桃蛀螟CYP6AE76基因参与对Cry1Ab蛋白的解毒作用
2022-10-18静大鹏黄晓丹张天涛王振营
静大鹏, 黄晓丹,2, 张天涛, 王振营,*
(1. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193;2. 吉林农业大学生物防治研究所, 天敌昆虫应用技术工程研究中心, 长春 130118)
细胞色素P450单加氧酶(cytochrome P450 monooxygenase, CYP450)是广泛存在于真核生物体内的一类含有血红色素单链蛋白基因组成的一个超家族(Werck-Reichhart and Feyereisen, 1999),是多功能氧化酶系统中的重要组成部分,能够同时活化氧分子和与底物结合(Field and Devonshire, 1998),在昆虫体内参与了许多重要的生理过程(Coon, 2005)。CYP450氧化酶可参与杀虫剂和植物次生物质的代谢(Guengerichetal., 2001),并可以降解杀虫剂,提高昆虫的抗药性(Bagchietal., 2016)。根据序列同源性和相似程度,CYP450基因可以分为许多不同家族,但CYP6, CYP9, CYP12, CYP18和CYP28这5个基因家族为昆虫所特有(Bergéetal., 1998)。而在昆虫中已克隆鉴定出的序列中,CYP4和CYP6基因家族成员占到了总数的60%以上,且这两个家族基因的过量表达被认为是与其抗药性相关(郭亭亭等, 2009)。随着研究的深入,更多的研究发现CYP6基因在介导植物与昆虫的相互作用中发挥了重要作用(Helvig, 2004; Jonesetal., 2011)。具体表现在CYP6家族基因可以降解和代谢多种植物内源性化合物以及杀虫剂(车燕飞等, 2014),这对于多食性的害虫而言就显得更加重要(Lietal., 2007; Mittapellyetal., 2019)。
桃蛀螟Conogethespunctiferalis作为一种多食性的害虫,其寄主十分广泛,是我国多种果树、经济作物以及玉米等农作物上的重要害虫(王振营等, 2006; 鹿金秋等, 2010)。迄今为止,化学杀虫剂仍然是防治桃蛀螟最普遍的方法,但同时也对非靶标害虫、有益生物、人类健康和环境产生了负面影响。而转苏云金杆菌Bacillusthuringiensis(Bt)杀虫基因玉米的商业化应用,为防治桃蛀螟防治提供了新策略。目前,虽已有很多研究报道了CYP6家族基因具有降解或者代谢杀虫剂功能,但至今未有CYP6家族基因具有降解和代谢Bt蛋白功能的相关报道。而前期通过对多食性害虫桃蛀螟和寡食性害虫松蛀螟C.pinicolalis幼虫转录组测序,筛选出在桃蛀螟中差异表达的CYP6AE76,此基因可能在桃蛀螟取食多种寄主植物的解毒过程中发挥了重要作用。基于此,本研究将通过RT-qPCR、RNAi和人工毒力测定等方法研究该基因在桃蛀螟幼虫中对Cry1Ab蛋白的解毒功能,为多食性害虫的防控提供一定的科学依据,同时也为研究CYP6家族基因与Bt家族中的Cry1Ab蛋白的关系提供一定数据支持。
1 材料与方法
1.1 供试昆虫
实验所用的桃蛀螟采自中国农业科学院植物保护研究所廊坊基地玉米雌穗中的幼虫,在室内温度27±1℃、相对湿度70%~80%、光周期16L∶8D的条件下采用人工饲料进行多代连续饲喂(Jingetal., 2021)。
1.2 CYP6AE76基因序列鉴定与分析
基于前期测得桃蛀螟幼虫转录组数据(SRR12989228-SRR12989230)中筛选得到的CYP6AE76序列,并将序列使用NCBI在线BLASTP检索NR库后确认序列的匹配程度及找到其开放阅读框。其次,采用ExPASy在线网站(https:∥web.expasy.org/protparam/)预测CYP6AE76基因所编码蛋白的理化性质。
1.3 CYP6AE76基因在桃蛀螟幼虫中的表达谱分析
分别收集桃蛀螟1龄幼虫(50头)、2龄幼虫(30头)、3龄幼虫(10头)、4和5龄幼虫(各5头),为1个生物学重复,收集4龄幼虫头(5头)、中肠(10头)、血淋巴(20头)和脂肪体(10头),为1个生物学重复,采用Trizol法(Invitrogen)进行RNA提取,随后将得到的RNA采用1%的琼脂糖凝胶电泳检测完整性并用核酸分析仪(NanoDrop 2000)检测RNA质量及浓度。采用全式金生物有限公司的TranScript One-Step gDNA Removal and cDNA Synthesis SuperMix(AT311)试剂盒并参照其说明书反转录合成cDNA。RT-qPCR参照PrefectStartTMGreen qPCR SuperMix试剂盒(北京全式金生物技术有限公司,北京)说明书进行。PCR反应体系: PrefectStartTMGreen qPCR SuperMix 1 μL, 上下游引物(10 μmol/L)各0.4 μL, 50×ROX Reference Dye 0.4 μL, cDNA 模板1 μL, Nase-free Water 6.8 μL。扩增程序: 94℃ 30 s; 94℃ 5 s, 60℃ 30 s, 共40个循环。引物采用Primer Premier 5.0软件设计,同时以RP49(杨苓等, 2017)为内参基因(表1)。实验设置3个生物学重复,每个重复中进行4次技术重复。
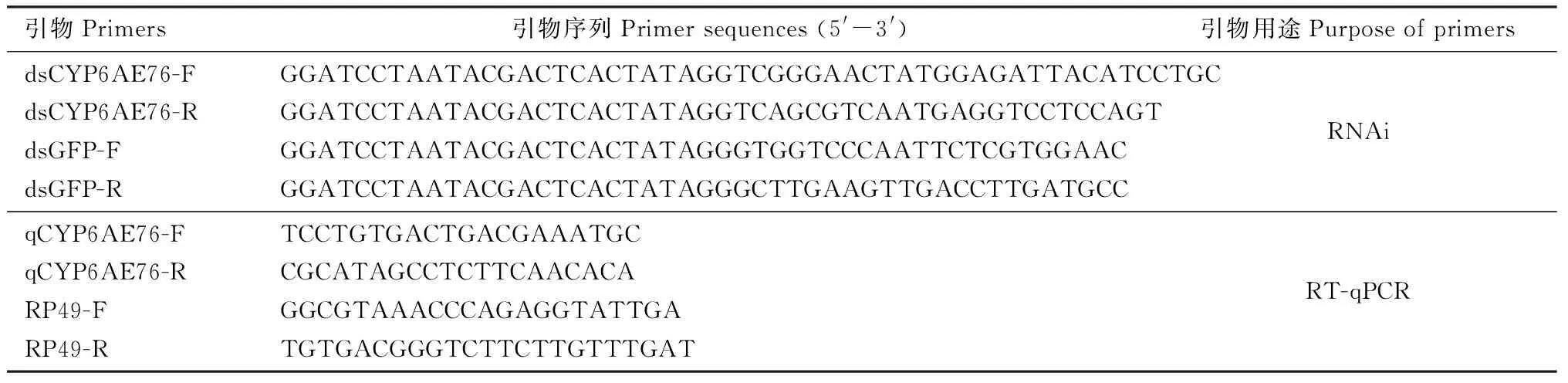
表1 引物信息Table 1 Primer information
1.4 幼虫取食Cry1Ab蛋白对CYP6AE76基因表达的影响
将4龄桃蛀螟幼虫转接到含有Cry1Ab蛋白(LC50=1.08 ng/cm2)人工饲料的24孔板中进行饲喂,每孔放1头幼虫。同时,以未加Cry1Ab蛋白人工饲料饲养的幼虫为对照,3 d后取存活的5头幼虫的中肠和20头幼虫的血淋巴为1个生物学重复,并提取总RNA,采用全式金生物有限公司的TranScript One-Step gDNA Removal and cDNA Synthesis SuperMix(AT311)试剂盒并参照其说明书反转录合成cDNA,利用RT-qPCR检测CYP6AE76基因的表达水平(方法同1.3节)。实验进行3次生物学重复。
1.5 RNAi和干扰效果检测
基于CYP6AE76的功能区序列设计特异性引物dsCYP6AE76-F和dsCYP6AE76-R,同时根据GFP(GenBank登录号: AB062168.1)序列设计特异性引物dsGFP-F和dsGFP-R作为对照(表1)。dsRNA的合成方法参考T7 Ribo MaxTMExpress RNAi System 试剂盒说明书(Promega)进行,最后dsRNA浓度调整为1 000 ng/μL备用。将人工饲料切成约0.3 cm3的小块,分别放入24孔板中;随后在饲料表面滴入2 μL的dsRNA, 空白对照以滴入无RNA酶的水替换dsRNA。取饥饿6 h桃蛀螟4龄幼虫分别放入饲料表面让其取食,并分别在幼虫取食后不同时间段(0, 24, 48, 72, 96和120 h)取5头幼虫的中肠进行RNA提取并合成cDNA,采用RT-qPCR对基因干扰效率进行检测(方法同1.3节)。此外,为检验干扰CYP6AE76是否影响幼虫的生长发育,分别测定了取食和未取食含有dsRNA饲料的幼虫120 h后的存活个数并计算存活率及对存活幼虫体重进称重并分析。
1.6 人工毒力测定
将初孵幼虫转接到分别含有dsCYP6AE76和dsGFP(阴性对照)的饲料中(方法同1.5节),并以未进行dsRNA处理的初孵幼虫作空白对照,饲喂2 d后再转接到含有Cry1Ab蛋白(浓度为LC50=1.08 ng/cm2)的人工饲料中进行饲喂(方法同1.4节),7 d后调查幼虫成活情况并计算存活率,同时对存活幼虫进行称重并分析。实验进行3次生物学重复。
1.7 数据分析
所有数据均采用SPSS软件进行独立样本t检验或单因素方差分析(one-way ANOVA)与LSD法进行统计学分析和比较。
2 结果
2.1 CYP6AE76的cDNA和氨基酸序列
CYP6AE76基因开放阅读框长1 572 bp,经过序列比对分析后发现桃蛀螟CYP6AE76与稻纵卷叶螟Cnaphalocrocismedinalis的CYP6AE76(GenBank登录号: CBB07053)的核苷酸序列一致性为62.72%,同源性最高,且该基因的开放性阅读框编码524个氨基酸,预测分子量大小为60.34 kD(图1)。此外,与稻纵卷叶螟、烟草粉螟EphestiaelutellaCYP6AE76(GenBank登录号: QZP43552)、东方粘虫MythimnaseparataCYP6AE76(GenBank登录号: QOI60672)和草地贪夜蛾SpodopterafrugiperdaCYP6AE76(GenBank登录号: QGA73297)的氨基酸序列对比分析后,发现几个物种均有CYP6A基因的典型特征序列:FxxGxxxCxG, ExxR和PxRF,且保守性较高。
2.2 CYP6AE76的表达谱
结果表明CYP6AE76在桃蛀螟不同龄期幼虫中均有表达,但在1龄幼虫中表达量最高(P<0.05),并随着幼虫龄期的增长,其表达量逐渐降低(图2: A)。在4龄幼虫不同组织中,CYP6AE76在中肠中相对表达量最高(P<0.05),其次是在血淋巴、脂肪体和头部(图2: B)。
2.3 幼虫取食Cry1Ab后对CYP6AE76表达的影响
桃蛀螟4龄幼虫取食含有Cry1Ab蛋白的饲料后,在蛋白LC50=1.08 ng/cm2浓度下会有约半数的幼虫死亡,但在存活个体中CYP6AE76相比对照,其表达量在血淋巴和中肠中显著上调(P<0.05)(图3),特别是在中肠的表达量变化最大。
2.4 CYP6AE76的RNAi干扰效果
与取食含有无RNase水的饲料(空白对照)和含dsGFP(阴性对照)的比较,桃蛀螟4龄幼虫取食含有dsCYP6AE76的饲料后,其CYP6AE76基因表达量在24 h时显著下调(P<0.05)(图4: A),说明合成的dsCYP6AE76可以有效抑制桃蛀螟幼虫CYP6AE76的表达量。 但随着时间推移, 其基因表达量又逐渐上升,并在处理后120 h与两对照组的无显著差异(图4: A)。而幼虫取食dsGFP处理后的饲料后,体内CYP6AE76基因表达量与饲喂水的空白对照相比未发生显著变化(P>0.05)。此外,幼虫取食含有dsCYP6AE76和dsGFP的人工饲料后,幼虫存活率(图4: B)和平均体重 (图4: C)与两对照组相比均无显著差异(P>0.05)。
2.5 RNAi干扰CYP6AE76对桃蛀螟幼虫Cry1Ab 蛋白解毒能力的影响
当桃蛀螟初孵幼虫取食含有dsCYP6AE76和dsGFP(对照)的饲料后,再取食含有Cry1Ab 蛋白的人工饲料,其存活率与对照相比虽有下降,但差异未达到显著水平(P>0.05)(图5: A),但平均体重与对照相比极显著下降(P<0.01)(图5: B)。而结合2.4节中结果,仅干扰CYP6AE76基因饲喂不含有Cry1Ab蛋白的人工饲料,并不会影响桃蛀螟幼虫的存活率和体重。综上所述,说明当CYP6AE76基因受到干扰时,桃蛀螟幼虫取食含有Cry1Ab的人工饲料后,其体重增长会受到明显的抑制。
3 讨论
本研究基于桃蛀螟幼虫转录组数据,通过同源性搜索对比获得并初步解析了桃蛀螟CYP6AE76的功能。CYP450家族基因在昆虫体内表达和分布具有多样性(张雅男, 2019)。袁星星等(2021)通过对桃蛀螟体内CYP4G113的研究后发现,该基因在桃蛀螟各发育阶段均有表达,其中1龄幼虫期表达量最高,5龄幼虫中表达量最低,这一结果与本研究中桃蛀螟CYP6AE76基因在幼虫各个龄期表达结果(图2)一致。其原因可能是桃蛀螟初孵幼虫需要CYP6AE76或者CYP4G113基因高度表达以迅速适应寄主植物中的次生代谢物质,以建立起代谢或者解毒功能的应激式的反应。但随着后期龄期的增长,幼虫逐渐对这些次生代谢物质不再敏感,故其表达量会逐渐降低。其次,研究发现CYP6家族基因在中肠组织中高度表达,这主要与其解毒功能密切相关(Krempletal., 2016; Shietal., 2018)。本研究中,CYP6AE76在桃蛀螟中肠中高度表达(图2),而中肠作为幼虫食物消化代谢的主要场所,也是其发挥解毒功能的主要场所,Cry类Bt蛋白主要也是通过与中肠上的受体结合产生杀虫功能。此外,稻纵卷叶螟的CYP6AE76基因在脂肪体和马氏管中高度表达,这被推测该基因在调节发育和衰老中也发挥了作用(Lietal., 2014; Liuetal., 2014),具体功能还需深入探究。
CYP6家族基因在昆虫体内可参与杀虫剂抗性,其表现为抗性品系中CYP6基因或蛋白质表达水平增加,从而使CYP6的解毒作用增强(Feyereisenetal., 2015; Modi and Dawson, 2015; Wangetal., 2018),但目前未有CYP6家族基因可以参与代谢体内Bt蛋白的相关报道。本研究中,桃蛀螟幼虫取食含有Cry1Ab蛋白的人工饲料后,其血淋巴和中肠中的CYP6AE76会显著上调(图3),说明幼虫取食Cry1Ab蛋白后可以导致体内CYP6AE76基因表达量上调。RNAi技术用于沉默该基因的表达,实验表明沉默CYP6AE76基因不会对桃蛀螟幼虫的存活率(图4: B)和体重(图4: C)产生显著影响,且使用dsCYP6AE76基因进行干扰,能够有效抑制幼虫体内CYP6AE76基因的表达量(图4: A),而引入外源的GFP基因对幼虫的存活率和体重无影响(图4: B, C)。此外,当沉默CYP6AE76基因的表达量后,发现桃蛀螟初孵幼虫取食Cry1Ab蛋白后的存活率并未有显著降低(图5: A),但是体重却显著受到抑制(图5: B)。这可能是由于CYP6AE76基因的下调导致了其体内解毒功能的下降,影响了幼虫对Cry1Ab蛋白的解毒功能,进而影响幼虫取食并导致体重显著下降。
昆虫中肠上能够结合Cry1Ab蛋白的特异性受体主要有氨肽酶N(aminopeptidases N, APN)、钙黏蛋白(cadherin, CAD)、碱性磷酸酯酶(alkaline phosphatase, ALP)和腺苷三磷酸结合盒转运蛋白(ATP-binding cassette transporter, ABC转运蛋白)(Jurat-Fuentesetal., 2021),而Cry1Ab蛋白与这些特异性受体结合后会形成晶体破坏幼虫中肠细胞并引发败血症而死亡(Brodericketal., 2006; Masonetal., 2011)。因此,特异性受体与Cry1Ab蛋白的结合能力是决定桃蛀螟幼虫的存活率的一个重要原因,而CYP6AE76仅是幼虫体内的解毒基因之一,故仅干扰CYP6AE76不能显著影响幼虫取食Cry1Ab蛋白后的存活率。但Cry1Ab蛋白进入昆虫中肠后,作为重要的解毒蛋白,也会发挥其解毒功能,故在LC50浓度Cry1Ab蛋白作用下,有一半的幼虫得以存活。但当干扰CYP6AE76基因的表达时,相较于对照,桃蛀螟幼虫的体重明显降低(图5: B),说明该基因的解毒功能也受到了抑制,从而间接影响了幼虫的取食,导致其体重降低。
本研究报道了桃蛀螟CYP6AE76基因对Cry1Ab蛋白有解毒作用,表明该基因可作为其幼虫在代谢Cry1Ab蛋白中的一个关键作用的基因,可抑制幼虫的取食和生长,以减少幼虫为害。这对于监测桃蛀螟幼虫对Cry1Ab的抗性以及未来使用或研发新的化学制剂来防控其敏感或抗性种群将至关重要。
