遮阴对濒危植物朝鲜崖柏光合特性的影响
2022-10-18苑景淇于忠亮兰雪涵李成宏田年军杜凤国
苑景淇,于忠亮,兰雪涵,李成宏,田年军,杜凤国,4*
(1. 北华大学林学院,吉林 吉林 132013;2. 吉林省林业科学研究院,吉林 长春 130033;3. 白山市林业科学研究院,吉林 白山 134300;4. 长白山特色森林资源保育与高效利用国家林业和草原局重点实验室,吉林 吉林 132013)
光是影响植物生存更新和生长发育的重要环境因子之一,不同生态习性的植物对光环境的适应策略不同。如阳性先锋树种具有高光饱和点和高光补偿点等特征,而阴性树种具有较低的光补偿点和较高的表观量子效率,以适应林下弱光环境[1]。近年来,通过光合生理生态研究进而分析濒危植物日趋减少的原因已经得到重视。如耐阴植物五桠果叶木姜子(Litseadilleniifolia)[2]和毛果木莲(Manglietiaventii)[3]受到森林持续片段化影响,强光严重阻碍林下幼苗生长,致使种群衰退;而林下光资源不足则成为喜光植物银缕梅(Parrotiasubaequalis)[4]和连香树(Cercidiphyllumjaponicum)[5]种群更新的主要限制因子。这些研究成果为濒危植物的就地/异地保护及繁衍等提供了理论依据。
朝鲜崖柏(Thujakoraiensis)为柏科(Cupressaceae)崖柏属常绿乔木或小乔木,是优良的用材和园林绿化树种,对探究古植物区系及柏科分类、系统发育具有重要学术研究价值[6]。朝鲜崖柏主要分布于吉林省长白山区海拔1 000~2 000 m、土壤贫瘠的山坡山谷等地,由于人类过度砍伐和生境破坏,导致野生朝鲜崖柏种群和个体数量急剧下降。到目前为止,对朝鲜崖柏的研究主要集中在群落特征[7-8]、繁育技术[9]、种子性状[10-11]、遗传变异[12]等方面,尚未见到关于朝鲜崖柏光合特性及环境因子对其光合作用影响的研究报道。学者野外调查发现,野生朝鲜崖柏幼苗幼树稀少且所处林分郁闭度较大,表明光照有可能是其更新的限制因子[1]。因此,本研究拟采用对朝鲜崖柏扦插苗人工遮阴的方法,探讨不同光环境下朝鲜崖柏光合作用与各环境因子的关系,旨在为这一珍稀濒危植物的有效保护提供科学依据。
1 材料与方法
1.1 试验材料
于2019年3月在吉林市龙潭区新山苗木基地的温室中选择植株大小相近、健壮无虫害的7年生朝鲜崖柏扦插苗(平均地径1.8 cm,平均株高48.43 cm,母树约50年生),室外种植于花盆中(花盆规格26 cm×30 cm,3/4埋入土壤,每盆1株),基质为草炭土和河沙(体积比3∶1),常规水肥管理。所有扦插苗在同等全光环境下适应30 d后,利用遮阴网进行人工遮阴,使用照度计设置4个光环境梯度的生境,即遮阴度分别为0%(全光照,L0)、25%(L1)、50%(L2)和75%(L3),每个光梯度5株扦插苗。待到2019年8月中旬(6:00—18:00日均温度30.14 ℃,日均湿度50.72%),进行光响应和日变化的测定。
1.2 光响应曲线测定
选晴朗无云稳定天气,在8:00—12:00选取朝鲜崖柏植株中部向阳新生叶片进行光响应曲线测定。光响应曲线所用光源为Li-6400配置的红蓝光LED光源,设定光辐射梯度为:2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、150、100、50、20、0 μmol/(m2·s),测定净光合速率(Pn)。根据叶子飘等[13]的直角双曲线修正模型拟合光响应曲线,得出光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(Rd)、最大净光合速率(Amax)以及表观量子效率(AQY)等参数。
1.3 光合日变化测定
光合日变化测定指标包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),以及光合有效辐射(PAR)、气温(Ta)、相对湿度(HR)、空气CO2浓度(Ca)等环境指标,并计算气孔限制值(Ls,Ls=1-Ci/Ca),测定时间分别为6:00、8:00、10:00、11:00、12:00、13:00、14:00、16:00和18:00,每个处理选定3盆进行测定,取平均值。
1.4 叶绿素含量测定
采用浸提法测定叶绿素含量。选取各环境梯度扦插苗植株中部向阳叶片清洗干净混匀,称取0.2 g置于试管中,加入25 mL乙醇丙酮(乙醇和丙酮体积比1∶1)混合液浸提24 h,将叶绿素色素提取液倒入光径1 cm的比色杯内,用UV-7504C紫外可见分光光度计在室温下测定440、645和663 nm波长下的吸光度,所得的光密度值代入叶绿素计算公式,根据唐延林等[14]的公式算出叶绿素和类胡萝卜素的含量(mg/g),测定3个重复值,结果取平均值。
1.5 数据处理与分析
采用Excel 2016和SPSS 19.0对数据进行统计分析,采用单因素方差分析法和LSD法进行方差分析和多重比较,应用通径分析计算统计各环境因子对Pn的直接和间接影响。应用Origin制图,图表中数据均采用平均值±标准误差表示。
2 结果与分析
2.1 主要环境因子日变化
测定当天光合有效辐射(PAR)和气温(Ta)日变化为早晚低、中午高的“单锋”曲线,在6:00—12:00逐渐升高达到最高值,分别为1 038 μmol/(m2·s)和34.36 ℃,然后下降,在18:00降至最低值,分别为291.55 μmol/(m2·s)和25.14 ℃。相对湿度(HR)在6:00时最大,为67.70%,此后随着Ta的升高而下降,在12:00时下降至最低值44.51%,之后逐渐升高。空气CO2浓度(Ca)和HR的变化趋势相似,在6:00时最高随后逐渐下降,13:00达到最低值后稍有回升(图1)。

图1 光合有效辐射(PAR)、空气CO2浓度(Ca)、气温(Ta)和相对湿度(HR)日变化Fig.1 The diurnal variation of photosynthetic active radiation(PAR), air CO2 concentration(Ca), air temperature(Ta)and relative humidity(HR)
2.2 光响应曲线及特征参数比较
不同光环境下朝鲜崖柏光合光响应曲线见图2。

图2 不同遮阴处理朝鲜崖柏光响应曲线Fig.2 The response photosynthetic curves of Thuja koraiensis under shading treatments
光响应曲线反映了植物净光合速率(Pn)随着光合有效辐射(PAR)改变的变化规律。不同遮阴处理朝鲜崖柏的净光合速率(Pn)对PAR的响应过程大致相同,当光合有效辐射(PAR)≤ 400 μmol/(m2·s)时,各处理Pn均随PAR的增加而迅速增加;当PAR升高至800 μmol/(m2·s)时,Pn增加速度减缓,逐渐趋于稳定;当PAR≥800 μmol/(m2·s)时,Pn略有下降,出现轻微的光抑制现象。
光响应曲线拟合特征参数结果(表1)表明,遮阴处理对各项数据影响显著(P<0.05)。其中,不同光环境下朝鲜崖柏的最大净光合速率(Pn,max)、暗呼吸速率(Rd)、光饱和点(LSP)和光补偿点(LCP)与遮阴度呈显著负相关,Pn,max、Rd、LSP和LCP数值在4种遮阴处理中由大到小的排序均为L0 > L1 > L2 > L3。而L1处理的表观量子效率(AQY)最高为0.046 mol/mol,分别是L0、L2和L3处理的1.24、1.27、1.77倍。

表1 不同遮阴处理朝鲜崖柏光响应曲线特征参数
2.3 不同遮阴处理朝鲜崖柏的光合日变化
2.3.1 净光合速率(Pn)和光合有效辐射(PAR)日变化
不同遮阴处理下朝鲜崖柏各时间点净光合速率(Pn)和光合有效辐射(PAR)日变化见图3。Pn的变化动态均为“双峰曲线”,Pn上午高于下午,在6:00时Pn最大。12:00时PAR为一天中最高时刻,此时Pn明显降低,朝鲜崖柏为“午休”状态。由于18:00时光照强度不足,Pn下降为负值。总体上看,不同光环境下朝鲜崖柏Pn和PAR值由大到小的排序均为L0 > L1 > L2 > L3,且在各处理间差异显著(表2)。

图3 不同遮阴处理朝鲜崖柏净光合速率(Pn)和光合有效辐射(PAR)的日变化Fig.3 The diurnal changes of Pn and PAR of Thuja koraiensis under different shading treatments

表2 不同遮阴处理朝鲜崖柏光合参数日平均值
2.3.2 气孔导度(Gs)和蒸腾速率(Tr)日变化
不同遮阴处理下朝鲜崖柏气孔导度(Gs)和蒸腾速率(Tr)日变化见图4。由图4可知,不同处理的Gs日变化几乎呈现相同变化趋势,即在6:00时最高,之后波动性变化,Gs变化范围在0.017~0.158 mmol/(m2·s) 之间。在10:00时Gs稍有降低,说明朝鲜崖柏不论遮阴与否在10:00时都会出现不同程度气孔关闭现象。气孔关闭势必影响朝鲜崖柏的蒸腾速率,Tr日变化曲线为单峰型,各处理均在12:00出现最高峰。遮阴度对蒸腾速率有重要影响,与全光照相比遮阴后Tr显著下降,且遮阴程度越高降低趋势越明显,与L0处理相比,L1、L2和L3处理的日平均蒸腾速率依次下降10.95%、21.54%和38.55%。

图4 不同遮阴处理朝鲜崖柏气孔导度(Gr)和蒸腾速率(Tr)的日变化Fig.4 The diurnal changes of Gr and Tr of Thuja koraiensis under different shading treatments
2.3.3 胞间CO2浓度(Ci)和气孔限制值(Ls)
不同遮阴处理下朝鲜崖柏胞间CO2浓度(Ci)和气孔限制值(Ls)日变化见图5。结合Pn日变化(图3)可以看出,各组处理Ci的变化趋势与Pn相反,其日变化曲线近似“W”形,且随遮阴程度增强,Ci逐渐增加,其中L0与L3差异显著(P< 0.05)。气孔限制值Ls与Ci的变化趋势相反,Ls日变化曲线呈现近似“M”形,但不同遮阴处理的第1个峰值出现时间略有不同,L0和L3在11:00,L1和L2出现在10:00。

图5 不同遮阴处理朝鲜崖柏胞间CO2浓度(Ci)和气孔限制值(Ls)的日变化Fig.5 The diurnal changes of Ci and Ls of Thuja koraiensis under different shading treatments
2.4 不同遮阴处理对朝鲜崖柏色素含量的影响
植物叶绿素含量与光合作用有密切联系。不同遮阴处理朝鲜崖柏的叶绿素含量存在显著差异(P< 0.05)。由表3可知,随遮阴度的升高朝鲜崖柏叶片中叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量逐渐增加,L3处理下最大。与L0相比,L3处理的叶片叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量分别增加64.44%、72.73%、67.16%和22.22%。L1处理叶绿素a/b值最大为2.110,随遮阴度增加叶绿素b合成幅度大于叶绿素a,使比值减小。L0处理类胡萝卜素含量最低为0.018 mg/g,遮阴处理L1、L2和L3相比于L0处理类胡萝卜素分别增加72.22%、61.11%和22.22%。

表3 不同遮阴处理朝鲜崖柏叶片色素含量及比值
2.5 不同遮阴处理朝鲜崖柏日变化净光合速率(Pn)与环境因子关系分析
2.5.1 相关分析
由不同遮阴处理下朝鲜崖柏日变化净光合速率(Pn)与环境因子Ta、Ca、HR、PAR之间的相关性检验结果(表4)可知,在L0、L1和L2处理中,Pn与HR呈极显著或显著正相关,但在L3处理中相关性不显著。在L2和L3处理中,Pn与PAR显著正相关。在L0处理中,Pn与Ca为极显著正相关。

表4 不同遮阴处理朝鲜崖柏日变化净光合速率(Pn)与各环境因子的相关性分析
2.5.2 多元线性回归分析
以Pn为因变量,分别以Ta、Ca、HR、PAR4个环境因子为自变量,进行逐步多元回归分析,得出Pn与各环境因子的回归方程见表5。由表5可知,回归模型均通过显著性检验。影响朝鲜崖柏Pn的主要环境因子因不同遮阴处理而存在差异,其中HR、Ta和Ca是L0和L1处理的主要环境影响因子,而L2和L3处理的主要环境影响因子为PAR和HR。

表5 不同遮阴处理朝鲜崖柏日变化净光合速率(Pn)与各环境因子的多元线性回归方程
2.5.3 通径分析
应用通径分析可定量分析各环境因子对Pn的影响,其中通径系数可以揭示环境因子对Pn的直接作用和间接作用,决定系数R2可将环境因子对Pn的综合作用进行排序。由通径分析结果(表6)可知,在不同遮阴处理下,比较各环境因子直接通径系数大小,在L0和L1处理中Ta直接通径系数分别达到0.899和1.101,远高于Ca和HR,直接作用最强,但Ta与Pn的相关系数较低(0.341和0.259),均未达到显著水平,可能是由于其直接作用被Ca和HR的间接副作用所掩盖;在L2和L3处理中直接通径系数均表现为PAR>HR。并且PAR与Pn相关系数随遮阴度增加,由L0和L1中的不显著,到L2和L3中为显著正相关,反映PAR对Pn直接影响作用逐渐增强。在L0和L1中决定系数均表现为HR>Ca>Ta,在全光照和轻度遮阴下,HR是影响Pn最重要的环境因子。随遮阴度增加,在L2和L3中决定系数值均表现为PAR>HR,说明PAR对Pn的综合作用最强,反映光照对朝鲜崖柏光合作用的重要性。
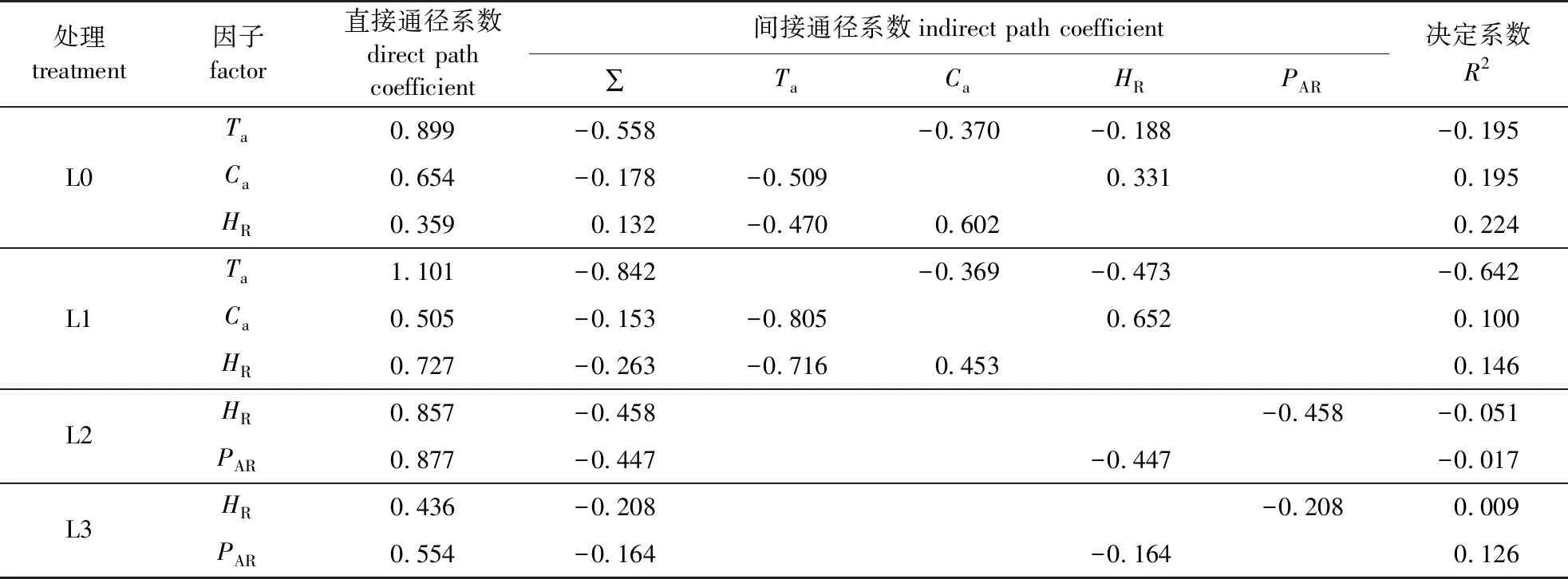
表6 不同遮阴处理朝鲜崖柏日变化净光合速率(Pn)与环境因子的通径分析
3 讨 论
光响应曲线对于了解植物光合作用过程中的光化学效率具有重要意义[15]。表观量子效率(AQY)反映了叶片在弱光情况下的光合能力,通常生长良好植物的表观量子效率为0.04~0.07 mol/mol[16]。本研究中L0、L1和L2处理下AQY处于或接近该区间,表明朝鲜崖柏具有一定的耐阴性,但L3处理未在该取值范围,反映在遮阴75%时,朝鲜崖柏叶片光合能力较弱,生长受到较大影响。有研究表明,阳生植物的光补偿点(LCP)一般在50~100 μmol/(m2·s)之间[17],植物在弱光条件下能通过降低光补偿点(LCP)和光饱和点(LSP)提高对弱光的捕获和利用能力,以促进合成和积累生长所需的能量[18]。本研究中,全光下朝鲜崖柏表现为阳生植物[LCP= 72 μmol/(m2·s)],而遮阴条件下表现为阴生植物[LCP< 50 μmol/(m2·s)]。相对L0,遮阴处理下LCP和LSP均不同程度降低,表明朝鲜崖柏对一定程度的弱光环境有较强的适应性,这与刘建锋等[1]对崖柏(Thujasutchuenensis)的研究结果一致。
光合作用日变化是植物生产过程中物质积累与生理代谢的基本单元,是分析环境因素影响植物生长和代谢的重要手段[19]。杨文娟等[20]研究表明,在环境条件和生理条件均达到最佳或较适宜的组合状态时,植物光合作用的Pn值最大。本研究中,不同光环境下朝鲜崖柏日变化净光合速率(Pn)均在6:00时为最高值,可能此时温湿度适宜,光照充足,气孔开启,经夜间呼吸作用后胞间CO2浓度较高,植物体内水分较充足,所以此时Pn值最大。光合“午休”是植物应对炎热环境做出的一种生态适应和自我调节反应,本研究中不同遮阴处理的朝鲜崖柏均在12:00时出现光合午休现象,且随遮阴程度增加Pn逐渐减小。引起叶片Pn降低的自身原因有气孔和非气孔因素两种,判断的依据是Pn和Ci同时降低时气孔限制值(Ls)增大为气孔因素,若Pn下降Ci上升时Ls下降为非气孔因素[21]。由表2可知,非气孔因素是朝鲜崖柏Pn下降的主要原因,朝鲜崖柏光合午休时光强和温度都达到一天中的最大值,表明朝鲜崖柏对高温、高光强耐受能力较弱,使叶肉细胞活性降低,导致光合能力下降。同时,随遮阴程度降低,Pn、Gs和Tr随之增大,体现了朝鲜崖柏的喜光性,这与赵顺等[22]对叉子圆柏(Juniperussabina)的研究结果一致。
相关研究表明,为适应弱光环境,植株可通过增加光合色素含量来提高对光能吸收利用的能力[23]。本研究中,遮阴处理下植株的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量显著增加,保证了在弱光环境中朝鲜崖柏能尽可能多地吸收和利用光能进行光合作用,缓解部分受光照不足的影响。同时,遮阴处理使叶片叶绿素a/b显著减小,说明叶绿素b的增长量相对更高。根据薛思雷等[24]的研究结果,叶绿素b相对含量的增高可提高对弱光的捕获能力。类胡萝卜素/总叶绿素的比值随遮阴程度增加呈先增加再减小的趋势,且在L1处理时最大,在L3处理时最小。而类胡萝卜素具有猝灭过剩光能的作用[25],说明相较于其他处理,朝鲜崖柏在L3弱光环境中对过剩光能淬灭能力会有所减弱,这是对光环境胁迫长期适应的体现。由此可见朝鲜崖柏通过对光合色素含量的调节增强了对弱光环境的适应,这与邓秀秀等[26]对马尾松(Pinusmassoniana)的研究结果一致。
逐步多元回归和通径分析结果表明,在全光照L0和轻度遮阴L1处理下,对朝鲜崖柏日变化净光合速率(Pn)直接作用最大的环境因子为Ta,随遮阴度增加的L2和L3处理中,HR对Pn的直接作用最大;而根据决定系数值大小排序结果,在L0和L1处理中HR影响最大,在L2和L3处理中PAR影响最大。本研究中,不同环境下湿度与温度均为极显著负相关(表4),湿度的变化直接影响温度,而温度是制约植物光合生产力的一个重要的环境因素[27]。温度制约着酶的活性,直接影响植物光合速率的关键酶Rubisco的最适温度为25~30 ℃[28-29]。本研究进行时正值夏季高温天气,白天日最高气温(34.36 ℃)和日均气温(30.18 ℃)均高于Rubisco酶的最适温度,可能使Robison对CO2的亲和性下降速度快于对O2的亲和性,导致光呼吸速率增高,光合速率降低。同时通径分析也显示,Ta对Pn的决定系数为负值,表明朝鲜崖柏适宜生长于凉爽的环境中,高温抑制了朝鲜崖柏的光合速率。植物光合作用能量的最终来源是光合有效辐射,光能不足则会限制光合作用[30]。本研究相关分析结果中,在L2和L3处理中Pn与PAR呈显著正相关,通径分析也显示,PAR对Pn的决定系数值最大,因此,在遮阴环境下PAR是影响Pn的决定环境因子,这与杨文娟等[20]对崖柏的研究结果一致。
综上所述,在全光照(L0)和遮阴度25%(L1)光环境下,朝鲜崖柏表现出较好的光合性能,相对湿度(HR)是影响光合作用的决策环境因子。随遮阴度增加(L2和L3),朝鲜崖柏通过降低净光合速率、蒸腾速率和气孔导度及增加胞间CO2浓度和叶片色素含量来适应遮阴环境,此时决策环境因子是光合有效辐射(PAR)。由文献[7]可知,野生朝鲜崖柏分布林分郁闭度为0.50~0.75,因此推测较高的郁闭度可能使野生朝鲜崖柏生存和繁衍受限。后续可对不同郁闭度下的野生朝鲜崖柏开展光合特性研究,并结合生长指标等测定进行长期观测,对于探讨朝鲜崖柏对光的响应和种群繁殖与复壮将更具实践意义。
