动物黑色素形成及调控机制研究进展
2022-10-12赵净颖李丰耘豆腾飞贾俊静
赵净颖,李丰耘,豆腾飞,王 坤,黄 英,贾俊静*
(1.云南农业大学动物科学技术学院,云南昆明 650201;2.云南省西双版纳州勐腊县畜牧兽医站,云南西双版纳 666300)
黑色素(Melanins)是一种广泛存在于动、植物和微生物中的生物色素,是由酪氨酸和相关酚类化合物组成的一类广泛分布的酚类生物聚合物。动物黑色素主要分为真黑色素(Eumelanin)和脱黑色素(Pheomelanin)2 类,真黑色素由酪氨酸氧化生成的吲哚单体5,6-二羟基吲哚(DHI)和5,6-二羟基吲哚-2-羧酸(DHICA)组成,不含硫原子,呈棕色至黑色;脱黑色素由半胱氨酸多巴单体氧化生成的苯丙噻嗪类物质组成,含硫原子,呈黄色至微红棕色。动物黑色素主要沉着在头发、皮肤、角质层、羽毛和眼睛等,黑色的深浅主要由真黑色素和脱黑色素的相对数量决定。黑色素在皮肤中起美容、排毒和光保护作用(防辐射损伤),在掠食动物和猎物中起着伪装和防御的作用,在两栖动物和冷血动物中起着体温调节的作用;此外,黑色素还有抗氧化和抗病性等作用。目前,由黑色素形成的“乌质性状”已成为畜禽育种工作者们关注的重要性状。常见动物的乌质性状见表1。因此,研究黑色素形成、影响因素及调控,对阐明动物乌质性状形成机制意义重大。

表1 常见畜牧动物的乌质性状
1 黑色素的生成
黑色素是由黑色素细胞生成并且储存的,脊椎动物黑色素细胞几乎都是从神经嵴发源而来(除视网膜上皮细胞之外),神经嵴细胞在神经管闭合时向组织扩散,沿着2 条空间离散的路径迁移,神经嵴来源的细胞直到分散后才会经历明显的分化,并在精确的位置产生外周神经元、胶质细胞和黑色素细胞。神经嵴来源的黑色素细胞在胚胎发生过程中通过间充质向表皮和毛囊迁移。黑色素细胞大多数储存在真皮中,占表皮细胞的2%~4%,随着年龄逐渐增加而慢慢减少。
黑色素是一种由黑色素细胞分泌的酪氨酸酶(TYR)催化体内酪氨酸羟化合成的聚合物,拥有复杂结构。在黑色素细胞内,黑色素在称为黑素体的独特的、专门的膜结合细胞器中合成。黑色素生成开始于TYR催化的-酪氨酸合成多巴醌,并通过连续的氧化和脱羧中间步骤进行,最常导致真黑色素和脱黑色素2 种黑色素形式产生。具体表现为酪氨酸经TYR 催化转化为多巴(即3,4-二羟基苯丙氨酸,DOPA),DOPA在TYR 作用下进一步氧化为多巴醌(DQ),DQ 经多聚化反应与氧化反应生成多巴色素,多巴色素经过羧化和脱羧化形成中间体5,6-二羟基吲哚(DHI)和5,6-二羟基吲哚羧酸(DHICA),之后二者在TYR 及其相关蛋白的作用下形成中间产物5,6-吲哚醌(IQ)和5,6-吲哚醌羧酸(IQCA),最后2 种中间产物结合形成的异聚体即为真黑色素;而在酪氨酸氧化刚开始时形成的DQ 多了谷胱甘肽和半胱氨酸参与后,产生谷胱氨酰多巴和谷胱氨酰多巴醌,最后形成脱黑色素。其中,真黑色素和脱黑色素含量是由TYR 活力决定的,高活力的TYR 通过真黑色素的途径合成真黑色素,需要高活性的酪氨酸酶相关蛋白-1(TRP1)与酪氨酸酶相关蛋白-2(TRP2)参与,反之则合成脱黑色素。黑色素生成过程如图1 所示。
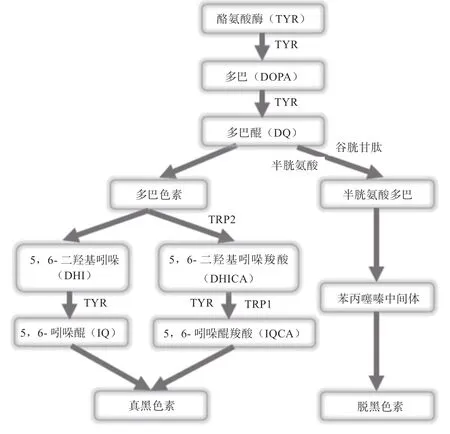
图1 黑色素生成过程
2 黑色素形成的影响因素
2.1 营养因素 由黑色素的合成可知,酪氨酸水平及TYR 活性对动物黑色素的作用极其重要,而日粮中酪氨酸含量可显著影响机体酪氨酸水平及TYR 活性。赵燕平等研究表明,1~4 周龄泰和乌骨鸡组织中黑色素含量随着日粮酪氨酸水平的提高先增加后降低,心脏、肌肉、皮肤中黑色素含量在1.18%酪氨酸水平时达到最高,而肝脏组织中黑色素的含量在0.98%酪氨酸水平达到最高。Li 等在淅川乌骨鸡基础日粮中分别添加0%(对照组)、0.2%、0.4%、0.6%、0.8% 和1.0% 的酪氨酸,饲养12 周后测定胸肌黑度及黑色素含量,结果表明,0.8%实验组胸肌中黑色素的沉积量最高。可见,在一定的酪氨酸水平范围内,提高日粮酪氨酸水平可提高泰和乌骨鸡组织中黑色素含量。除酪氨酸外,有研究发现日粮中添加微量元素可增加乌骨鸡组织中黑色素的含量。张倩雲等研究发现,日粮中添加适量的硒或酪氨酸均能提高凌云乌鸡血清与胸肌、肝脏、皮肤等组织中的黑色素含量及TYR 活性,并且硒与酪氨酸对于黑色素的含量存在互作效应。熊小文等研究表明,9~12 周龄泰和乌骨鸡肝脏、肾脏、胸肌和皮肤黑色素含量随日粮硒添加水平的提高呈先上升后下降的趋势。许兰娇等研究表明,在9~12 周龄泰和乌骨鸡饲粮中添加适量铜可显著或极显著影响其血清、肝脏、肾脏、皮肤及肌肉的黑色素含量、TYR 活性,并呈现出随饲粮铜添加水平提高而先升后降的趋势。此外,日粮中添加中草药可提高TYR 活性来增加乌骨鸡组织黑色素的含量。刘子暄等研究表明,中草药饲料添加剂可提高泰和乌骨鸡肌肉中的TYR 活力,从而促进其肌肉中黑色素的合成。
2.2 紫外线 紫外线可激活皮肤中的TYR,促使基底层黑色素细胞中的酪氨酸经过一系列反应生成黑色素,黑色素被黑色素细胞的树状分支输送到表皮细胞中,皮肤黑色素细胞形成黑色素以吸收紫外线,阻止紫外线进一步损害机体。在高紫外线辐射下,深色皮肤具有生物优势,可防止紫外线引起的晒伤、皮肤癌、免疫抑制和叶酸光解,叶酸是胚胎神经管发育和生育所必需的代谢产物;而在低紫外线辐射水平下,浅色皮肤具有生物优势,需要紫外线B(UVB)对应于皮肤中7-脱氢胆固醇转化为维生素D的有效波长,对健康产生多种影响,包括从骨代谢、固有免疫反应、细胞增殖和分化到生育等多方面。闫娟等研究表明,不同剂量UVB照射黑素细胞和表皮细胞后,部分黑素细胞和表皮细胞发生早衰,但酪氨酸介导的黑色素合成途径中TYR 活性升高和黑素细胞刺激表达增加,黑色素含量增加。周梦云等研究发现,与正常皮肤区域相比较,小型猪紫外线照射区皮肤的黑色素指数显著增高。
2.3 雌激素 基底皮肤中的黑色素细胞通过合成黑色素来控制皮肤色素沉着,而雌激素可增加黑色素细胞的色素生成。雌激素对黑素细胞具有增生作用,可能导致色素沉着过度。有研究报道,高雌激素会增加恶性黑色素瘤生成的风险;但也有研究表明这两者没有明显关联。唐建兵研究表明,雌激素对黄褐斑的发病可能具有促进作用,一定浓度的雌激素可以提高TYR 活性,促进黑色素细胞合成色素。Tang 等研究不同外源雌激素二乙基己烯雌酚(DES)浓度对黑素细胞增殖和TYR 活性的影响,结果表明向培养基中添加DES(10~10mol/L)增强了黑素细胞增殖,在浓度为10mol/L 时黑素细胞的TYR 活性显著增加。简丹研究表明,己烯雌酚对黑色素合成有上调作用,并呈浓度依赖性,且促黑色素合成可能与上调TYR 表达、增加其活性有关。
2.4 遗传因素 黑色素生成主要受到诸如种族的预先确定的遗传因素的影响,并且这一过程可以为动物机体提供保护和维持体内平衡。皮肤颜色会随物种的不同出现较大差异,对于个体而言,身体各个部位的颜色也有差异。Durairaj 等研究黑色素在人、猴、兔子、小型猪和狗模型中的眼内分布情况,结果表明在人类中,巩膜黑色素水平高于视网膜和中央脉络膜;在其他所有物种中,黑色素含量的等级顺序为:脉络膜RPE>视网膜≥巩膜。可见,黑色素含量在不同物种之间甚至同一物种不同组织区域之间都不同。徐方正等通过肌肉背最长肌研究表明,乌骨山羊肌肉组织中存在黑色素颗粒,而普通山羊组织中则没有,乌骨山羊肌肉内黑色素含量为0.416%,其含量是普通山羊的7.6 倍,两者差异极显著,可见乌骨山羊的“乌肉”特征是由于肌肉组织中含有大量黑色素而形成的乌质性状;乌骨山羊血清中的褐黑素、碱溶性黑色素含量与普通山羊并无显著差异,在肌肉组织中没有发现黑素细胞分布,由此推测黑色素经血液运输到肌肉组织中沉积,缓慢形成乌骨山羊肌肉的“乌色”特征。王高富等研究发现,酉州乌羊皮肤中真黑色素含量极显著高于板角山羊皮肤中真黑色素含量,说明羊皮肤真黑色素含量与皮肤颜色之间有一定相关性。五指山猪的背部皮肤呈黑色,腹部皮肤呈白色,Xu 等使用高通量RNA 测序比较五指山猪白皮肤和黑皮肤样本中编码和非编码RNA 的表达谱,功能分析发现黑色素A(),前黑素小体蛋白(),,多巴色素互变异构酶(),褪黑素()和钙/钙调素依赖蛋白激酶2()等调节黑色素生成的几个关键基因在两个皮肤组织中的表达水平明显不同,检查了潜在的lncRNA-miRNA-基因相互作用发现共有15 个lncRNA,11 个miRNA 和7 个基因形成了23 个lncRNA-miRNA-基因对,这表明五指山猪的肤色特征是编码和非编码基因调控的复杂网络。这些结果均表明黑色素存在种族差异性,且同一个体的不同部位也存在较大差异。
3 黑色素形成相关信号通路的研究
黑色素生成是一个非常复杂的过程,众多调控因子与信号通路相互作用,共同参与动物黑色素生成的调控。黑色素形成主要是由黑皮质素受体1(MC1R)/-黑素细胞刺激激素(-MSH)信号通路、环磷酸腺苷(cAMP)/蛋白激酶A(PKA)信号通路、丝裂原活化蛋白激酶(MAPK)/ 细胞外信号调节激酶(ERK)信号通路、磷酸肌醇3 激酶(PI3K)/蛋白激酶B(Akt)信号通路、Wnt/连环蛋白(-catenin)信号通路等参与调控的(图2)。
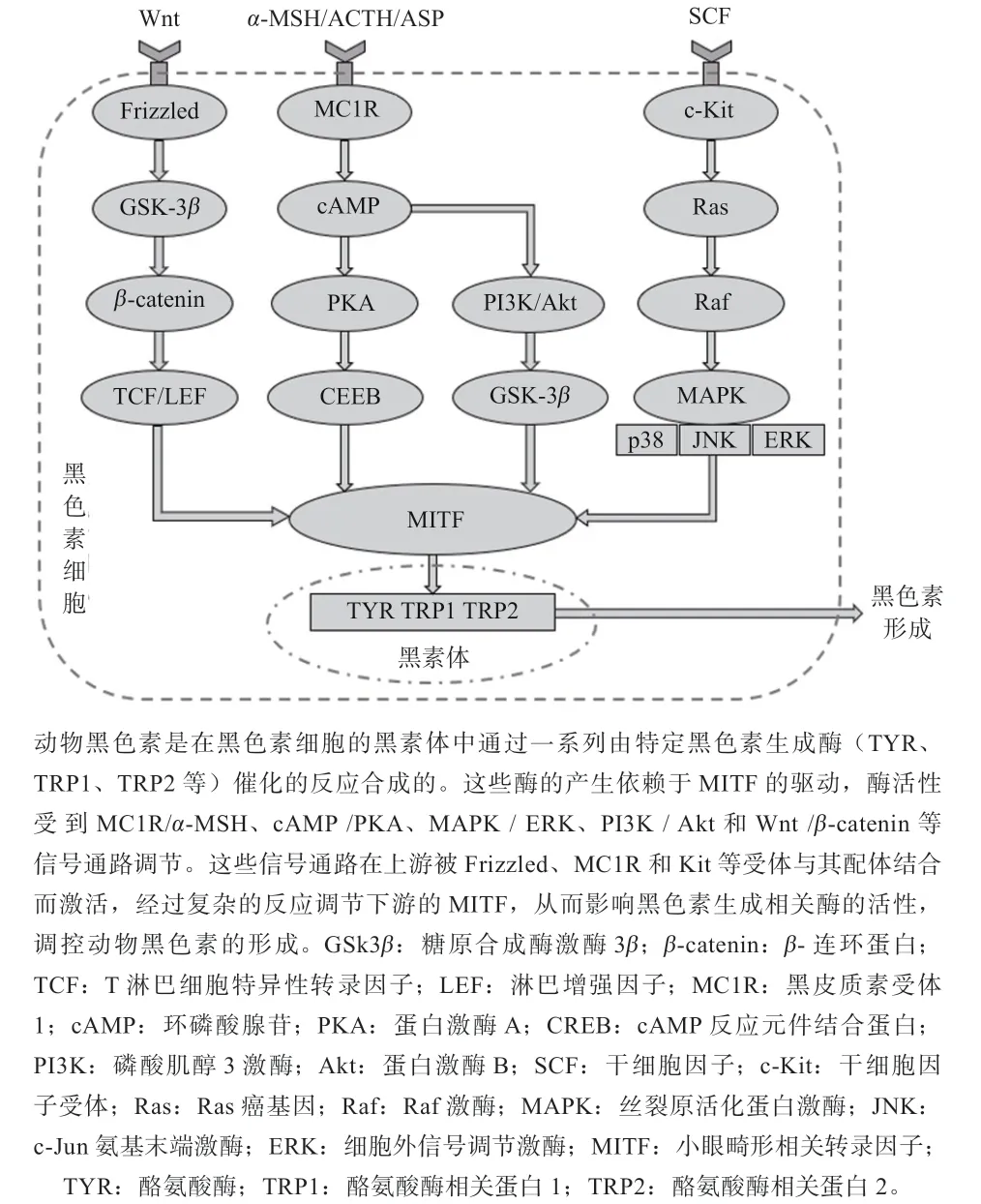
图2 动物黑色素生成相关信号通路网络图
3.1 MC1R/-MSH 信号通路是黑皮质素(MC)受体家族的成员之一,该家族还包括和等成员,这些成员具有不同的组织表达和功能。研究表明,与动物的毛色性状有关,可作为毛色选育的候选基因。-是可以诱导TYR 表达的重要因子,是垂体中间部细胞分泌的一种多肽激素,能调节小鼠黑色素瘤细胞中的表达和功能。存在于黑色素细胞中的,其激动剂包括-MSH,它可以激活腺苷酸环化酶,增加细胞内cAMP和激活cAMP 反应元件结合蛋白(CREB)。功能由激动剂-MSH 和促肾上腺皮质激素(ACTH)以及拮抗剂Agouti 信号蛋白(ASIP)控制。激动剂与的结合最终使TYR 活性增加,导致真黑色素合成;相反,当ASIP 与结合时,TYR 活性降低,导致脱黑色素产生。
在MC1R/-MSH 信号通路中,-MSH 结合MC1R激活腺苷酸环化酶产生cAMP,cAMP 激活PKA,并增加黑素细胞特异性小眼畸形相关转录因子()的表达,调节黑色素生成酶的水平,从而诱导黑色素的生成。其中,MITF 是黑色素细胞存活和增殖的主要正调节剂,在黑色素生成中起关键作用,可调节黑色素生成酶(TYR、TRP1 和TRP2)的表达。Almeida 等研究显示,人类头发黑色素的产生增加与和基因表达升高以及表达降低有关,随后也证实了在人类头发色素沉着中的关键作用。余丽莎研究显示,以窄波紫外线(NB-UVB)照射豚鼠背部裸露的皮肤后,豚鼠背部裸露区域呈明显的深棕黑色,色素均匀分布,黑色素颗粒含量大幅增长,且皮肤组织中-的表达量均增加;而经离子体射频技术(Plasma)与强脉冲光(IPL)治疗后,NB-UVB 所致的豚鼠背部皮肤的色素沉着均一定程度上减少,且豚鼠背部皮肤中-的表达降低,促进了NB-UVB 所致的豚鼠背部皮肤色素沉着的褪去;可见,豚鼠皮肤中-和的表达与皮肤的色素沉着存在一定的正相关关系。
3.2 cAMP/PKA 信号通路 诱导黑素生成的最重要的信号途径之一是cAMP/PKA 途径。-MSH 刺激黑素MC1R 通过G 蛋白信号激活腺苷酸环化酶,从而增加cAMP 的产生;cAMP 通过激活PKA 和cAMP 响应元件结合蛋白1 转录因子来上调的表达,后者控制黑素生成酶(和)mRNA 水平,从而调节黑色素的生成。郑嫩珠等研究表明,基因表达量在白绒乌鸡各组织间差异极显著,其中以皮肤表达量最高;各组织黑色素的沉积规律与各组织基因的表达规律大体相似,总体趋势表现为皮肤>肾>肌胃>肝>肌肉;且白绒乌鸡组织中基因的表达量与黑色素含量呈显著正相关。结果表明基因表达具有组织特异性,对白绒乌鸡体内黑色素沉积起正向调控作用,其高表达可能有助于乌鸡体内黑色素的形成。Yun 等研究表明,抑制CREB 的cAMP/ PKA 信号磷酸化,并抑制cAMP 调节的转录共激活因子1(CRTC1)的去磷酸化,下调-或基因的表达,可减少黑色素的沉着。Kim 等研究表明,通过激活cAMP/ PKA 信号传导和CREB 磷酸化并上调cAMP 信号传导激活的、和表达来刺激TYR 的活性和表达,从而可以刺激鼠B16F10黑色素细胞中黑色素的生成。Lee 等研究显示,白僵菌素可降低黑色素细胞的黑色素含量和TYR 活性,降低、TYR、TRP1 和TRP2 的蛋白水平,抑制cAMP-PKA-CREB 信号传导,通过调节cAMP/PKA 途径抑制黑色素生成,从而导致黑色素水平降低。可见,cAMP/PKA 信号通路的激活可调节黑色素的生成。
3.3 MAPK/ ERK 信号通路 MAPK 家族成员包括ERK、c-Jun 氨基末端激酶(JNK)和p38 MAPK。有研究表明,与哺乳动物的黑色素生成有关。刘学贤等研究表明,利用miR-146a 抑制其靶基因和的表达,可通过影响MAPK 级联路径中等重要因子的表达,进而调控羊驼黑色素细胞的增殖和迁移能力。在黑色素细胞中,可激活MAPK/ ERK 途径,从而通过调节的激活和稳定性来调控黑色素合成。cAMP 的上调激活了黑色素细胞中的MAPK/ ERK 途径,而该途径中ERK 或JNK 的活化(磷酸化)会触发的表达,导致其降解,从而减少黑色素生成。Kim 等研究表明,的磷酸化会抑制斑马鱼黑色素生成,而抑制的磷酸化可通过增加基因的表达,使和上调促进斑马鱼中黑色素的生物合成。Chae 等研究发现,下调、腺苷酸环化酶2、和的水平可减少黑色素产生,且PI3K/ Akt 和MAPK/ ERK 途径的激活可能起到抑制黑色素生成的作用。可见,MAPK/ ERK 途径激活可能负调控黑色素的生成。
3.4 PI3K/Akt 信号通路 PI3K/Akt 信号通路是一种重要的有丝分裂信号通路,在多种细胞过程中发挥着重要作用。近年来的研究表明,PI3K/Akt 信号通路对动物黑色素的生成具有调控作用。Shin 等研究表明,黑素细胞中核因子E2 相关因子2()的过表达导致PI3K/Akt 信号转导明显激活,降低了和基因的表达,从而导致黑色素数量显著减少;可见,PI3K/ Akt 信号传导途径的激活负调控黑色素生成。Moon 等研究显示,氯雷他定下调黑色素细胞中的和表达,且持续激活Akt,而PI3K/Akt 途径抑制剂LY294002 则解除了了氯雷他定引起的黑色素含量降低。综上所述表明,氯雷他定可能通过PI3K/Akt 信号导致黑素生成蛋白的抑制,调节黑色素生成。可见,PI3K/Akt 途径的激活可能对黑色素的生成发挥负调控作用。
3.5 Wnt/-catenin 信号通路 据报道,经典Wnt 信号,特别是和在黑色素细胞的发展中发挥重要作用。Wnt 信号通路参与调控的表达,Wnt 途径的激活抑制糖原合酶激酶3()的磷酸化并导致-的积累,积聚的-被转运到细胞核中,并与淋巴增强因子(LEF)/T 淋巴细胞特异性转录因子(TCF)形成复合物,之后引起表达增加;其中,参与调节黑色素生成,抑制活性可通过Wnt/-catenin 途径的激活增加黑色素合成。因此,Wnt/-catenin 信号通路的激活可能增加黑色素的产生。Su 等研究表明,MEK/ERK、PI3K/Akt 的激活 和Wnt/-catenin 信号通路的抑制组成了黑色素生成信号级联,没食子酸的激活可通过下调及其下游信号通路减少黑色素合成。Molagoda 等研究发现,非瑟酮(Fisetin)与GSK-3结合,可负性调节GSK-3,增加-catenin 的表达,导致黑色素生成;而Wnt/-catenin 抑制剂FH535 显著抑制了Fisetin 介导的斑马鱼幼虫黑色素生成。综上所述,Wnt/-catenin 信号通路的激活可能促进黑色素的生成。
4 结 语
动物的毛色性状及乌质性状是动物重要的经济及生产性状,黑色素在这些重要性状中起着主导作用。自然界中动物黑色素资源丰富,具有许多重要的生物学功能,对动物的生长发育及应对环境的影响至关重要。然而目前的研究中,黑色素生成的分子机制尚不明确,还需更多的研究来支撑。因而,本文对动物黑色素的形成及分子机制进行综述,以期为动物黑色素的研究提供理论依据。未来应着重研究外部因子对黑色素沉积的调控作用,为动物育种创造新思路,为治疗黑色素相关疾病(皮肤癌等)提供治疗策略。
