草地贪夜蛾取食Cry1Ab和Cry1Fa蛋白后中肠转录组及ABC基因表达分析
2022-10-18黄晓丹静大鹏张天涛王振营
黄晓丹, 静大鹏, 张天涛, 王振营,*
(1. 吉林农业大学生物防治研究所, 天敌昆虫应用技术工程研究中心, 长春 130118;2. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193)
苏云金芽孢杆菌Bacillusthuringiensis(Bt)主要以其芽孢形成时产生多种有杀虫作用的晶体蛋白而发挥作用(El-Menofyetal., 2014)。当害虫取食Bt蛋白后,Bt蛋白会在中肠的碱性环境中被溶解活化,从而与中肠刷状缘膜囊泡上的特异性受体结合而发挥作用。中肠上的特异性受体主要有:氨肽酶N(aminopeptidase N, APN)、钙黏蛋白(cadherin, CAD)、碱性磷酸酯酶(alkaline phosphatase, ALP)和腺苷三磷酸结合盒转运蛋白(ATP-binding cassette transporter, ABC)(Jurat-Fuentesetal., 2021)。 其中,ABC转运蛋白是一类广泛存在于原核生物和真核生物体内的跨膜转运蛋白,可利用ATP水解产生的能量对底物进行逆浓度梯度的跨膜运输(Wuetal., 2019),这种运输具有内向转运与外向转运两种构象,转运方向的变化由两种构象对底物的亲和力决定,在真核生物中大多表现为外向转运(刘艳青和赵永芳, 2017),这种特殊的“双向”功能可使得其能够参与生物的细胞内外渗透压平衡、营养摄入、细胞解毒、信号转导等过程。当前,ABC转运蛋白在人体中研究最多的是其外排功能,如魏宁等(2010)研究发现人体肿瘤细胞的多药耐药性与ABC转运蛋白的功能密切相关。 当细胞内P-糖蛋白(ABCB1)、多药耐药相关蛋白(ABCC1)、乳腺癌耐药蛋白(ABCG2)基因过度表达时,会将大量化疗药物排出体外,开启ABC转运蛋白的外排泵功能;而在鳞翅目昆虫中,ABC转运蛋白被证明与昆虫对Bt蛋白抗性产生有关(Heckel, 2012)。例如,在棉铃虫Helicoverpaarmigera、甜菜夜蛾Spodopteraexigua、亚洲玉米螟Ostriniafurnacalis等昆虫体内ABCC2和ABCC3转运蛋白的表达量下调及基因突变提高了这些害虫对Cry1A类蛋白抗性水平(徐丽娜, 2010; Parketal., 2014; Zhouetal., 2016)。因此,明确Bt蛋白对昆虫体内ABC基因表达水平的影响,筛选并发现与抗性相关的新基因并进行功能研究,对防治草地贪夜蛾Spodopterafurgiperda乃至对其他害虫的防控都十分必要。
草地贪夜蛾属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae),源于美洲热带及亚热带地区(Murúaetal., 2009),2019年1月在云南省首次发现其入侵我国。由于其繁殖能力强、迁飞速度快、寄主范围广等特点,快速在我国南方地区完成定殖,并随着季节变暖及玉米的种植逐渐向长江流域、黄淮海夏玉米区和北方春玉米区蔓延为害(Guoetal., 2019)。草地贪夜蛾主要为害玉米、高粱、小麦和水稻等农作物。当前入侵我国的草地贪夜蛾为玉米型,主要取食玉米,对国内玉米生产安全造成了严重威胁(Jingetal., 2020)。在美洲,防控草地贪夜蛾主要以种植转苏云金芽孢杆菌(Bt)抗虫转基因玉米为主(李红梅等, 2019),包括表达Cry1Ab的Bt玉米MON810和表达Cry1Fa的玉米TC1507 (Buntin, 2008)。然而,在美洲,随着Bt作物的连续种植,草地贪夜蛾已经对表达这两种Bt蛋白基因的抗虫玉米产生了不同程度的抗性。为此,研究草地贪夜蛾对Bt蛋白的抗性机制,为我国转基因抗虫玉米产业化种植,延缓抗性产生具有重要意义。本研究利用转录组测序技术研究了草地贪夜蛾幼虫分别饲喂Cry1Ab和Cry1Fa两种Bt蛋白后,中肠上ABC转运蛋白基因表达量变化,为明确草地贪夜蛾ABC转运蛋白在Bt蛋白杀虫机制中的作用,以及合理应用Bt蛋白防治草地贪夜蛾,延缓抗性发展提供了理论依据。
1 材料与方法
1.1 试虫饲养
草地贪夜蛾幼虫于2019年6月1日采自云南省德宏州芒市冬玉米田,后经中国农业科学院植物保护研究所玉米害虫组多代饲养,饲养温度为(26±1)℃,相对湿度为(65±5)%,光周期为16L∶8D,利用草地贪夜蛾人工饲料(王世英等, 2019)饲养至4龄幼虫进行试验。饲养过程中未接触任何Bt制剂和Bt杀虫蛋白。实验所用的Cry1Ab和Cry1Fa活化晶体蛋白购自乐士宁公司,纯度为98%的白色粉末晶体,均用50 mmol/L、pH 10.0的Na2CO3溶解。另外,使用Easy Protein Quantitative Kit试剂盒(北京全式金生物技术有限公司)测定蛋白浓度,保存在-20℃用于后续实验。
1.2 幼虫中肠RNA提取及测序
前期按照一定晶体蛋白浓度梯度进行了草地贪夜蛾初孵幼虫的生物测定,得到初孵幼虫的LC50和LC70。因为草地贪夜蛾4龄幼虫属于高龄幼虫,在进行转录组测序时,为保证4龄幼虫充分取食了两种Bt蛋白进入中肠并确保体内一些基因发生变化,因此选择了LC70作为饲料混合标准。每个处理选取体型相近的8头草地贪夜蛾4龄幼虫为1个生物学重复,饥饿处理4 h,使用含有活化晶体蛋白Cry1Ab(LC70=240.2 μg/g)和Cry1Fa(LC70=270.0 μg/g)的人工饲料利用饲料混合法进行饲喂。具体方法为:将做好的人工饲料冷却到50℃时(未凝固前),将配制好的Cry1Ab和Cry1Fa蛋白倒入人工饲料后立刻搅拌均匀,晾干后分装在24孔板中;以饲料中加入同等体积的水为对照。每个处理组和对照组均设3个生物学重复。预实验中通过测定幼虫取食蛋白后体内谷胱甘肽S-转移酶(glutathione S-transferase)的变化情况而确定取食48 h为体内表达量的高峰期,表明其体内解毒功能在此时间段发挥较好作用(数据未展示),从而确定本试验幼虫取食48 h后采用TRIzol法提取草地贪夜蛾幼虫中肠总RNA。使用NanoDrop 2000超微量分光光度计和Agilent 2100生物分析仪进行RNA质量检测,采用Illumina NovaSeq 6000平台进行转录组测序(北京百迈客生物科技有限公司)。
1.3 mRNA建库、组装与功能注释
采用带有Oligo(dT)的磁珠富集mRNA,随后加入Fragmentation Buffer将mRNA进行随机打断,以mRNA为模板,用六碱基随机引物(random hexamers)合成cDNA第1链,然后加入缓冲液、dNTPs、RNase H和DNA Polymerase Ⅰ合成cDNA第2链,利用AMPure XP Beads纯化cDNA;将纯化的双链cDNA再进行末端修复、加A尾并连接测序接头,然后用AMPure XP Beads进行片段大小选择,最后通过PCR富集得到cDNA文库。文库经过质检后即可上机测序,随后对初步得到的原始数据(raw reads)进行数据评估,过滤除杂和冗余处理,得到高质量的clean reads后通过Hisat 2软件(Kimetal., 2015)与草地贪夜蛾的基因组数据(assembly ZJU_Sfru_1.0)gtf文件进行对比,随后将比对上的reads采用Stringtie(Perteaetal., 2015)进行转录组组装。最后,采用KOG/COG, KEGG, GO数据库对BlastX转录组基因进行注释。
1.4 差异表达基因筛选
比较经Cry1Ab和Cry1Fa分别处理后的幼虫与未添加任何Bt蛋白正常人工饲料饲喂的对照中肠转录组数据,筛选差异表达基因,采用FPKM(fragments per kilobase million fragments)方法进行计数,获得基因相对表达水平,并采用Pearson相关系数(R)作为评价每个处理组3个生物学之间相关性的指标。使用DESeq2软件(Loveetal., 2014),以Cry1Ab和Cry1Fa处理组与对照组分别相比之后,各比较组的整体FDR<0.01和FPKM值的Fold change≥2作为阈值来确定基因表达水平的差异显著性。
1.5 RT-qPCR验证差异表达ABC基因
前期经转录组数据筛选及RT-qPCR验证在Cry1Ab和Cry1Fa处理后,Cry1Ab处理组与对照组相比有4个ABC基因表达量显著变化,Cry1Fa处理组与对照相比有5个ABC基因表达量显著变化。其中,有2个相同基因在两种蛋白处理后均发现有上调趋势,综合后共筛选到6个基因。利用RT-qPCR验证1.4节筛选的差异表达ABC基因(表1)表达量。采用Trizol法提取草地贪夜蛾幼虫中肠的总RNA,加入1 μg RNA模板按照One-step gDNA Removal and cDNA Synthesis Kit(北京全式金生物技术有限公司,北京)说明书进行反转录,合成cDNA第1链。RT-qPCR利用ABI 7500 Fast Real-time PCR System(Applied Biosysterm, 美国)两步法进行,同时以GAPDH作为内参基因(Shuetal., 2021),采用2-ΔΔCt法计算基因的相对表达量(Livaketal., 2014)。RT-qPCR参照PrefectStartTMGreen RT-qPCR SuperMix试剂盒(北京全式金生物技术有限公司,北京)说明书进行。PCR反应体系: PrefectStartTMGreen qPCR SuperMix 10 μL, 上下游引物(10 μmol/L)各0.4 μL, 50×ROX Reference Dye 0.3 μL, cDNA模板1 μL, RNase-free H2O 7.9 μL。扩增程序: 94℃ 30; 94℃ 5 s, 60℃ 30 s, 共40个循环。实验设置3个生物学重复,3个技术重复;所有引物见表1。
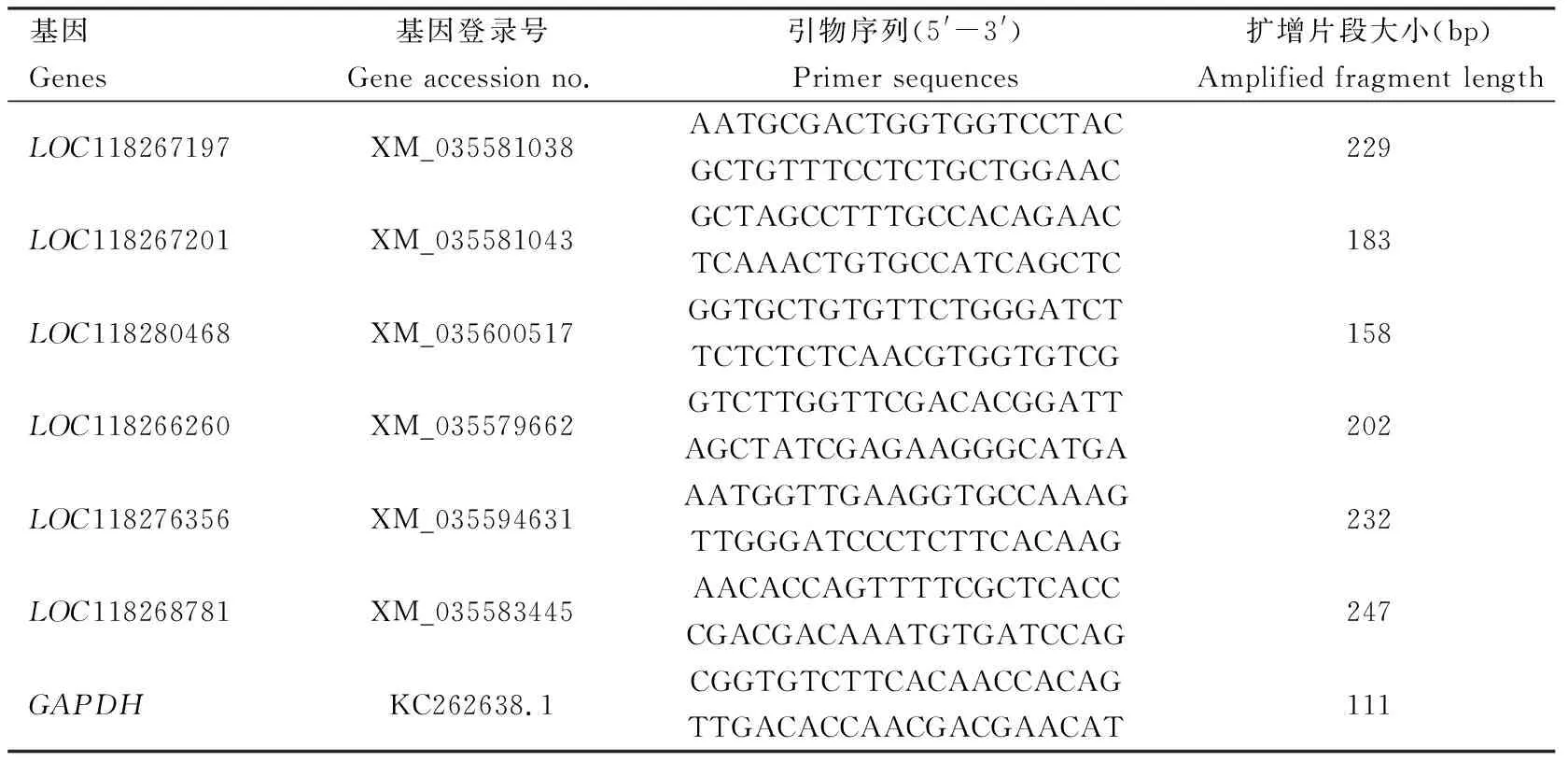
表1 RT-qPCR引物信息Table 1 Primer information for RT-qPCR
1.6 数据分析
应用PoLo-Plus软件计算草地贪夜蛾初孵幼虫生物测定结果LC70值及95%置信区间。RT-qPCR结果利用SPSS 23.0采用单因素方差分析进行差异显著性检验,显著水平设定为P=0.5。
2 结果
2.1 中肠转录组
取食含240.2 μg/g Cry1Ab和270.0 μg/g Cry1Fa人工饲料的处理组(Cry1Ab和Cry1Fa处理组)及取食正常人工饲料的对照组草地贪夜蛾4龄幼虫中肠转录组分别获得了19 427 579, 24 201 561, 20 865 946, 21 655 163, 20 029 632, 27 003 412, 22 705 906, 28 317 771和24 258 102条Reads。对照组、Cry1Ab处理组和Cry1Fa处理组转录组GC含量平均值分别为48.31%, 49.04%和49.45%,均大于45%;Q30平均值分别为95.29%, 94.92%和94.74%,均大于94%,说明测序数据质量良好,获得了高质量reads。此外,将所获得的reads与草地贪夜蛾参考基因组进行比对匹配基因组的reads达到了80%以上,多个比对位置也达到10%以上,说明总read和参考基因组匹配程度较高,可以做进一步的分析。
2.2 差异表达基因鉴定及GO分类
Cry1Ab处理后草地贪夜蛾4龄幼虫中肠转录组与对照组相比检测到1 305个差异表达基因,700个上调,605个下调;Cry1Fa处理后的草地贪夜蛾4龄幼虫中肠转录组与对照组相比检测到1 202个差异表达基因,663个上调,539个下调。基因GO分类功能注释结果表明(图1),Cry1Ab和Cry1Fa处理组分别有994和912个差异表达基因被注释到生物学过程(biological process)、分子功能(molecular function)和细胞组分(cellular component)三大类。
2.3 差异表达ABC基因RT-qPCR验证
与对照组相比,饲喂Cry1Ab和Cry1Fa后草地贪夜蛾4龄幼虫中肠转录组中有9个ABC基因显著差异表达。其中,Cry1Ab处理组中有4个ABC基因(LOC118267197,LOC118267201,LOC118267200,LOC118280468)差异表达,3个上调,1个下调;Cry1Fa处理组与对照组比较有5个ABC基因(LOC118266260,LOC118276356,LOC118268781,LOC118267201,LOC118267200)差异表达,2个上调, 3个下调;Cry1Ab和Cry1Fa处理组共有的739个差异表达ABC基因中有2个基因(LOC118267201和LOC118267200)在Cry1Ab和Cry1Fa处理组中同时显著上调,没有同时下调的基因。
RT-qPCR结果显示(图2),Cry1Ab处理组与对照组中肠转录组比较有4个ABC基因的表达量显著上调,且有3个(LOC118267197,LOC118266260和LOC118276356)表现为极显著上调(P<0.001),另外2个ABC基因(LOC118268781和LOC118280468)表达量分别显著降低(P<0.05)和极显著降低(P<0.001);Cry1Fa处理组与对照组中肠转录组比较有5个ABC基因(LOC118267197,LOC118266260,LOC118276356,LOC118280468和LOC118267201)表达量上调,其中LOC118267197,LOC118266260和LOC118267201基因为极显著差异(P<0.001),另外1个基因(LOC118268781)下调,但与对照组之间无显著差异 (P>0.05)。将转录组数据中FPKM值与RT-qPCR结果关联分析后,由以上结果可看出,在Cry1Ab和Cry1Fa两种蛋白分别处理后,除个别基因外,有多个ABC相关基因表达量与转录组中的基因表达趋势发生一致变化(图3)。 经比对后,LOC118266260为多药耐药蛋白同系物ABCC家族基因,LOC118280468为ABCG8基因。值得注意的是,经筛选比对发现,LOC118280468(ABCG8)在Cry1Ab处理后表达量极显著下调(P<0.001)(图2: E),在Cry1Fa处理后表达上调但与对照组之间无显著差异(P>0.05)。其他未命名基因需进一步筛选鉴定。
3 讨论
本研究通过转录组测序技术筛选草地贪夜蛾幼虫分别取食Cry1Ab和Cry1Fa蛋白后体内ABC转运蛋白在中肠的表达变化情况。结果表明,取食Cry1Ab后,共有4个ABC转运蛋白编码的基因表达量发生变化,取食Cry1Fa后,共有5个ABC基因表达量发生变化(图2)。其中,有两个相同基因在取食含有Cry1Ab和Cry1Fa的人工饲料后表达量都发生显著变化。Cheng等(2017)在比较基因组分析中发现多食性害虫斜纹夜蛾Spodopteralitura的解毒相关基因家族,如P450, GST, COE, APN和ABC等基因的拷贝数明显上调。已有研究表明,基因的复制或扩增使基因过表达是节肢动物适应性进化中的常见机制,一些代谢酶就是复制“热点”。因此,害虫可能通过扩增过表达这些解毒基因从而降解进入体内的化学农药或Bt蛋白,为多食性害虫进化成一种全球性重大害虫提供了依据(Bass and Field, 2011)。ABCC家族作为目前最多被研究的与Bt蛋白产生抗性相关联的家族,其被报道介导了昆虫对多种Bt蛋白的抗性。其中,使用Cry1Ab和Cry1Ca两种Bt蛋白处理甜菜夜蛾,当ABCC2与ABCC3表达量降低时,甜菜夜蛾死亡率显著降低。同时,ABCC2与ABCC3两种转运蛋白序列同源性高于70%,很可能这两个蛋白都可以作为Cry1A毒素的受体(Parketal., 2014)。本研究通过转录组测序和RT-qPCR技术验证了当草地贪夜蛾幼虫经两种Bt蛋白分别处理后,ABCC家族基因(LOC118266260)在Cry1Ab和Cry1Fa处理后表达量均上调而ABCG8基因在Cry1Ab处理后表达量下调,Cry1Fa处理后表达量上调(图2)。表明这两个ABC基因参与了Bt蛋白作用于草地贪夜蛾幼虫中肠的过程。研究发现,ABCG家族在植物中存在内向转运的机制,但在真核生物中还尚未发现(Gräfe and Schmitt, 2020)。值得注意的是,ABCG家族在人类疾病的研究中,特别是ABCG2即乳腺癌耐药蛋白,可形成二聚体结构的活性转运复合物,以“药泵”形式减少胞内ATP依赖性药物蓄积而产生耐药被人们所熟知(Coley, 2008)。本研究初步筛选出ABCC家族与ABCG家族的ABCG8基因,在经过Bt蛋白处理后表达量均发生显著变化,表明ABCC家族与ABCG家族基因表达量的变化可能会影响草地贪夜蛾对Bt蛋白的敏感性,可能导致草地贪夜蛾抗药性的产生,具体的功能还需要深入验证。此外,在烟粉虱Bemisiatabaci和小菜蛾Plutellaxylostella等研究中提出ABC基因表达量下调介导昆虫对噻虫嗪抗性以及Bt毒素Cry1Ac抗性的产生(Yangetal., 2013; Zhouetal., 2020),类似的功能基因还有ABCB1(P-糖蛋白)和ABCC亚家族(通常称MRP亚族)等(魏宁等, 2010)。目前尚未见ABCG1与ABCG8在昆虫中的研究报道,但通过对7种节肢动物ABC转运蛋白亚家族和人类ABC转运蛋白亚家族进行系统全面分析后发现,推断其可能具有保守功能的同源关系(Dermauw and Leeuwen, 2014)。因此,还需要更加深入的研究。
基因上调不仅可以达到解毒的目的,许多ABC基因下调或者基因序列缺失也可影响其抗药性。有研究表明敲除小菜蛾的ABCC2基因后,其对Cry1Ac的抗性升高了3.8倍,而同时敲除ABCC2和ABCC3基因后其对Cry1Ac的抗性倍数显著升高了1 000倍以上(Wangetal., 2020)。当敲除草地贪夜蛾ABCC2基因时,与野生型相比,对Cry1Fa的抗性倍数提高了118倍,但对营养期杀虫蛋白Vip3A类蛋白没有抗性,则证明ABCC2是Cry1Fa的功能性受体(Jinetal., 2021)。因此,ABC基因的突变或者表达量下调也可能会影响昆虫利用这些转运体来清除自身毒素的能力。而在本研究中也发现了显著下调的ABC家族相关基因(LOC118268781),可能与昆虫解毒或者抗性密切相关,这为后续的研究奠定了基础。
本研究通过测定Cry1Ab和Cry1Fa两种Bt蛋白处理后草地贪夜蛾幼虫中肠的基因变化,除了与抗性相关的ABC转运蛋白的挖掘,还测定到其他抗性相关的免疫蛋白,如农药抗药性相关基因,在后续研究昆虫对Bt蛋白以及农药产生抗性的作用靶标突变及各类抗药性基因的相互作用关系奠定了基础,为寻找解决昆虫抗药性提供了方向。
