虫感染关系的研究发现,梳理网络中雌性的中心性与某些线虫的感染风险呈正相关,但与体表寄生虫(例如虱子)的感染成负相关,这与它们相互接触和梳理有关[58]。这表明,宿主的梳理行为可以增加肠道寄生虫的感染,但同时减少了体表寄生虫的感染[10]。此外,非人灵长类动物群体中,个体之间也会由于社会等级的不同而产生肠道寄生虫易感性差异。一项关于屋久岛(Yakushima Island)雌性日本猕猴Macacafuscatayakui宿主的优势等级与线虫传播关系的研究发现,群体社会网络中雌性的中心性与优势等级表现出正相关关系,而优势等级与寄生虫丰富度呈明显负相关关系[59]。受肠道寄生虫感染的非人灵长类动物也会由于自身的病理作用或寄生虫的操控作用引起行为改变,最终占据不同的网络地位,改变群体的社会结构[60]。寄生虫的传播也会随着群体规模的增大而增加,例如在斯里兰卡(Sri Lanka)西紫脸叶猴Semnopithecusvetulus中发现,群体规模越大,毛首鞭形虫、类圆线虫、蛔虫和原生动物的感染率也越大[49]。而肠道寄生虫也会引起非人灵长类动物活动水平的下降,通常表现为嗜睡、食欲下降[61]。一项有关乌干达(Uganda)基巴莱国家公园(Kibale National Park)红疣猴Piliocolobustephrosceles的行为和鞭虫感染关系的研究指出,感染了鞭虫的红疣猴会增加休息时间,而减少其他更耗能的行为(例如梳理行为和交配行为)[61],而对黑脸绿猴Chlorocebusaethiops驱虫后发现,其休息时间减少了,日漫游时间增加了[62]。综上表明,非人灵长类动物的群居性生活和社会行为增加了肠道寄生虫传播的可能性,而肠道寄生虫也对非人灵长类动物的群居生活和社会行为产生一定影响。然而,目前关于非人灵长类动物群体对肠道寄生虫传播的研究主要集中在群体内部联系的影响方面,对于跨物种传播以及群体规模传播的研究还比较少[10],未来可给予更多关注。
3 肠道寄生虫对非人灵长类动物生长发育的影响
非人灵长类动物的生长与繁殖受肠道寄生虫丰富度和患病率的影响。例如,蠕虫和原生动物可以通过病理作用和降低非人灵长类动物的身体状况直接或间接地影响其自身的生长和繁殖[11]。对于寄生在非人灵长类动物中的肠道寄生虫来说,其所需的能量来自非人灵长类动物体内,它与非人灵长类动物相互适应又受非人灵长类动物的体质量、寿命和饮食等影响,最终又可改变非人灵长类动物的行为、外观和形态,抑制其生长和繁殖,甚至导致死亡[63]。例如,坦桑尼亚(Tanzania)一个野生黑猩猩感染食道口线虫Oesophagostomumsp.后,出现体质量减轻和腹泻的症状[61]。Springer等[64]在比较6种狐猴(贝氏倭狐猴Microcebusberthae、倭狐猴Microcebusmurinus、肥尾鼠狐猴Cheirogaleusmedius、科氏倭狐猴Mirzacoquereli、红额美狐猴Eulemurrufus、维氏冕狐猴Propithecusverreauxi)肠道寄生虫的研究中也发现,体质量最小和寿命最短的倭狐猴其肠道寄生虫丰富度最高,而体质量最大和寿命最长的维氏冕狐猴其肠道寄生虫的丰富度最低。
肠道寄生虫可以影响非人灵长类动物的觅食营养,从而间接影响其生长发育。肠道既是非人灵长类动物进行食物消化、营养吸收的主要场所,也是肠道寄生虫的主要栖息地。因此,肠道寄生虫与非人灵长类动物的食物和营养之间存在密切联系。“恶性循坏”假说(vicious circle hypothesis)指出,当食物资源短缺,食物供应不足时,宿主的防御能力降低,可能更容易受到寄生虫感染,最后导致宿主身体状况变差,受寄生虫感染强度增高[65-66]。这与宿主免疫以及其他相互作用(例如稳态、生长和繁殖)竞争的营养能量需求过程之间的平衡有关[62]。当宿主面临营养可利用性减少时,它们可能被迫调整体内的生理状况,进而影响免疫的能量投入[3],从而更容易受到寄生虫感染[67];当宿主营养状况良好时,宿主则分配更多的资源用以抵抗寄生虫的侵害[68]。
肠道寄生虫被认为具有以下3个特点:1)利用宿主作为栖息地;2)与宿主形成营养依赖关系;3)对宿主造成“伤害”[14]。因此,肠道寄生虫在食物链、食物网以及生态系统中发挥着重要作用,它们可以通过营养相互作用在不同宿主间传播,从而提供许多关于非人灵长类动物生物学和食物网的有用信息,并且可以削弱非人灵长类动物消化营养物质、防御更多有害病原体的能力[69]。宿主-寄生虫系统的研究表明,寄生虫可能会导致宿主食物摄入量减少,产生营养压力[70]。当宿主蛋白质营养不良时,会降低肠道屏障功能的耐受性,导致蠕虫负荷增多[71]。
在非人灵长类动物中,宿主的觅食策略和觅食生境会影响肠道寄生虫的感染,且个体觅食生态差异影响寄生虫的暴露情况[12]。对69种非人灵长类动物(包括猕猴等)与寄生虫的关系总结发现,宿主食物中叶子的数量与寄生虫丰富度呈显著正相关关系,这与叶食性灵长类所需的食物资源有关;相对于果实性和虫食性非人灵长类动物来说,叶食性非人灵长类动物需要消耗更多的食物资源——树叶,所感染寄生虫的机会也随之增大[72]。此外,对乌干达基巴莱国家公园红疣猴的研究发现,种群大小、食物的可利用性和寄生虫感染指数相关,食物供应通过寄生虫感染间接影响红疣猴的种群数量[73];比较不侵袭农作物红疣猴和侵袭农作物东非黑白疣猴Colobusguereza的肠道寄生虫感染率发现,红疣猴的肠道寄生虫感染率更高,表明营养的增加可能促进了东非黑白疣猴对肠道寄生虫更为有效的免疫。目前关于肠道寄生虫对非人灵长类动物生长发育的影响研究还不够深入,但在人类中已经有更加深入的研究。一项关于400名儿童肠道寄生虫感染与生长发育的研究发现,感染了肠道寄生虫(例如蛔虫、鞭虫或钩虫)的儿童会显著发生消瘦、生长迟缓、体质量低下和恶性营养不良的情况[74]。这表明肠道寄生虫可以通过直接或间接的作用影响非人灵长类动物的生长发育。
4 非人灵长类动物的免疫作用与肠道寄生虫的相互作用
非人灵长类动物长期处于肠道寄生虫高负荷水平会降低宿主自身的免疫力,进而降低出生率和繁殖率,影响种群的数量和稳定;同时,非人灵长类动物携带的某些疾病也会传染人类,对人类健康造成威胁[75]。研究表明,蠕虫可以调节宿主免疫系统,使宿主失去清除寄生虫的能力,同时也抑制了许多特异性免疫反应的进行[76]。反过来,宿主自身的状况,例如年龄、性别、体质量、生殖状况和免疫抑制激素等,也影响寄生虫的传播和流行[46]。一项关于日本猕猴年龄、性别和季节对线虫影响的研究指出,宿主的免疫是直接传播线虫的重要媒介,非人灵长类动物年龄与寄生虫感染的关系呈现出先增后减的趋势,这被认为是宿主对寄生虫的适应性免疫有关[52]。在非人灵长类动物中,某些固醇类激素(例如睾酮、糖皮质激素等)的免疫抑制作用也可能直接影响肠道寄生虫的生命周期[51]。在黑猩猩中,睾酮和皮质醇的浓度与肠道寄生虫的丰富度呈显著正相关,等级地位越高的雄性个体,睾酮水平越高,蠕虫的丰富度也越高[77]。此外,虽然宿主可以通过自噬作用清除体内寄生虫,但寄生虫也可以通过宿主的自噬作用进化出自身的免疫抵抗机制,促进其自身的生长发育[78]。
5 非人灵长类动物对肠道寄生虫的防御
当非人灵长类动物对其肠道寄生虫的生理适应不足时,会产生避免或限制与寄生虫接触的行为反应,以应对高密度的肠道寄生虫感染[79]。在非洲类人猿(African great apes)中,黑猩猩、倭猩猩Panpaniscus等广泛存在的吞叶行为(leaf-swallowing),就是宿主为控制线虫和绦虫感染而产生的一种有意识的适应行为策略,因为整片树叶在肠道中增加了肠道寄生虫排出或肠道运输的可能性[80]。目前发现,亚洲非人猿的白手长臂猿Hylobateslar为驱逐寄生虫采取了与非洲类人猿类似的吞叶行为策略[81]。为减少肠道寄生虫感染的风险,非人灵长类动物也会采取回避的行为策略,例如交替使用栖息地或回避粪便,这支持了寄生虫回避假说(parasite avoidance hypothesis)[82]。研究发现山魈Mandrillussphinx和食蟹猕猴会避免食用被粪便污染的食物,从而减少与通过粪便传播的寄生虫(例如胃肠道线虫等)接触[83-84]。此外,非人灵长类动物还通过摄取具有药用价值的植物,避免或消除肠道寄生虫的感染。猴子和猿类被寄生虫感染时,会改变各自的饮食,转为食用能够缓解症状的树叶[14]。对屋久岛日本猕猴摄取的45种植物进行甲醇提取物药用价值分析发现:37%的植物提取物与治疗寄生虫、病原体和相关症状有关,且所有提取物均具有抗罗得西亚布氏锥虫Trypanosomabruceirhodesiense的活性;超过80%的提取物可以抑制恶性疟原虫Plasmodiumfalciparum和杜氏利什曼原虫Leishmaniaspp的生长;50%的提取物表现出对克氏锥虫Trypanosomacruzi抗原虫的潜力;还有部分提取物对曼氏血吸虫Schistosomamansoni的幼虫和成虫阶段有活性[85]。在野生大猩猩的研究中也发现,其摄取的植物具有药用价值,包括抗寄生虫、抗真菌、抗病毒等药理作用[86]。在猕猴、台湾猴MacacacyclopisSwinhoe和橄榄狒狒Papioanubis中发现,宿主自身具有不同的自愈能力,宿主感染血吸虫后会发生自愈现象,但其自愈机制目前还不清楚[87],未来可进一步深入探究。
6 非人灵长类动物及其肠道寄生虫对人类活动的响应
大多数非人灵长类动物的生态位与人类生态位重叠,空间重叠和资源共享改变了寄生虫的传播动态[15]。许多研究表明,寄生虫经常从野生或圈养的非人灵长类动物传播给共享栖息地的人类[18]。同时,人类活动可能会增加人类主导的景观中野生生物的多样性,并突出了寄生虫多样性分布的复杂格局[88]。对印度南部狮尾猕猴的研究表明,与非人类居住区附近的种群相比,生活在人类居住区附近的种群具有更高的肠道寄生虫感染率[43]。与此相反,研究者发现居住在人类居住区附近森林碎片的黑吼猴Alouattacaraya其肠道寄生虫更少,而居住在远离人类居住区附近的黑吼猴肠道寄生虫感染率更高,推测其原因是人类干扰降低了寄生虫与非人灵长类宿主的连通性[50]。在印度尼西亚巴厘岛(Bali Indonesia),人类直接接触或圈养食蟹猕猴会加剧其感染肠道寄生虫的风险[15]。张波等[47]对川金丝猴Rhinopithecusroxellana的研究发现,生态旅游增加了肠道寄生虫的感染率和丰富度,尤其是人类和非人灵长类动物接触频率的增加提高了野生动物直接接触或间接接触感染源的机会。在尼日利亚(Nigeria),盗食人类作物的橄榄狒狒群体比野生觅食群体的寄生虫感染率更高,表明作物盗食导致的营养摄入、行为和人类邻近性的变化可能对野生灵长类种群有重要的流行病学影响[44]。
当前,随着人类活动增加,大部分非人灵长类动物栖息生境的破碎化趋势加剧[89]。生境破碎化不仅影响生境数量和质量,还影响种间关系,其中包括动物和寄生虫的交互关系[90]。野生动物种群的寄生虫传播方式明显受到宿主特征(例如漫游方式、密度、种内和种间关系、食物)的影响[91]。然而,这些特征又随着生境破碎化而发生改变。生境破碎化导致生境面积减少,限制动物的漫游范围,增加群的聚集度和同种间的生境重叠,导致个体接触病原体的几率升高[88];种群密度的增加直接导致寄生虫多样性和感染率的升高[92];生境破碎化导致食物资源减少和质量下降,营养缺乏降低了动物的免疫力而更易受寄生虫感染[13]。同时,受破碎生境中隔离效应(isolation effects)影响,种群间个体的正常迁移受到限制,从而降低斑块间的基因流水平,导致物种遗传多样性降低[93],使它们更容易被寄生虫和病原体感染[13]。破碎生境的景观特征也可能影响野生动物、人类和牲畜间接触的频率和性质,增加病原体的传播潜能[94]。生境破碎形成不同大小的斑块,引起种群拥挤效应,其内部有限的觅食空间迫使动物更多地利用边缘生境,而这些区域存在更多人-牲畜共患的病原体/寄生虫。研究表明,与生活在连续适宜生境或生境斑块内部的灵长类种群相比,生活在破碎生境或边缘生境的种群具有更高的寄生虫感染率和感染更多种类的寄生虫[39-40]。
7 展望和结语
传染病已成为导致生物多样性丧失的一个重要威胁,尤其是肠道寄生虫引起的传染病往往在其中扮演重要角色。作为与非人灵长类动物密切相关的类群,肠道寄生虫对非人灵长类动物的生活甚至生存具有不可估量的影响。随着人类与非人灵长类动物接触的不断增加,人畜共患病原体在人类和非人灵长类动物之间传播的可能性也随之增加。目前人畜共患的新发疾病已成为人类面临的巨大挑战,研究并了解野生动物的肠道寄生虫和人畜共患肠道寄生虫非常必要,对人类公共卫生具有重要意义。未来研究方向主要有:
1)利用分子标记、寄生虫基因组计划等分子手段对非人灵长类动物的肠道寄生虫展开深入研究,并结合传统的形态学方法和分子手段对肠道寄生虫进行更深一步的分类和鉴定。
2)建立长期的非人灵长类动物的健康监测系统。
3)利用肠道寄生虫治疗疾病。肠道寄生虫与宿主已经进化出相互适应的关系,目前利用肠道寄生虫治疗疾病也刚刚开始探索,可以继续深入研究其相互适应的进化机制,并用于治疗一些自身免疫病,例如生产蠕虫性治疗药物。
4)探讨建立隔离区的可能性、方式和途径。在人类与非人灵长类动物密切接触的地区,可以建立相应隔离区,将人类和非人灵长类动物分隔开,减少人类与非人灵长类动物栖息地的资源共享,减少相互接触的机会,达到减少人畜共患肠道寄生虫传播的目的。
本文主要探讨了肠道寄生虫对非人灵长类动物各方面的影响,但肠道寄生虫与宿主的相互进化作用极其复杂,还需进一步探索,这对于评估和促进非人灵长类动物的健康非常重要。
