阳春砂与海南砂BPPS启动子比较及GCN4-motif正调控作用的鉴定
2022-10-14林晓静梁慧琳赵海莹吴清文黄琳璇伍思容杨锦芬
林晓静,梁慧琳,赵海莹,吴清文,黄琳璇,伍思容,杨锦芬
• 药材与资源 •
阳春砂与海南砂启动子比较及GCN4-motif正调控作用的鉴定
林晓静,梁慧琳,赵海莹,吴清文,黄琳璇,伍思容,杨锦芬*
广州中医药大学中药学院 中药资源科学与工程研究中心,岭南中药资源教育部重点实验室(广州中医药大学),国家中成药工程技术研究中心南药研发实验室,广东 广州 510006
龙脑基二磷酸合酶(bornyl diphosphate synthase,BPPS)是砂仁药效萜类合成途径中的关键酶,获得海南砂的启动子并与阳春砂启动子进行调控元件和瞬时表达的比较,以期解析在种子中高表达的分子机制。通过FPNI-PCR的方法从海南砂gDNA中克隆的启动子,并与阳春砂启动子进行序列比较,对启动子相似区域进行截短,获得其相应的截短片段;对启动子截短片段中的GCN4-motif进行突变;构建由上述启动子驱动β-葡萄糖苷酸酶基因(β-glucuro-nidase gene,)的重组表达载体,利用农杆菌介导法注射侵染本氏烟草叶片进行瞬时表达,验证启动子的活性和GCN4-motif的功能。克隆获得365 bp的启动子序列,3’端约300 bp的序列与启动子的相应序列相似度高达93%,但启动子不含有启动子的GCN4-motif。构建成功与GUS报告基因融合的系列启动子重组载体——(启动子全长)、(启动子截短至320 bp)、(截短启动子且突变GCN4-motif)和(启动子全长)、(启动子截短至305 bp)。通过GUS染色发现虽然上述启动子均具有驱动基因转录的活性,然而,含有GCN4-motif的启动子,包括全长或截短的启动子(和)的活性均高于GCN4-motif突变()或无GCN4-motif的启动子(和)。GCN4-motif对基因转录具有正调控作用,为进一步探究启动子的调控机制奠定了基础。
启动子;龙脑基二磷酸合酶;阳春砂;海南砂;GCN4-motif;瞬时表达
砂仁是我国“四大南药”之一,具有化湿开胃、温脾止泻等功效,可用于湿浊中阻、脾胃虚寒等症,《中国药典》2020年版收录砂仁来源于姜科豆蔻属植物阳春砂Lour.、海南砂T. L. Wu或绿壳砂Lour. var.T. L. Wu et Senjen的干燥成熟果实,并规定挥发油和乙酸龙脑酯为其主要药效成分[1]。阳春砂因高含量挥发油和乙酸龙脑酯,其品质通常优于海南砂和绿壳砂,国内市场对阳春砂的需求远大于后两者,使其长期处于供需紧张的状况[2-4]。因此,分析阳春砂关键酶的分子调控机制,从分子角度解析阳春砂品质优良的原因,为优化海南砂和绿壳砂品质奠定基础,进而缓解品质优良砂仁的供需缺口。
龙脑基二磷酸合酶(bornyl diphosphate synthase,BPPS)是砂仁药效萜类合成途径的关键酶,催化-半乳糖-1-磷酸酯(geranyl diphosphate,GPP)生成龙脑基二磷酸合酶(bornyl diphosphate,BPP),而BPP则为砂仁药效物质龙脑的直接前体和乙酸龙脑酯的间接前体[5]。Wang等[6]、李萌[7]分别克隆并鉴定阳春砂和海南砂的和,除了自身基因功能差异,同时期种子团中表达量远高于[6-7]。启动子在基因的表达调控中起着重要的作用,推测和的启动子所调控的转录差异是导致海南砂和阳春砂品质差异较大的重要原因之一。因此,将比较分析海南砂和阳春砂BPPS启动子序列,以期阐明不同来源BPPS转录差异的原因。
课题组前期已克隆获得470 bp的启动子序列,发现启动子序列中具有参与胚乳特异表达的GCN4-motif[TGA(G/C)TCA][8]。GCN4-motif是一种在谷作物种子贮藏蛋白启动子中高度保守的顺式元件,对控制醇溶蛋白和谷蛋白的胚乳特异性表达起着核心作用[9-11]。然而GCN4-motif在非贮藏蛋白种子特异性启动子的研究较少,作为非贮藏蛋白,表现出在种子中高表达特点和其产物表现出种子富集的特性[6,12],初步推断单个GCN4-motif在非贮藏蛋白种子特异性启动子中可以介导基因在种子中特异性表达且具有正调控的作用。因此,本研究重点关注启动子中的GCN4-motif,鉴定GCN4-motif对非贮藏蛋白AvBPPS的正调控作用,为提高关键基因的表达提供分子调控元件,为砂仁品质优化及药效成分的代谢工程奠定基础。
1 材料与试剂
1.1 材料
海南砂种植于广州中医药大学大学城校区,经广州中医药大学何国振教授鉴定为海南砂T. L. Wu,本研究取其嫩叶并保存至−80 ℃超低温冰箱备用;野生型本氏烟草L.、pLB-p载体质粒和pCAMBIA1301植物表达载体质粒均为本实验保存。
1.2 试剂
快速定点突变试剂盒、植物基因组DNA提取试剂盒(离心柱型)、pLB零背景快速克隆试剂盒和DNA切胶回收试剂盒购于天根生化科技(北京)有限公司;根瘤农杆菌EHA105、DH5α感受态细胞购于上海唯地生物技术有限公司;Infusion连接酶、限制性内切酶、PCR酶PrimerSTAR、LA Taq和DL2000 Maker购于Takara公司;内切酶I和I均购于New England Biolabs(NEB)公司;GUS染色试剂盒购于索莱宝生物科技有限公司;引物合成和基因测序由北京擎科生物科技有限公司完成。
2 方法
2.1 海南砂基因组DNA的提取
参照植物基因组DNA提取试剂盒的操作说明,提取海南砂叶片的基因组DNA(gDNA),用1%琼脂糖凝胶电泳和微量紫外分光光度计检测DNA完整性及浓度。
2.2 AlBPPS启动子序列的克隆
根据FPNI-PCR引物设计原则以及参考和gDNA序列[8],在5’端参考使用3条方向一致的特异性引物(gene specific primers,GSP),即GSP TPS3-1、GSP TPS3-2和GSP TPS3-3,见表1。
FPNI-PCR分为3轮扩增,第1轮PCR取适量海南砂gDNA,用9条通用简并引物FP1~9分别与特异引物GSP TPS3-1进行热不对称PCR;第2轮PCR取第1轮PCR产物为模板,巢式特异引物FSP1分别与3个基因的特异引物GSP TPS3-2进行普通PCR第3轮方法与第2轮类似,以逐步分离目的DNA片段。将3轮PCR产物进行琼脂糖凝胶电泳检测,用第3轮PCR产物直接测序或连入pLB载体测序。利用Snapgene软件对测序结果与已获得的目的基因gDNA序列分别比对,确认3’端与其gDNA序列的5’端部分序列是否重叠。
2.3 AlBPPS启动子序列分析、比较
利用Neural Network Promoter Prediction(http://www.fruitfly.org/seq_tools/promoter.html)预测转录起始位点,利用PlantCARE(http:// bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测启动子潜在的顺式元件。利用Jalview软件将和启动子序列进行比对,并使用TBtool软件对该启动子的顺式元件位置分布进一步可视化。
2.4 AlBPPS和AvBPPS启动子截短、AvBPPS启动子GCM4-motif突变及其表达载体的构建
通过对比海南砂和阳春砂BPPS启动子的序列,其中高度相似区域包含转录起始位点至起始密码子之前约50 bp,利用In-fusion原理设计同源臂引物并扩增启动子全长和相似区域序列。PCR扩增条件:98 ℃,预变性5 min;98 ℃,变性10 s,58 ℃,退火30 s,72 ℃,延伸2.5 min,30个循环;最后72 ℃延伸5 min。分别回收PCR产物,参照快速定点突变试剂盒的说明,设计引物并进行突变,将启动子截短片段中的TGAGTCA突变为TGTCA,使用VBTp-F和Bp-R的引物扩增并回收该PCR产物(引物见表1)。
将获得的目的片段,即和全长启动子、各自截短至保守区域的启动子,以及启动子截短加GCN4-motif突变的启动子,通过In-fusion酶连接上已经过内切酶I和I酶切的pCAMBIA1301线性化载体,构建与基因融合的植物表达载体,将连接产物转化至DH5α 中,利用菌液PCR和测序验证含有目的片段的阳性重组密码子,并将重组质粒分别命名为和。
2.5 农杆菌介导法转化烟草验证启动子活性
将“2.4”项的重组质粒用液氮速冻法转化农杆菌EHA105菌株,选取阳性的单菌落在25 μg/mL利福平(Rif)和50 μg/mL 卡那霉素(Kan)的LB液体培养基中28 ℃培养36 h,按照1∶50的比例将菌液转接到含有25 μg/mL Rif和50 μg/mL Kan的100 mL液体LB培养基中,200 r/min,28 ℃培养至600值为0.6,4 000×g、28 ℃离心20 min弃上清收集菌体,用含有10 mmol/L MES、10 mmol/L MgCl2和100 μmol/L AS的MS液体培养基重悬菌体,振荡至600为0.8~1.0,在室温静置3 h。最后用针头在烟草叶子背面轻划破小口子,将重悬液用1 mL注射器(去掉针头),从烟草叶片背部缓慢注入,做好标记,保湿培养2 d,用GUS组织化学染色法检测GUS活性,GUS染色参照GUS 染色试剂盒说明书进行。
表1 引物序列
Table 1 Primers sequences
引物用途引物名称引物序列 (5’→3’) 启动子序列扩增GSP TPS1-1GAAGCCCTCTGATCTGGTCCTCACA GSP TPS1-2CAGCTGCGAGCTCACGCCGTCTAAA GSP TPS1-3CGACGACGACCACTACACTCGATCG GCN4-motif突变GM-FGGAAAAAAAAAATGATTGTCAAATTTTCTAGGGAAGG GM-RCCTTCCCTAGAAAATTTGACAATCATTTTTTTTTTCC pCAMBIA1301-BPPS启动子表达载体构建VBp-FGCAGGCATGCAAGCTTTGTGTAGTACCAAACATATTTTCTATTTTCTAAAAATATATTAGA L/VBPTp-F*GTACCCGGGGATCCGGTTTGATTAAAATATACTGCCCTATAATTATTATTAAATTTGTAAATAG LBp-FGCAGGCATGCAAGCTTAGACGACTGAAAAGAAGCGCTG Bp-R*CAGATCTACCATGGGTTTGCAATCAAAAAATCGTAAAATATATCTCAACTCCA
*VBPT和LBPT的5’端和3’端一致,使用同一对引物
The 5’end and 3’end of VBPT and LBPT are the same, and the same primers are used
3 结果与分析
3.1 AlBPPS启动子序列的获得
利用gDNA核心序列设计特异性引物,以海南砂叶片gDNA为模板,进行FPNIPCR,将反应产物进行凝胶电泳检测。结果表明在第3轮PCR扩增中FP3和FP7获得了符合长度的条带,见图1。以PCR产物测序,只有FP7的3’端有262 bp与gDNA序列的5’端重合,初步认定为的5’端上游序列,截去转录起始位点3’端序列之后,获得含有启动子区域的片段365 bp,其AT含量为70%,为植物启动子主要特征之一,见图2。

FP1~FP9-第1轮PCR为9条不同的简并引物的PCR产物 1、2、3分别代表第1、2、3轮PCR产物 M-Marker 橙框为启动子
FP1~FP9-PCR products of first PCR amplified by nine different degenerate primers 1, 2, and 3-PCR products of first, second, and third round of PCR M-Marker The orange box is thepromoter
图1启动子序列FPNI-PCR扩增产物
Fig. 1 Amplification ofpromoter using FPNI-PCR

图2 AlBPPS启动子序列及其顺式作用调控元件
3.2 AlBPPS启动子序列的生物信息学分析
将克隆获得的365 bp序列进行生物信息学分析。利用Neural Network Promoter Prediction进行转录起始位点预测,以预测值0.9为cut-off值,确定起始转录位点位于起始密码子ATG上游62 bp处。并利用PlantCARE对启动子中可能存在的顺式作用元件进行预测,分析启动子元件的类型、数量及位置(图2和表2)。其中“+”“−”区分碱基的位置和正负链,“366”为转录起始位点的位置。依此类推。启动子包含激活蛋白的核心元件TATAbox和提高转录活性的增强元件CAATbox等多个真核生物启动子保守元件。此外,该启动子还有光响应元件Box 4、G-box,涉及干旱诱导的结合位点MBS,涉及厌氧诱导顺式作用的ARE元件,涉及低温诱导顺式作用的LTR元件,涉及脱落酸反应的ABRE,以及参与茉莉酸甲酯响应的TGACG-motif和CGTCA-motif的元件。
表2 AlBPPS启动子预测的顺式作用调控元件
Table 2 Predicted cis-elements in AlBPPS promoter
元件序列功能位置 TATA-boxTATATA核心启动子元件280− TATA-boxTATAAATA 334− TATA-boxTATAAAT 89− 335− TATA-boxTATAAA 90− 336− TATA-boxTATAA 91− 337− 92+ TATA-boxTATA 125+ 135+ 176+ 282− 338− TATA-boxATATAT 279− 281− CAAT-boxCAAAT启动子和增强子区域常见的顺式作元件39− 150− 163− 213+ CAAT-boxCAAT 37+ 58− 240− 323− CAAT-boxCCAAT 109− AT~TATA-boxTATATA 280− AREAAACCA参与厌氧诱导的顺式作用调控元件111− MBSCAACTG参与干旱诱导的 MYB 结合位点45+ G-BoxCACGTT参与光响应的顺式作用调控元件310− Box 4ATTAAT参与光响应的顺式作用调控元件236− LTRCCGAAA参与低温响应的顺式作用调控元件53+ ABREACGTG涉及脱落酸反应的顺式作用调控元件311+ TGACG-motifTGACG涉及茉莉酸甲酯反应的顺式作用调控元件210− CGTCA-motifCGTCA涉及茉莉酸甲酯反应的顺式作用调控元件210+
“+”“−”表示正反链,转录起始位点的位置为366
“+” and “−” indicate the positive and negative strands, and the position of the transcription start site is 366
3.3 AlBPPS与AvBPPS启动子序列的比较与分析
将获得的启动子与启动子进行序列对比,发现有一段相似度高达93%、长度约300 bp的区域(包含转录起始位点至起始密码子之前50 bp)(图3),推测该相似序列为砂仁启动子的保守区域。在这段保守区域中,启动子含有胚乳特异性元件GCN4-motif,而启动子在对应的位置则为茉莉酸甲酯响应元件CGTCA-motif(图3、图4)。鉴于和所表现的转录差异和GCN4-motif的功能报道,初步推测GCN4-motif在的表达中具有胚乳特异性表达以及正调控的作用。
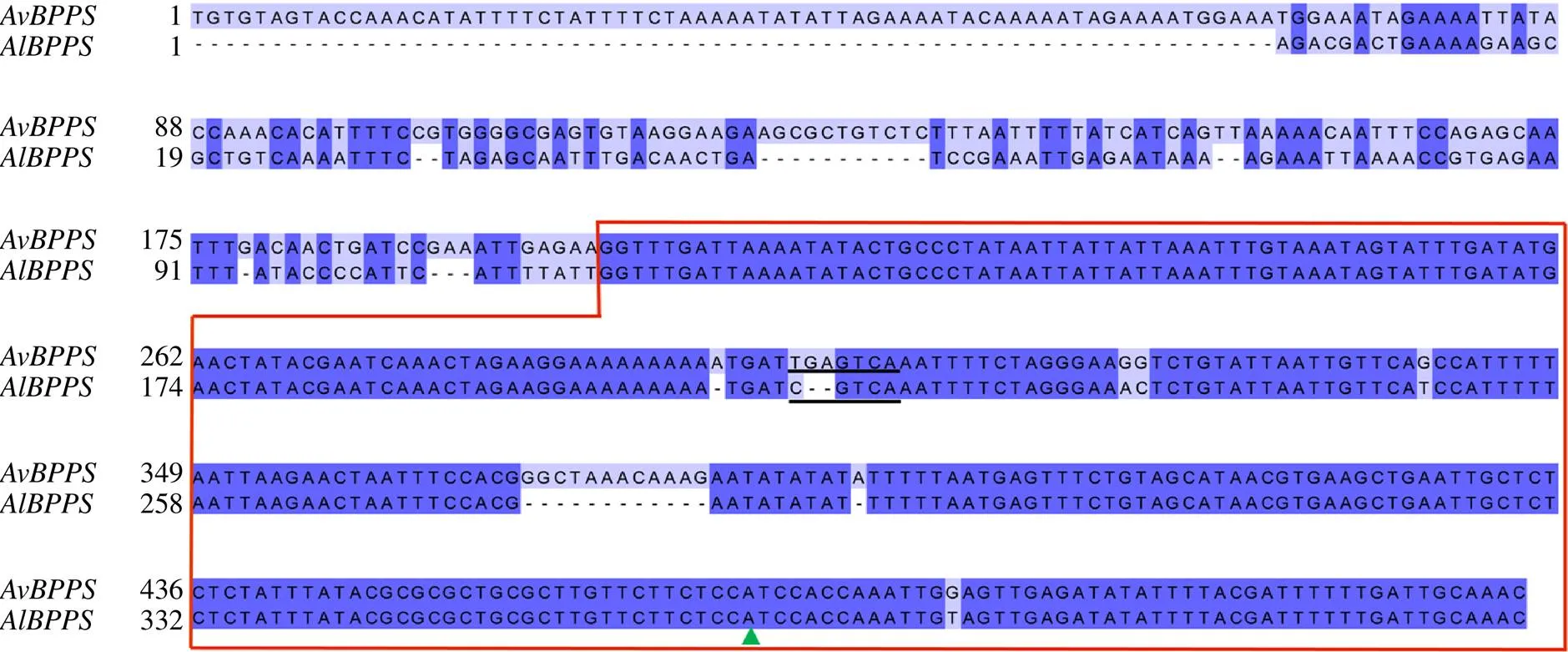
绿色三角形代表转录起始位点,转录起始位点右边为非编码区,红框为高度相似区域,黑色下划线分别代表GCN4-motif和CGTCA-motif
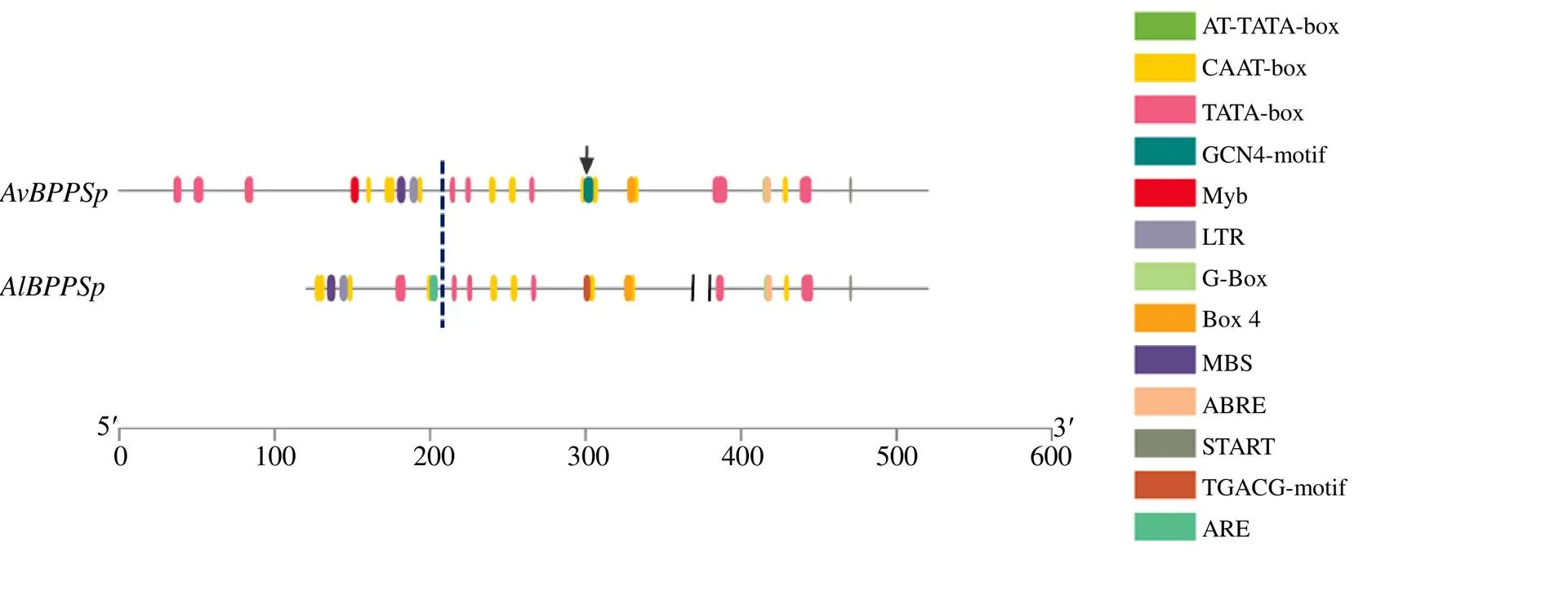
蓝色虚线右边为相似区域,黑色箭头表示GCN4-motif
3.4 BPPS启动子驱动GUS表达载体的构建
构建了和表达载体(图5),对重组质粒进行PCR鉴定,PCR扩增启动子片段与目标片段大小一致(图6),获得阳性重组子,测序结果验证了载体构建成功;其中的GCN4-motif—— TGAGTCA突变成了TGTCA。

TB(R)、TB(L)是T-DNA的左右边界HygR潮霉素抗性基因 GUS-β-葡糖苷酸酶

1~5-VBP∷GUS、VBPT∷GUS、VBPT-GM∷GUS、LBP∷GUS和LBPT∷GUS PCR检测产物 M-Marker
3.5 启动子活性比较分析
分别将重组质粒和阳性对照pCAMBIA1301利用农杆菌介导侵染本氏烟草叶片,保湿培养2 d,同时以未侵染的叶片作为阴性对照,进行GUS染色。结果如图7所示,阴性对照(野生型)不能染成蓝色,阳性对照叶片呈大面积较深蓝色,和均能染上蓝色,说明上述启动子均能驱动基因的表达。然而,启动子全长或截短的和均呈现较大面积且较深的颜色,而GCN4-motif突变的启动子与其野生型对照相比,颜色明显较浅;不含GCN4-motif的启动子,包括全长和截短的,与其相应的对照启动子和相比,颜色也明显较浅,仅表现出微弱的活性。结果显示含有GCN4-motif的启动子比GCN4-motif突变或缺失的启动子显示出更强的活性,证明GCN4-motif对启动子活性起正调控作用。

图7 BPPS启动子瞬时表达的GUS染色
4 讨论
阳春砂萜类物质主要富集于种子团中,而BPPS作为砂仁药效萜类合成途径的关键酶,也相应地表现出种子特异性表达[6,12]。另外,在阳春砂种子团的表达量不仅显著高于其他的萜类合酶,例如芳樟醇合酶(linalool synthase,AvLIS)和葎草烯合酶(humulene synthase,AvHUS)等萜类合酶,并且在开花后45 d种子团中的表达量约为的200倍[7-8]。基因表达量的差异受上游启动子的影响,因此,比较、和启动子之间的序列和顺式元件差异,有助于解析在种子团中高表达的原因。赵海莹等[8]克隆了、和的启动子,发现启动子具有和启动子不具备的GCN4-motif。本研究在此基础上克隆了海南砂启动子,该启动子具有多个顺式作用元件,包括参与光、低温、干旱、茉莉酸甲酯等逆境响应的顺式作用调控元件;启动子与启动子3’端序列高度相似,然而,启动子却不具有GCN4-motif,相应位置的碱基序列变成了CGTCA-motif—一个参与茉莉酸甲酯(methyl jasmonate,MeJA)响应的调控元件。通过进一步的瞬时表达实验进行验证,发现截短至相似区域与各自相应全长的启动子相比,其活性表现较为一致,说明这段截短至相似区域为砂仁BPPS启动子序列中的关键保守区域。此外,在这段核心保守区域中,包含GCN4-motif的阳春砂启动子活性明显高于不具有GCN4-motif的海南砂启动子。因此,推测GCN4-motif是调控在种子团高表达的重要因素之一。
据文献报道,GCN4-motif通常以组合的模式在贮藏蛋白种子特异性启动子进行调控。例如,在醇溶蛋白启动子中,GCN4-motif与prolamin-box-motif构成双因子胚乳盒,又或者与bZIP类转录因子BLZ2结合进而调节醇溶蛋白的表达[11,13-14];而GCN4-motif在谷蛋白启动子中主要发挥着胚乳特异性表达的作用,通常与ACGT-motif和AACA-motif组成最小元件组合或者与bZIP类转录因子结合调控谷蛋白在胚乳中特异性表达,并起正调控作用。而根据海南砂和阳春砂转录模式和启动子序列,推测单个GCN4-motif对非贮藏蛋白起着胚乳特异性作用以及正调控作用[9-10,15]。本研究对启动子中的GCN4-motif进行突变,瞬时表达的结果显示,突变体与野生型VBPT比较,活性明显降低(图7)。本研究验证了GCN4-motif是非贮藏蛋白启动子的正调控元件,对的表达起促进作用。
本研究首次克隆了海南砂启动子,并将它与阳春砂启动子进行比较,发现砂仁启动子的关键保守序列并鉴定了启动子的关键顺式元件GCN4-motif的正调控作用。下一步将对启动子缺失体和突变体构建稳定遗传的转基因烟草,分析GCN4-motif与种子特异性表达的关系,加深认识GCN4-motif在非贮藏蛋白启动子中的调控机制,为进一步阐明阳春砂和海南砂中的BPPS转录差异模式奠定理论基础。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典 [S]. 一部. 2020: 264.
[2] Ao H, Wang J, Chen L,. Comparison of volatile oil between the fruits ofLour. andLour. var.T. L. wu et Senjen based on GC-MS and chemometric techniques [J]., 2019, 24(9): 1663.
[3] 覃权, 蒋春林, 蒋孟良, 等. 不同产地砂仁中乙酸龙脑酯与总挥发油含量比较研究 [J]. 中医药导报, 2017, 23(14): 70-72.
[4] 黄绿. 阳春砂仁资源综合开发利用研究 [D]. 昆明: 云南中医药大学, 2020.
[5] Despinasse Y, Fiorucci S, Antonczak S,. Bornyl-diphosphate synthase from: A major monoterpene synthase involved in essential oil quality [J]., 2017, 137: 24-33.
[6] Wang H, Ma D M, Yang J F,. An integrative volatile terpenoid profiling and transcriptomics analysis for gene mining and functional characterization of AvBPPS and AvPS involved in the monoterpenoid biosynthesis in[J]., 2018, 9: 846.
[7] 李萌. 阳春砂单萜和倍半萜合酶的功能鉴定及BPPS的功能比较 [D]. 广州: 广州中医药大学, 2019.
[8] 赵海莹, 李萌, 林晓静, 等. 阳春砂3个萜类合酶基因启动子的克隆及其参与萜类调控的分析 [J]. 中草药, 2021, 52(4): 1117-1127.
[9] Wu C, Washida H, Onodera Y,. Quantitative nature of the Prolamin-box, ACGT and AACA motifs in a rice glutelin gene promoter: Minimal-element requirements for endosperm-specific gene expression [J]., 2000, 23(3): 415-421.
[10] Yoshihara T, Washida H, Takaiwa F. A 45-bp proximal region containing AACA and GCN4motif is sufficient to confer endosperm-specific expression of the rice storage protein glutelin gene, GluA-3 [J]., 1996, 383(3): 213-218.
[11] Marzábal P, Busk P K, Ludevid M D,. The bifactorial endosperm box of gamma-zein gene: Characterisation and function of the Pb3 and GZM-acting elements [J]., 1998, 16(1): 41-52.
[12] 赵海莹. 阳春砂萜类合酶及其启动子参与挥发性萜类合成的研究 [D]. 广州: 广州中医药大学, 2020.
[13] Oñate L, Vicente-Carbajosa J, Lara P,. Barley BLZ2, a seed-specific bZIP protein that interacts with BLZ1and activates transcription from the GCN4-like motif of B-hordein promoters in barley endosperm [J]., 1999, 274(14): 9175-9182.
[14] Müller M, Knudsen S. The nitrogen response of a barley-hordein promoter is controlled by positive and negative regulation of the GCN4and endosperm box [J]., 1993, 4(2): 343-355.
[15] Onodera Y, Suzuki A, Wu C Y,. A rice functional transcriptional activator, RISBZ1, responsible for endosperm-specific expression of storage protein genes through GCN4 motif [J]., 2001, 276(17): 14139-14152.
Comparison ofpromoters betweenandand identification of GCN4 motif positive regulation
LIN Xiao-jing, LIANG Hui-ling, ZHAO Hai-ying, WU Qing-wen, HUANG Lin-xuan, WU Si-rong, YANG Jin-fen
Research Center of Chinese Herbal Resource Science and Engineering, Key Laboratory of Chinese Medicinal Resource from Lingnan (Guangzhou University of Chinese Medicine), Ministry of Education Joint Laboratory of National Engineering Research Center for Pharmaceutics of Traditional Chinese Medicines, College of Traditional Chinese Medicine, Guangzhou University of Chinese Medicine, Guangzhou 510006, China
Bornyl diphosphate synthase (BPPS) is an important enzyme in the synthetic pathway ofmedicinal terpenoids. To investigate the molecular mechanism ofhigh expression in seed, the promoter ofwas obtained fromand was compared with the promoter offrom.The promoter ofwas cloned fromgDNA by FPNI-PCR, and its sequence was compared with the promoter of; The similar regions of the promoter were truncated, and the corresponding truncated sequences were obtained. Meanwhile, the GCN4 motif was mutated from the truncated sequence ofpromoter. The recombinant expression vectors of β-glucuronidase gene (GUS) driven by the above-mentioned promoter were constructed, and the-mediated method was used to infect the leaves offor transient expression to verify the promoter activities and the functions of GCN4 motif.The 365bppromoter sequence was cloned. The 3'end of 300 bp sequences was 93% similarity to the corresponding sequence of thepromoter. In this sequence, only thepromoter contained GCN4 motif. We successfully constructed a series of vectors fused with the reporter gene of GUS:(promoter full length),(promoter truncated to 320bp),(the mutantpromoter on GCN4 motif),(promoter full length) and(promoter truncated to 305 bp). Through GUS staining, it was found that all of the above-mentioned promoters have the activity of driving the transcription of GUS. However, promoters containing GCN4 motif, including full-length or truncatedpromoters (and) are more active than GCN4 motif mutation () or the promoters lack of GCN4 motif (and).This study improves GCN4-motif has a positive regulatory effect onand provides a basis for further exploration of the regulation mechanisms ofpromoter.
promoter; BPPS;Lour.;T. L. Wu; GCN4-motif; transient expression
R286.12
A
0253 - 2670(2022)19 - 6159 - 08
10.7501/j.issn.0253-2670.2022.19.021
2022-02-08
国家自然科学基金面上项目(81872954)
林晓静(1996—),女,硕士研究生,研究方向为药用植物次生代谢。Tel: 13724047090 E-mail: 517507071@qq.com
杨锦芬(1978—),女,研究员,研究方向为药用植物萜类化合物生物合成与调控。Tel: (020)39358331 E-mail: yangif@gzucm.edu.cn
[责任编辑 时圣明]
