淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区浮游植物群落结构的时空格局及影响因子
2022-09-30王东伟龚江杨晓曦赵伟周彦锋
王东伟,龚江,杨晓曦,赵伟,周彦锋,*
(1.南京农业大学 无锡渔业学院,江苏 无锡 214081; 2.中国水产科学研究院淡水渔业研究中心 农业农村部长江下游渔业资源环境科学观测实验站,江苏 无锡 214081; 3.怀远县农业综合行政执法大队,安徽 蚌埠 233499)
浮游植物是水域生态系统的初级生产者,其作为水生动物的能量来源,是影响渔业生产的重要生物因素之一,浮游植物生长周期短、易受环境因子变化影响,其密度、生物量及时空分布特征均可作为评价水域生态系统环境因子变化的重要指标[1-2]。浮游植物是水生态系统的基础,也是水生生物食物链的重要环节,对水域生态系统物质循环、能量流动和水域生态系统稳定具有显著影响[3-4]。郭杰等[5]研究表明,长江荆江段浮游植物的种类组成、丰度和生物量具有明显的空间异质性,溶氧、温度和氨氮是影响浮游植物群落结构的主要环境因子。马煜等[6]研究表明,松花江哈尔滨段浮游植物群落结构对该河段生境的时空异质性响应明显,电导率、总磷、pH和浊度是驱动浮游植物群落结构时空分布的主要环境因子。因此,探究浮游植物群落结构不仅对水质理化因子监测具有重要意义,同时也可为制定水生生物资源保护措施与管理策略提供科学依据。
水产种质资源保护区对科学利用自然资源、维护生态平衡及保护生物多样性具有十分重要的意义。淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区(以下简称保护区)于2013年成立,位于安徽省怀远县淮河荆涂峡山口、涡河下游和茨淮新河下游。陈豪等[7]、张珊等[8]研究了淮河流域浮游生物及其环境影响因子,但针对该保护区浮游植物的生物学研究鲜见报道。本研究中,通过调查保护区浮游植物的群落结构组成,分析其结构特征、时空格局及其环境影响因子,以期为淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区的生态保护及生物完整性评价提供数据支撑,为动态调整增殖放流规划和制定水生生物保护规范提供科学依据。
1 材料与方法
1.1 研究区域和采样点
1.1.1 研究区域概况 保护区(117°05′39″~117°15′31″E,32°47′59″~33°00′03″N)全长为32.9 km,总面积为16.71 km2,其中,核心区面积为7.53 km2,试验区面积为9.18 km2。该保护区北至涡河入淮河口处,西至荆山,东至涂山,南至茨淮新河入淮河口处,包括淮河荆涂峡山口上下游及两条支流(涡河、茨淮新河),属温带半湿润季风气候,年平均气温为15.4 ℃,降雨量900 mm左右,年径流量为808.82 m3/s[9],沿河两侧以岗坡地、湖地和湾地为主,周边土地多用于农业种植和人口居住[10]。保护区保护对象除鲤和长吻鮠外,还包括青鱼、草鱼、鲢、鳙、刀鲚、黄颡鱼和鲇等。
1.1.2 采样站位及采样时间 根据保护区水域地形地貌和水文特征,共设置6个监测断面。断面1、2位于涡河,断面5、6位于淮河干流,断面3位于涡河与淮河交汇处,断面4位于茨淮新河和淮河交汇处,断面3、4属于保护区核心区水域。每个断面设置3个采样点,共计18个采样点(图1)。采样时间分别为2020年冬季(1月)、春季(4月)、夏季(8月)和秋季(10月),每个季节采集1次,共采集4次。

图1 保护区调查站位示意图Fig.1 Schematic diagram of survey sample stations in the reserve
1.2 方法


1.2.2 浮游植物样品的采集与鉴定 浮游植物样品按照《淡水浮游生物研究方法》[11]进行采集。采用25#浮游生物网(孔径75 μm,网袋入口直径20 cm,网锥体侧面动线长50 cm)采集定性样品,在水深0.5 m处以0.5 m/s的速度呈“∞”型拖拉5 min,带回实验室在10×40倍光学显微镜下观察分类。浮游植物鉴定主要依据《淡水微型生物与底栖动物图谱(第2版)》[12]和《中国淡水藻类:系统、分类及生态》[13]。
用有机玻璃采水器在水深0.5 m处采集样品 1 L,现场加入15 mL鲁哥试剂并摇匀,带回实验室静置沉淀24 h后,浓缩并定容至25 mL待镜检。用0.1 mL浮游植物计数框在10×40倍光学显微镜下进行计数。计数时充分摇匀浓缩液,取0.1 mL样品放入计数框中,观察100个视野。对量小而个体大的物种在10×10倍下全片计数。每个样品计数两片,取其平均值作为最终结果。若两片计数结果相差15%以上,则进行第三片计数,取其中个数相近的两片平均值。最后换算成每升水样中藻类的细胞个数,即为细胞数量(cells/L)。由于浮游植物的密度接近1,故可以直接由浮游植物的体积换算为生物量(湿质量),即生物量为浮游植物的数量乘以各自的平均体积(mg/L)。
1.2.3 浮游植物多样性及水质营养状态评价 浮游植物Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和 Margalef丰富度指数(D)计算公式分别为
(1)
J=H′/log2S,
(2)
D=(S-1)/log2S。
(3)
其中:S为浮游植物物种数;N为浮游植物个体总数;Ni为第i种浮游植物个体数。
浮游植物优势种属采用优势度(Y)描述,优势度计算公式为
Y=fi·Pi。
(4)
其中:fi为第i种浮游植物的出现频率;Pi为第i种浮游植物密度占总浮游植物密度的比例,当Y≥0.02时,定为优势种。
水质综合营养状态评价参照王明翠等[14]的方法。利用Chl-a、TP、TN、SD和CODMn指数计算得出,综合营养状态指数(TLI)计算公式为
(5)
其中:Wj为第j种参数的营养状态相关权重;TLI(j)为第j种参数的营养状态指数;m为评价参数的个数。
1.3 数据处理
采用SPSS 26.0软件对浮游植物生物量、密度、物种数和物种多样性数据之间的时空差异进行单因素方差分析(ANOVA)和成对样本T检验。采用Canoco 5软件进行冗余RDA(redundancy analysis)分析。为使数据符合正态分布,其优势种种群数据矩阵经过lg(x+1)转换,环境因子数据矩阵除pH外进行lg(x+1)转换,并对环境因子进行蒙特卡洛检验(Monte Carlo permutation test),选择出显著影响的因子,显著性水平设为0.05。
2 结果与分析
2.1 浮游植物种类组成
保护区调查期间,共鉴定到浮游植物8门71属143种,包括蓝藻门Cyanophyta 13属23种、甲藻门Pyrrophyta 2属2种、硅藻门Bacillariophyta 15属28种、隐藻门Cryptophyta 2属3种、裸藻门Euglenophyta 3属13种、绿藻门Chlorophyta 33属71种、金藻门Chrysophyta 2属2种和黄藻门Xanthophyta 1属1种。其中,绿藻门在浮游植物物种数上有显著优势,占总物种数的49.65%;其次为硅藻门和蓝藻门,分别占19.58%和16.08%,黄藻门最低,占0.70%。浮游植物种类呈现夏季(101种)>春季(58种)=秋季(58种)>冬季(41种)。从空间格局来看,试验区和核心区的浮游植物物种数无显著性差异(P>0.05),但涡河水域的裸藻门、绿藻门物种数均显著高于淮河干流水域(P<0.05)。不同季节各采样断面浮游植物物种数占总物种数的比例如图2所示。

图2 不同季节各采样断面浮游植物物种数占比Fig.2 Proportion of phytoplankton species in each sampling section in different seasons
2.1.1 优势种种类组成 保护区浮游植物优势种共有13种(表1),其中,蓝藻门和绿藻门各4种,硅藻门和隐藻门各2种,金藻门1种。优势种数量呈现秋季>夏季=冬季>春季的季节变化特征,且夏季优势种多为蓝藻门,优势种的优势度峰值主要呈现春夏季蓝藻门、秋季硅藻门、冬季蓝藻门的变化特征。梅尼小环藻Cyclotellameneghiniana、尖尾蓝隐藻Chroomonasacuta和衣藻Chlamydomonassp.在全年的优势较明显,在3个季度均形成优势。

表1 不同季节浮游植物优势种及优势度Tab.1 Dominant species and dominance of phytoplankton in different seasons
2.1.2 群落结构多样性 从表2可见:保护区浮游植物Shannon-Wiener多样性指数变动范围为1.48~3.08,平均值为2.41±0.44;Pielou均匀度指数为0.46~0.96,平均值为0.78±0.14;Margalef丰富度指数为0.48~2.58,平均值为1.55±0.59。Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数在空间格局上无显著性差异(P>0.05)。从时间上来看,Shannon-Wiener多样性指数春季和冬季的平均值显著低于夏季和秋季(P<0.05),呈现夏季最高、春季最低的态势;Pielou均匀度指数在季节变化中无显著性差异(P>0.05);Margalef丰富度指数冬季的平均值显著低于其他3个季节(P<0.05),春季和秋季的平均值显著低于夏季(P<0.05),呈现夏季最高、冬季最低的态势。

表2 不同季节各断面浮游植物多样性指数变化Tab.2 Changes in phytoplankton diversity index at various sections in different seasons
2.1.3 细胞密度与生物量 从表3和图3可见:保护区4个季节浮游植物细胞密度为9.84×104~7.86×107cells/L,其中春季浮游植物密度主要以绿藻为主,平均细胞密度为5.87×106cells/L,占比为28.36%;4个季节浮游植物生物量为0.02~29.07 mg/L,平均生物量为3.70 mg/L;保护区浮游植物密度和生物量在季节上无显著性差异(P>0.05)。从空间格局来看,试验区(断面1、2、5、6)与核心区(断面3、4)的浮游植物密度及生物量均无显著性差异(P>0.05),但涡河水域(断面1、2)浮游植物密度和生物量均显著高于淮河(断面3、4、5、6)干流水域(P<0.05)。

表3 不同季节各断面浮游植物密度及生物量变化Tab.3 Changes in the density and biomass of phytoplankton at various sections in different seasons
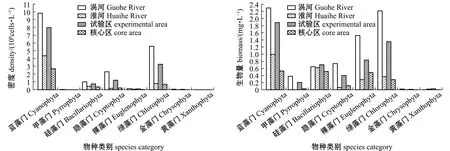
图3 不同区域浮游植物密度和生物量Fig.3 Phytoplankton density and biomass in different regions
2.2 水质营养状态及理化因子
保护区水质综合营养状态指数在季节上表现为春、夏和冬季处于轻度富营养化状态,秋季处于中度富营养化状态(表4)。空间格局上,保护区水域均处于轻度富营养化状态。保护区4个季节均处于磷限制状态。各季节不同采样断面水质理化因子数值变化情况见表5。

表4 水质综合营养状态指数Tab.4 Water quality comprehensive trophic state index

表5 全年水环境因子平均值Tab.5 Average value of annual water environment factors
2.3 浮游植物群落结构与环境因子的关系

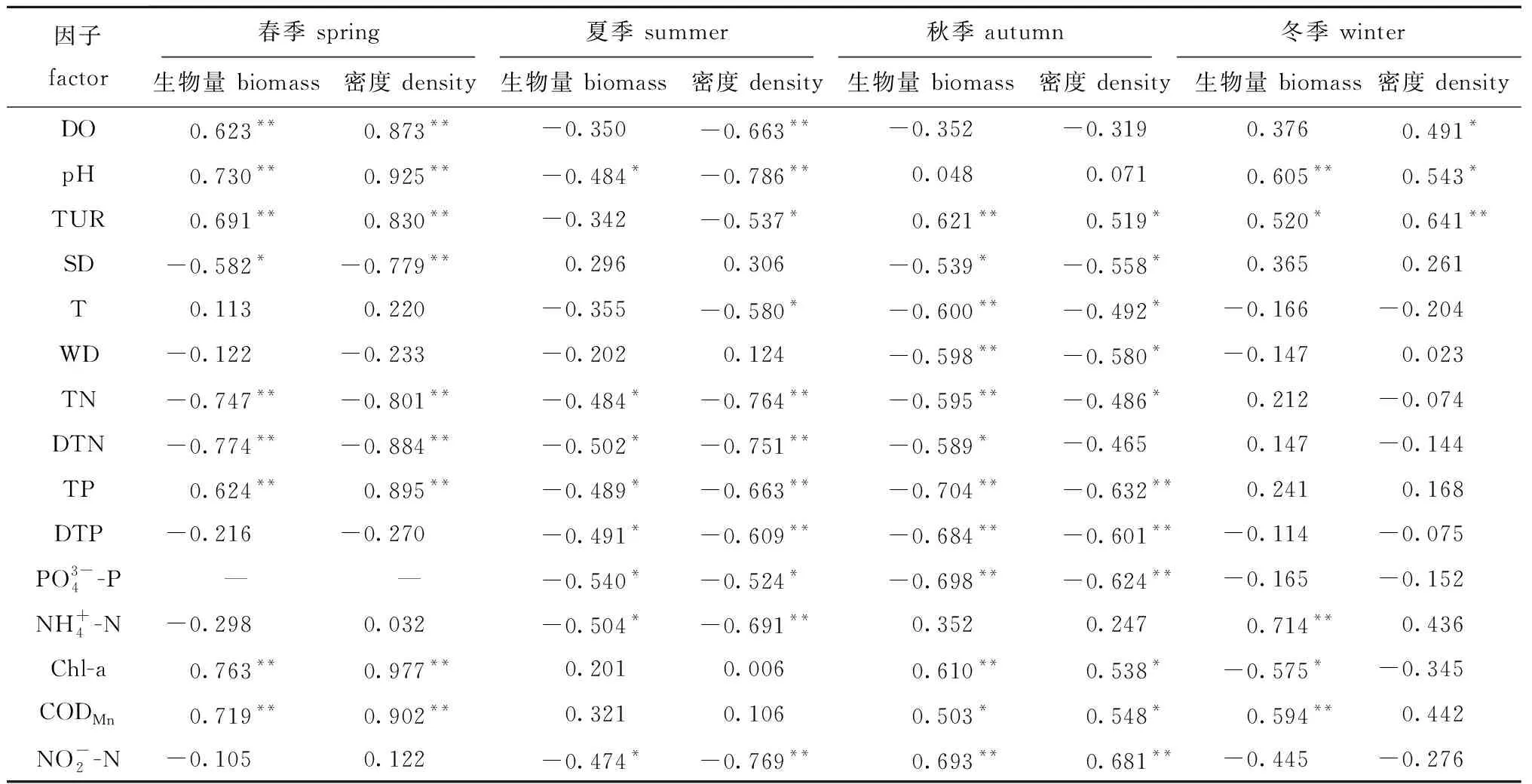
表6 浮游植物密度和生物量与环境因子的相关性分析(相关系数R)Tab.6 Correlation analysis between phytoplankton and environmental factors(correlation coefficient R)
趋势对应分析(DCA)显示,4个季节排序轴长度最大值为1.54(<3),故适用基于线性的RDA分析。经蒙特卡洛置换检验对水质环境因子进行筛选,春、夏、秋和冬季分别筛选出9、12、5和7个与浮游植物呈显著性相关的环境因子。选取浮游植物优势种与筛选出的环境因子进行RDA分析,结果如图4所示。春季RDA结果显示,前两轴累计变异百分率解释了物种数据的89.72%,环境因子对浮游植物密度解释了91.50%,主导环境因子是SD,解释了77.60%的浮游植物群落信息;夏季RDA结果显示,前两轴累计变异百分率解释了物种数据的81.77%,环境因子对浮游植物密度解释了86.60%,主导环境因子是TN,解释了71.80%的浮游植物群落信息;秋季RDA结果显示,前两轴累计变异百分率解释了物种数据的34.39%,环境因子对浮游植物密度解释了42.80%,主导环境因子是WD,解释了15.00%的浮游植物群落信息;冬季RDA结果显示,前两轴累计变异百分率解释了物种数据的46.30%,环境因子对浮游植物密度解释了55.00%,主导环境因子是CODMn,解释了25.70%的浮游植物群落信息。

图中物种代码见表1。See Table 1 for species codes in the figure.图4 各季节浮游植物优势种与环境因子RDA分析Fig.4 RDA analysis of dominant species of phytoplankton and environmental factors in each season
3 讨论
3.1 保护区浮游植物群落结构分析
本研究中,在淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区共鉴定出浮游植物8门71属143种,高于赵秀侠等[15]在2015年对淮河干流安徽段浮游植物的调查结果(6门35属57种),与其在2012年的调查结果(8门61属147种)[16]基本一致。3次调查结果中,浮游植物的群落结构存在相似性,如主要浮游植物占据优势地位的物种均为蓝藻门、硅藻门和绿藻门,浮游植物密度均呈现蓝藻门>绿藻门>硅藻门。然而,本次调查中蓝藻门的生物量占比在快速增长,其原因可能是淮河流域极高的人口密度及庞大的水系组成,带动沿岸工农业发展和河流输入,致使流域内高浓度的营养盐水体刺激蓝藻增殖[17],进而导致蓝藻生物量增加。
浮游植物物种组成和群落结构受水体营养盐浓度、水动力学特征、浮游动物觅食压力和水文动态的直接作用,除此之外,时间与地理空间变化的影响也不容忽视[18]。水温的季节性差异、地区水文特征和水域受污染状况,可能是造成本研究中保护区浮游植物群落结构的优势种呈现春夏季蓝藻门、秋季硅藻门、冬季蓝藻门变化特征的主要因素。其中,水温会对浮游植物的群落种类和分布格局造成影响[19],藻类种类不同,其适宜生存的环境也不尽相同,硅藻适宜在16~26 ℃生长[20],绿藻适宜在18~30 ℃生长[21],蓝藻适宜在25~35 ℃生长[22]。本研究中,保护区夏季平均水温为30.59 ℃,持续的高温为蓝藻的快速生长提供了良好的环境,虽然绿藻在温度较高时也可以快速生长,但在持续的高温环境中,蓝藻可以产生藻毒素并抑制绿藻增殖[23]。同时,保护区流域暴雨及洪水集中在汛期6—9月,暴雨会加剧周边农田和工业生产污水对水体的影响[24],水流量的增加也会将水体中营养盐类物质重新混合,致使水体的营养物质浓度升高,满足蓝藻生长的营养需求。相关研究表明,硅藻各优势属的生长具有极强的季节性,由于硅藻硅质外壳较重及缺乏鞭毛而容易在水体中沉降,因此,在湖泊水体混合期间硅藻容易占据优势[25],一般存在于低流速、高营养盐和适中温度的春季至初夏及高温之后的秋季[26],这与本研究中保护区硅藻群类结构相符合。有研究显示,淮河流域中沙颍河及涡河水体污染较为严重,淮河流域内多数样点处于中-重污染状态,隐藻在淮河流域分布广泛且占比较高,在受到一定有机污染的环境中形成优势种[27-28]。因此,保护区浮游植物中隐藻门长期形成优势可能与该水域受污染状况有关。
从空间格局来看,本研究中,试验区与核心区的浮游植物群落结构无显著性差异,但涡河流域浮游植物生物量、密度,以及裸藻门、绿藻门物种数均显著高于淮河干流。其主要原因可能是涡河闸坝的建设过于密集,虽然实现了水资源的高效利用,但影响了水体交换,导致营养盐类富集,给藻类的增殖提供了物质基础[27]。此外,保护区水域综合营养状态指数的空间变化与浮游植物生物量、密度有显著相关性(数据未列),由此可见,水文和水质变化对浮游植物的群落结构具有重要影响。
3.2 保护区浮游植物群落分布与环境因子的关系

4 结论
1)淮河荆涂峡鲤、长吻鮠国家级水产种质资源保护区浮游植物物种主要由蓝藻门、硅藻门和绿藻门构成,优势种以蓝藻门和绿藻门为主。浮游植物密度及生物量在时间格局上无明显差异,但空间格局上涡河显著高于淮河干流,应重点关注涡河水域的浮游植物生长状况,以防止蓝藻和绿藻过量增殖,从而影响保护区水生态健康。
2)春、夏、秋和冬季影响保护区浮游植物群落结构的主要环境因子分别为SD、TN、WD和CODMn,对不同时期的相应环境因子应予以重视,以维护浮游植物多样性。
3)保护区流域内水质均处于轻度富营养化状态,因此,需持续关注保护区的水环境状态,以防止水域污染。
