西伯利亚蝗卵发育过程的比较转录组分析及滞育关联基因筛选
2022-10-18吕雪峰扈鸿霞蒋思涵叶小芳
赵 娜, 吕雪峰, 扈鸿霞, 宋 余, 蒋思涵, 季 荣, 叶小芳,*
(1. 新疆师范大学生命科学学院, 中亚区域跨境有害生物联合控制国际研究中心,新疆特殊环境物种保护与调控生物学实验室, 乌鲁木齐 830054;2. 新疆畜牧科学院畜牧业质量标准研究所, 乌鲁木齐830000)
滞育是动物受到环境条件诱导所产生的一种静止状态,可发生在昆虫的卵、幼虫、蛹和成虫任意发育阶段。且滞育可提高昆虫的越冬存活率,帮助度过不良环境以维持种群发育一致(黄少虹, 2009)。目前有关昆虫滞育的研究主要集中在昼夜节律、光周期、激素调节以及分子调控等方面,但由于滞育的生理过程非常复杂,且物种和滞育虫态不同,滞育机制差别很大,涉及多个通路和基因的表达调控,因此现有研究对于阐明昆虫滞育机制远远不够(Wuetal., 2021)。
近年来,昆虫滞育状态下相关差异表达基因(differentially expressed genes, DEGs)和蛋白以及比较分析滞育相关基因在滞育与非滞育个体中的表达动态的研究报道较多(Zhangetal., 2011; Colinetetal., 2012; Haoetal., 2012; 齐晓阳等, 2016)。研究发现,滞育期间昆虫发育受阻,代谢通路受到抑制,会引发一种独特的基因表达模式(Gongetal., 2013)。通过比较飞蝗Locustamigratoria滞育卵与非滞育卵在转录和翻译水平上的差异,发现212个差异表达蛋白,卵滞育相关基因有Gst和JHE等(Tuetal., 2015)。飞蝗成虫卵巢和脂肪体转录组共富集到137个DEGs,其中调控滞育的71个DEGs参与Foxo信号通路(Haoetal., 2019)。伞裙追寄蝇Exoristacivilis的转录组测序结果表明滞育相关基因与抗寒、氨基酸代谢和能量代谢通路有关(张博等, 2020)。
西伯利亚蝗Gomphocerussibiricus属直翅目(Orthoptera)蝗科(Acrididae),是新疆草原的一种主要优势害虫,严重破坏当地草场生态,且对农牧业经济影响较大;其卵期持续近10个月,面对新疆严寒冬季,蝗卵可通过进入滞育来抵御寒冷,且滞育维持时间长达5个多月(闫蒙云等, 2018; 赵娜等, 2021),因此,滞育作为西伯利亚蝗生活史中的一个重要生理阶段,对蝗卵的顺利孵化和胚后的个体发育及种群数量至关重要。目前,有关西伯利亚蝗的研究主要集中在成虫的呼吸代谢水平、呼吸蛋白表达、酶活性、抗逆物质、色素沉着等方面(李娟等, 2014; 李爽等, 2016; 钱雪等, 2016, 2017; Shahetal., 2019),而有关该物种的滞育研究较少,仅限于蝗卵的滞育形态描述、发育及滞育过程的血蓝蛋白表达(何立志等, 2017; 闫蒙云等, 2018; 赵娜等, 2021)。究其原因,主要是因为该物种的基因序列信息缺乏,严重阻碍了其滞育机理研究。为弄清西伯利亚蝗卵的滞育分子机理,本研究采用高通量测序技术对不同发育阶段蝗卵进行转录组测序,比较分析蝗卵滞育期和非滞育期的转录组数据,筛选蝗卵在滞育期间特异性表达或差异表达的滞育关联基因,可为该物种的基因组研究提供有用信息资源,且有助于阐明蝗卵滞育的分子调控机理,为进一步揭示其发生机制以及实现人工滞育调控提供理论依据。
1 材料与方法
1.1 供试蝗虫
西伯利亚蝗卵于2018年6-8月采于新疆哈密巴里坤县白石头乡的土壤中5 cm处。将采集的西伯利亚蝗雌雄成虫放置于采集地室外养虫笼中,用当地优势作物天山赖草Leymustianschanicus和针茅Stipacapillata进行饲喂,并放置装有土壤的花盆以供雌虫产卵。每日定时收集新鲜蝗卵,并将蝗卵埋于室外5 cm深土壤处让其自然发育,以供后续实验使用。参照闫蒙云等(2018)和王香香等(2019)的方法,每5~8 d取西伯利亚蝗卵进行解剖,观察胚胎发育形态。收集早期发育阶段(early developmental stage, ES)、滞育阶段(diapause stage, DS)和滞育后发育阶段(post-diapause developmental stage, PS)的蝗卵,储存于-80℃超低温冰箱(三洋MDF-382E,日本)中,每个阶段分为3组,每组100粒。
1.2 转录组测序、组装及基因功能注释和分类
采用Trizol Reagent(Invitrogen,美国)提取1.1节3个阶段西伯利亚蝗卵总RNA,检测合格后进行文库构建,cDNA文库构建与测序由北京诺禾致源生物信息科技有限公司协助完成。测序后,将得到的原始数据进行筛选、组装。使用BLAST软件(将Unigenes与KEGG, Nr, Nt, Pfam, KOG, Swiss-Prot以及GO数据库比对,获得西伯利亚蝗卵基因的序列信息及功能信息。
1.3 DEGs筛选及功能注释
参考序列为Trinity软件(v2.4.0)拼接得到的转录本序列,基因表达水平采用FPKM计算。利用DEGseq2确定滞育与非滞育蝗卵间基因表达水平的差异,筛选DSvsES和PSvsDS两比较组的DEGs条件为padj<0.05且|log2fold change|>1。
GO注释使用Blast2GO软件(version 2.5)(https:∥www.geneontology.org/)进行分析。经校正q值<0.05的GO条目被认为在DEGs中显著富集。参考KEGG数据库(http:∥www.genome.jp/kegg/),使用KAAS软件(r140224)进行KEGG通路注释。经校正q值<0.05的KEGG通路被认为在DEGs中显著富集。
1.4 滞育关联基因筛选
根据DSvsES和PSvsDS两比较组转录组测序结果、GO和KEGG富集分析结果并结合已报道的滞育相关通路,选取Wnt信号通路、胰岛素信号通路、Foxo信号通路、细胞周期以及昆虫激素生物合成通路进行聚类热图分析,预测以上通路富集的DEGs在ES, DS和PS阶段的表达模式,从而筛选滞育关联基因。
1.5 DEGs定量验证
从1.4节筛选出的20个滞育关联基因中选取6个重要基因进行 qRT-PCR验证,包括热激蛋白70kDa(heat shock 70kDa protein, HSPA1 s)基因、海藻糖酶 (trehalase, TREH)基因、海藻糖转运蛋白 (facilitated trehalose transporter1, TRET1)基因、钙调蛋白(calmodulin, CALM)基因、细胞分裂周期14 (cell division cycle 14, CDC14)基因和WNT6(wingless-type MMTV integration site family, member 6)基因,β-actin为内参基因。使用Primer Premier 5.0设计引物,引物序列见表1,并由生工生物工程(上海)有限公司合成。采用TB Green®Premix Ex TaqTMⅡ(Takara, 日本)进行qRT-PCR实验。反应体系(20 μL): TB Green Premix Ex Taq GC10 μL, 上下游引物 (10 μmol/L) 各0.8 μL, ES, DS和PS阶段西伯利亚蝗卵cDNA模板2 μL, RNase-free ddH2O 6.4 μL。反应程序(WNT6除外): 95℃ 30 s(4.4℃/s); 95℃ 5 s(4.4℃/s), 60℃ 30 s(2.2℃/s), 72℃ 15 s, 40个循环; 95℃ 5 s (4.4℃/s),60℃ 1 min(2.2℃/s), 95℃ 1 s(0.11℃/s),Acquisition Mode: Continuous;降温50℃, 30 s(2.2℃/s)。WNT6反应程序: 95℃ 30 s(4.4℃/s); 95℃ 5 s(4.4℃/s), 65℃ 30 s(2.2℃/s), 72℃ 15 s, 40个循环; 95℃ 5 s(4.4℃/s),60℃ 1 min(2.2℃/s), 95℃ 1 s(0.11℃/s),Acquisition Mode: Continuous;降温50℃ 30 s(2.2℃/s)。每组重复3次,结果采用2-ΔΔCt计算基因相对表达量。

表1 引物信息Table 1 Primer information
2 结果
2.1 不同发育阶段西伯利亚蝗卵的转录组
3个发育阶段的西伯利亚蝗卵转录组数据经过滤后,平均剩余97.97%的高质量clean reads。每个样本Q20均大于97%,Q30均大于93%,说明测序质量可靠。对转录本进行聚类和组装分析,得到176 081条unigenes,其拼接长度范围在301~43 324 bp,平均长909 bp,N50为1 221 bp。unigene数量随其组装长度增加呈平缓的下降趋势,说明原始数据拼接质量较高,可用于后续功能注释和DEGs分析;转录组原始数据已上传NCBI(GenBank登录号: SRP239011)。此外,对ES, DS和PS 3个发育阶段共9个样本的基因表达水平进行聚类分析(图1: A)及相关性分析(图1: B),结果证明样本间相似性较高,重复性好,组间样本差异性较好。
2.2 转录组序列注释
通过对获得的unigene序列与七大数据库比对分析发现:Nr数据库中注释最多,占33.25%,其后依次是Pfam(占27.72%), GO(占27.72%),Swiss-Prot(占18.85%),Nt(占13.23%),KOG(占8.24%)和KEGG (占2.46%)。
GO注释分类发现,所有unigene被注释在55个功能类别中(图2)。在生物学过程分类中,注释基因最多的功能是细胞过程(cellular process)、代谢过程(metabolic process)和单一生物过程(single-organism process),三者均参与细胞增殖和组织器官发育。在细胞分组分类中,注释基因最多的功能是细胞(cell)、细胞部分(cell part)和膜(membrane)。在分子功能分类中,注释基因最多的功能是结合(binding)、催化活性(catalytic activity)和转运活性(transporter activity)。
KEGG分析将unigene分为5大类——细胞过程(cellular processes)(A)、环境信息加工(environmental information processing)(B)、遗传信息加工(genetic information processing)(C)、新陈代谢(metabolism)(D)和生物体系统(organismal systems)(E),共注释到32条通路。通过对32条通路下的unigene进一步分析发现,其中翻译(translation)、信号转导(signal transduction)、碳水化合物代谢(carbohydrate metabolism)通路中涉及的unigene最多(图3)。
2.3 DEGs筛选
不同比较组DEGs筛选结果显示:DSvsES组和PSvsDS组分别有12 419和4 789个DEGs,且这2比较组间共表达的DEGs有2 206个,在上述相应时期特异性表达的DEGs分别有10 213和2 583个(图4: A)。DSvsES组DEGs中上调基因6 923个、下调基因5 496个;PSvsDS组的DEGs较少,其中上调基因2 612个、下调基因2 177个。分析发现,两比较组的DEGs均上调表达居多(图4: B)。
2.4 DEGs的GO功能注释
DSvsES组中(图5: A), DEGs富集的分子功能占比较大,有14条;细胞组分分类有5条;生物学过程分类中仅有1条,被注释为细胞骨架组织(cytoskeleton organization)。第2圈中颜色越红,q值越小,富集越显著。其中,富集最显著的GO条目为GO:0005515,被注释为蛋白结合(protein binding);其次为GO:0003779, GO:0008092和GO:0007010,分别被注释为肌动蛋白结合(actin binding)、细胞骨架蛋白结合(cytoskeletal protein binding)、细胞骨架组织,这都对胚胎发育具有重要作用。第3圈表示该GO条目中上调、下调的DEGs数量,其中,DSvsES组的GO条目中基因上调表达居多,且GO:0008076, GO:0034705, GO:0005249, GO:0005267和GO:0022836等条目中上调基因所占比例较大,经注释发现其均与通道活动和通道复合物相关。
由图5(B)可知,PSvsDS组中,DEGs富集的前20条GO条目中分子功能所占比例较大,细胞组分次之,生物学过程最少。第2圈富集显著性表明,PSvsDS组中富集最显著的功能条目是GO:0042302,被注释为表皮结构组成(structural constituent of cuticle);其次是GO:0015926, GO:0004564和GO:0090599,分别被注释为葡糖苷酶活性(glucosidase activity)、β-呋喃果糖苷酶活性(beta-fructofuranosidase activity)和α-葡萄糖苷酶活性(alpha-glucosidase activity)。根据第3圈DEGs上调、下调的DEGs数量可以看出,PSvsDS组中上调和下调的基因数总和基本一致。此外,除富集显著的前4条GO条目外,还富集到与胚胎发育有关的GO条目(GO:0007423, GO:0015629和GO:0003779),分别被注释为感觉器官发育(sensory organ development)、肌动蛋白细胞骨架(actin cytoskeleton)和肌动蛋白结合。PSvsDS组中DEGs富集的GO条目主要包含酶活性、细胞骨架构建及蛋白结合等过程。
2.5 DEGs的KEGG富集
DSvsES组(图6: A),富集最显著的前20条通路中,前3条通路分别是轴突导向(axon guidance)、Wnt信号通路(Wnt signaling pathway)和癌症通路(pathways in cancer),此外,还富集到与生长发育相关的通路,如脂肪细胞脂肪分解调控(regulation of lipolysis in adipocyte)、细胞粘附分子(cell adhesion molecules, CAMs)、甲状腺激素信号通路(thyroid hormone signaling pathway)、调节干细胞多能性的信号通路(signaling pathways regulating pluripotency of stem cells)、甘油磷脂新陈代谢(glycerophospholipid metabolism)和血管内皮生长因子信号通路(VEGF signaling pathway)等。富集到的DEGs主要有GSK3B,PKA,PLCB,WNT7,WNT6,CSNK1E,WIF1,AKT,HSPA1s和NOS1等。
PSvsDS组中(图6: B),除与人类疾病相关的麻疹(measles)通路外,DEGs富集最显著的通路是抗原加工提呈(antigen processing and presentation)、MAPK信号通路(MAPK signaling pathway)和吞噬体(phagosome)。此外,还富集到肌动蛋白细胞骨架调控(regulation of actin cytoskeleton)、调节干细胞多能性信号通路(signaling pathways regulating pluripotency of stem cells)、癌症通路(pathways in cancer)、趋化因子信号通路(chemokine signaling pathway)、胰岛素信号通路(insulin signaling pathway)等,富集到的DEGs主要有IFI30,CTSL,NLK,MKP,TUBB,TUBA,LIMK1,VAV,MYL2,WNT11,JNK,PRKCZ,FBP,AKT和FASN等。
2.6 滞育关联基因
结果显示,Wnt信号通路中富集的DEGs有20个(图7: A),除PRKCA外,其余基因表达量均在DS阶段呈上调表达。WNT6,NLK,RVB1,JNK和GSK3B在ES, DS和PS 3个阶段表达量整体呈下降-上升-下降的表达趋势,WNT11表达量在DS阶段也显著上调。PRKCA,CNB,WISP1,RHOA,WNT7和SMAD2_3的表达模式整体呈上升趋势,在PS阶段表达量最高。此外,WIF1与SFRP5作为Wnt信号通路的抑制基因,SFRP5表达量逐渐增加;WIF1表达量在DS阶段表达量最高,PS阶段虽仍呈上调表达,但表达量较DS阶段有所下降。
胰岛素信号通路在3个阶段中富集到15个DEGs(图7: B)。在西伯利亚蝗卵整个发育过程中FASN,AKT,MNK,JNK,GSK3B和FBP这6个基因表达量明显呈先升后降的趋势,在DS阶段表达量最高。CALM和KRAS在ES至PS阶段表达量上调,在PS阶段达到最高。PKLR这一丙酮酸激酶基因在ES阶段表达量最高,而在DS阶段表达量最低。
Foxo信号通路所富集的DEGs有10个(图7: C),其中NLK,AKT和JNK的表达量在整个发育过程呈上升-下降的表达趋势,在DS阶段最高。仅富集到1个F-box蛋白家族基因为FBXO25_32,其表达量在整个发育过程逐渐升高,在PS阶段达到最大值;与FBXO25_32表达趋势相同的基因还有PLK2,PIK3R1_2_3和SMAD2_3。SIRT1则在ES阶段表达量上调,随后逐渐降低。
细胞周期通路中共有7个DEGs(图7: D),分别是GSK3B,MPS1,MCM2,CDH1,SMAD2_3,SCC3和CDC14,且较ES阶段均上调表达。GSK3B,MPS1,MCM2和CDC14在DS阶段表达量较高,在DS至PS阶段表达量下降。
昆虫激素生物合成通路中仅富集到2个DEGs (SAD和SPO)(图7: E)。在ES至DS阶段,SAD与SPO均呈上调表达,其中SPO表达量较高;在DS至PS阶段,两者表达量均有所降低。
综合GO功能、KEGG富集分析以及部分通路的聚类热图分析结果,筛选出20个重要的滞育关联基因(表2)。
2.7 qRT-PCR验证
为验证转录组结果的准确性,根据DEGs的GO功能和KEGG富集分析结果,并结合相关文献报道,从20个滞育关联基因中选取与糖代谢、环境胁迫、生长发育以及细胞周期进程相关的6个基因进行qRT-PCR验证。结果显示,qRT-PCR检测结果与转录组数据(FPKM)表达趋势基本一致(图8)。
3 讨论
本研究基于转录组数据,对西伯利亚蝗卵早期发育(DS)、滞育(ES)及滞育后发育阶段(PS)进行比较分析,发现DSvsES和PSvsDS两比较组的DEGs均上调表达居多(图4),说明滞育阶段生理活动和生长速度虽然较慢,但一些基因仍呈上调表达。GO条目富集图分析发现(图5),DSvsES组富集最显著的GO条目被注释为蛋白结合,其次为肌动蛋白结合、细胞骨架蛋白结合等;PSvsDS组中DEGs富集的GO条目主要包含酶活性、细胞骨架构建及蛋白结合等过程,对胚胎发育具有重要作用。
有研究发现Wnt信号通路与生长发育息息相关,在胚胎分化、控制胚胎轴向的正常发育中起关键调控作用(Reya and Clevers, 2005)。Wnt信号失调将导致骨骼肌发育产生缺陷或胚胎夭折(Ridgewayetal., 2000; 韩琳和冯新港, 2008; 张宗明等, 2019)。本研究中,Wnt信号通路显著富集(q<0.05),富集的Wnt基因为WNT6和WNT7,推测WNT6和WNT7可能对西伯利亚蝗卵胚胎组织及神经发育具有重要作用。此外,蝗卵在ES至DS阶段的WNT6和WNT7均呈上调表达,而DS至PS阶段WNT6呈下调表达,WNT7则持续上调(图7: A),说明蝗卵的上皮组织可能在滞育阶段已经发育完善,而神经发育是一个持续过程,在滞育后发育阶段WNT7的上调表达使该发育过程更加活跃。Gros等(2009)证明由WNT11介导的PCP通路能直接影响早期肌细胞定向生长,本研究中WNT11基因在PSvsDS组的调节干细胞多能性的信号通路中显著富集,因此认为WNT11对西伯利亚蝗卵肌细胞的定向生长极为重要,为后续蝗虫发育生物学研究提供了方向。此外,WIF1与SFRP5作为Wnt信号通路的抑制基因,SFRP5表达量逐渐增加,WIF1表达量则在滞育阶段最高,说明这些抑制因子可能协同调控蝗卵发育。
已有研究表明,SAD和SPO属于蜕皮类固醇酶,且均为细胞色素P450酶(Ogiharaetal., 2017);细胞色素P450在合成蜕皮激素过程中起关键作用,而蜕皮激素又在滞育调节中具有重要作用(Gilbert and Warren, 2005; Cuietal., 2019)。甘蓝夜蛾Mamestrabrassicae中SPO的表达高于其他促蜕皮激素基因,且SAD和SPO在滞育期间表达量均降低(Ogiharaetal., 2017),而西伯利亚蝗卵的SAD与SPO在DSvsES组均呈上调表达,且滞育阶段SPO表达量更高,推测蝗卵滞育期SPO在蜕皮激素合成过程中起主要作用。此外,本课题组通过检测西伯利亚蝗卵各阶段蜕皮激素含量,发现滞育阶段的蝗卵蜕皮激素含量高于早期发育阶段(未发表),这与本研究中蝗卵SAD和SPO的表达趋势一致(图7: E),由此推测在蝗卵整个发育过程中SAD与SPO协同调控蝗卵发育,但其具体功能还需进一步验证。
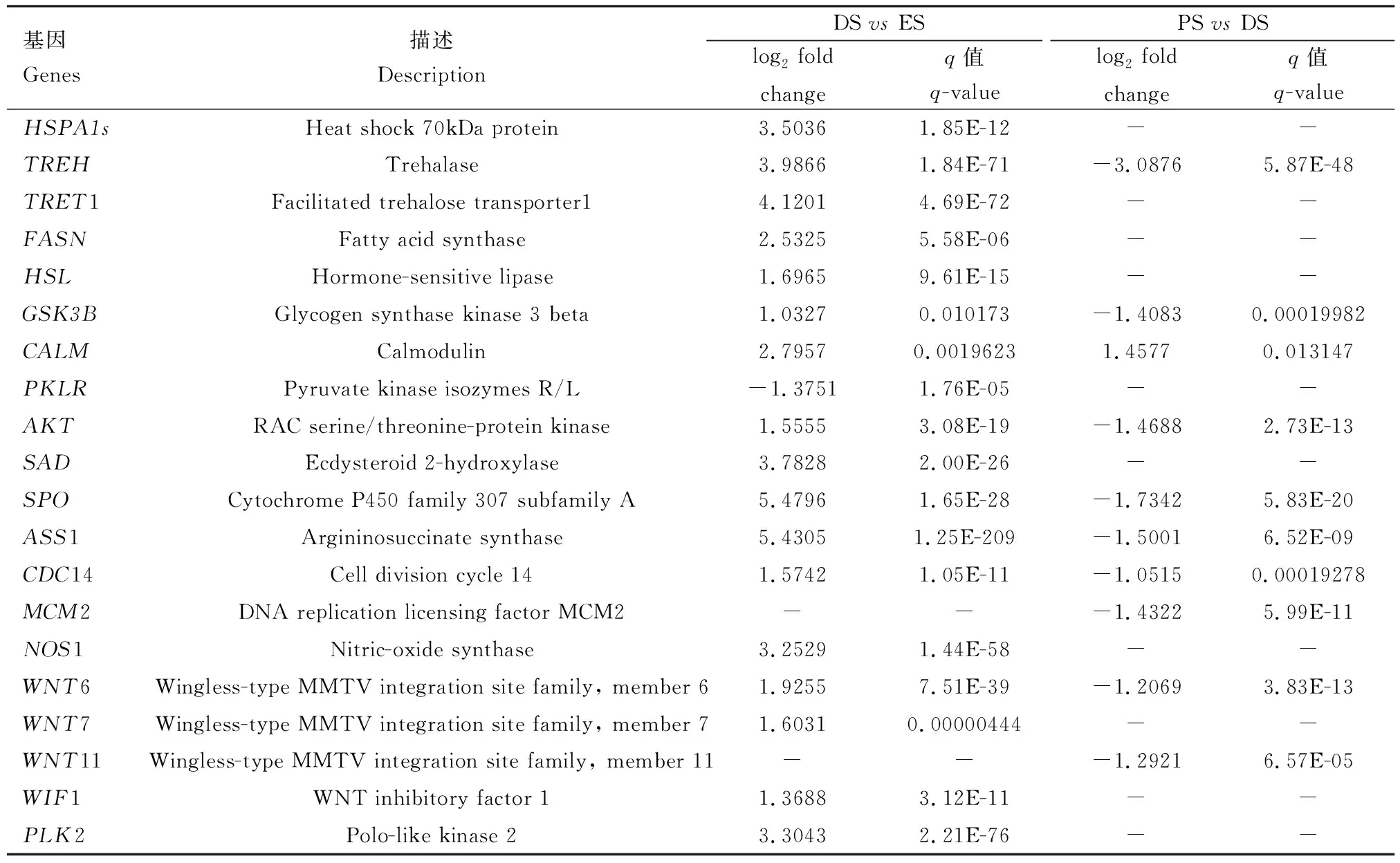
表2 西伯利亚蝗卵潜在的滞育关联基因Table 2 Potential diapause-associated genes of Gomphocerus sibiricus eggs
本研究中,DSvsES组有4个基因(Cluster-37469.111066, Cluster-37469.103701, Cluster-37469.68378和Cluster-37469.32248)共同编码热激蛋白;其中,Cluster-37469.111066被注释为差异基因HSPA1s,且呈上调表达。定量研究发现,HSPA1s在PS阶段仍呈上调表达。蝗卵在滞育期间,为应对外界低温,通过升高HSPA1s的表达从而顺利度过滞育期;随着外界环境温度升高,蝗卵解除滞育,HSPA1s表达量仍然升高,可能是因为刚解除滞育的蝗卵还不能完全适应外界环境变化,仍需合成大量热激蛋白以帮助蝗卵应对外界环境。皱纹盘鲍Haliotisdiscushannai幼鲍在经过热激后其HSPA1s的表达水平显著上调(高婷婷, 2019),这与西伯利亚蝗卵滞育后发育阶段HSPA1s的表达趋势一致;而麦红吸浆虫幼虫破茧恢复发育后HSP则降至最低水平(赵佳佳等, 2018),说明热激蛋白在不同物种体内调控机制不同。HSP90A可与类固醇受体和激酶结合,PSvsDS组中HSP90A(cluster-37469.5850)呈下调表达,说明蜕皮类固醇激素水平相应较低,这与蝗卵中蜕皮激素相关基因SPO表达趋势一致。丝光绿蝇Luciliasericata中HSP90的表达不受滞育影响,但在滞育后发育过程其表达量迅速上调(Rinehartetal., 2007),说明不同热激蛋白在卵滞育及滞育解除过程中可具有不同功能。
昆虫滞育的一个重要标志是细胞周期停滞(Kostáletal., 2009),而胚胎滞育通常发生在细胞周期的S期之前(Pavlidesetal., 2011)。CDC14为细胞分裂所必需,采用RNAi干扰秀丽隐杆线虫CaenorhabditiselegansCDC14的表达后出现胚胎死亡(Grunebergetal., 2002)。我们研究发现,在细胞周期通路中CDC14,MCM2,MPS1以及GSK3B在DS阶段表达量最高(图7: D),说明这4个DEGs在蝗卵滞育阶段发挥重要调节作用。尤其是MPS1和MCM2这2个基因仅在PSvsDS组被注释为DEGs,说明这2个基因可能对蝗卵滞育解除也具调控作用。细胞分裂周期蛋白(CDC14)广泛参与调控细胞周期中各生理过程。如,CDC14在真核生物DNA合成后期阻滞中发挥稳定的保守性作用(苏日古嘎和孟峻, 2018; 刘儒等, 2019)。由此推测DS阶段MCM2与CDC14可协同影响西伯利亚蝗卵滞育期的DNA复制与细胞周期进程。
昆虫胰岛素信号通路能够介导糖原合成酶激酶3基因(GSK3B)调控体内糖原及海藻糖等糖代谢过程,从而控制昆虫的各项生命活动;GSK3B可使糖原合成酶磷酸化,导致糖原合成酶失活,并阻碍胰岛素信号通路的传导,从而抑制糖原合成(丁艳娟等, 2019)。本研究中,DSvsES组胰岛素通路(P<0.05)有13个DEGs,除丙酮酸激酶基因(PKLR)下调表达外,其余基因均上调表达。与早期发育阶段相比PKLR在滞育期下调表达,可有效促进糖酵解的发生,表明西伯利亚蝗卵在早期发育阶段的糖酵解通路较活跃,在滞育阶段其糖代谢活性降低,可为蝗卵的滞育维持以及后期发育提供能量。GSK3B在蝗卵滞育阶段上调表达,导致蝗卵机体内糖原含量减少,滞育后发育阶段则显著下调,促进糖原和海藻糖之间转化,在滞育期通过调控胰岛素信号通路及海藻糖代谢通路等相关基因表达来调控蝗卵的糖原及海藻糖代谢,以维持蝗卵滞育期的正常活性;通过对滞育相关通路进行聚类热图分析发现(图7),GSK3B在分析的各通路中均有表达,这印证了该基因在西伯利亚蝗卵发育过程中具有非常重要的作用。TREH在西伯利亚蝗卵滞育期的上调表达,可导致滞育蝗卵的海藻糖含量降低,这与西伯利亚蝗滞育卵内糖原和海藻糖含量较低的结果(Songetal., 2021)一致。脂质是滞育昆虫的基础能量来源,也是胚胎发育的主要能源(夏丹等, 2016)。脂肪酸合成酶是合成脂肪酸的关键酶,与滞育昆虫的脂质积累密切相关,HSL作为激素敏感脂肪酶基因,可调动储存的脂肪用于氧化供能(闫昭明等, 2021)。本研究中,西伯利亚蝗卵滞育阶段FASN和HSL的上调表达,有利于蝗卵在滞育期积累大量脂肪,且HSL可氧化脂肪,为滞育蝗卵供能,这与美国白蛾Hyphantriacunea的转录组结果(Dengetal., 2018)一致。钙调蛋白(calmodulin, CALM)作为Ca2+信号传导通路中的主要信号转导分子,可调节细胞增殖、激素分泌等重要生理过程(赵岚等, 2007)。本研究中,在PSvsDS组,胰岛素通路中CALM的持续上调表达说明该基因对西伯利亚蝗卵发育具有重要意义。
除上述滞育相关通路中所富集的DEGs外,还有部分基因在DSvsES组和PSvsDS组中差异显著。其中,精氨酸琥珀酸合酶(ASS1)是精氨酸生成的限速酶,精氨酸又可以通过一氧化氮合酶(NOS)生成NO,NO则作为一种重要的信息分子参与调节昆虫的嗅觉、视觉和发育(王晓安和郑哲民, 2003; 黄祝, 2014)。西伯利亚蝗卵在滞育阶段,其ASS1和NOS1都呈上调表达,推测蝗卵可能通过大量表达ASS1和NOS1使得NO含量增加,从而调控蝗卵复眼的发育,滞育蝗卵的复眼已基本发育完成,且有大量色素堆积;PLK2作为polo样激酶在细胞周期中具有重要作用(Maetal., 2003),且PLK2的过表达可促进糖原合成酶激酶GSK3B磷酸化,而PLK2敲除则降低GSK3B磷酸化水平(Fanetal., 2021),结合本研究结果,推测PLK2在蝗卵滞育阶段的上调表达可能参与蝗卵滞育的关键调控。
猜你喜欢
杂志排行

昆虫学报的其它文章
- 草地贪夜蛾烟碱型乙酰胆碱受体α6和α7亚基基因克隆及其对乙基多杀菌素胁迫的响应
- Effects of arena shape and exit location on the escapingbehavior of the Formosan subterranean termite, Coptotermesformosanus (Blattodea: Rhinotermitidae)
- 环丙氨嗪对鸡粪中亮斑扁角水虻生长的影响及添加活性炭对环丙氨嗪的解毒作用
- 多寄主型寄生蜂蝇蛹俑小蜂对新寄主的选择与适应性
- 独栖蜂黄缘蜾蠃的产卵行为及其与寄生胁迫的关系
- Microstructure and defense function of the acoustic organ ofPapilio xuthus (Lepidoptera: Papilionidae) pupae