甘薯KUP/HAK/KT基因家族的全基因组鉴定和表达模式分析
2022-09-30吴胜男张永光吉林省农业科学院经济植物研究所吉林公主岭3605扎赉特旗音德尔镇综合保障和技术推广中心内蒙古兴安盟37600
吴胜男,孙 凯,张 海,刘 峰,张永光,王 凤(.吉林省农业科学院经济植物研究所,吉林 公主岭 3605;.扎赉特旗音德尔镇综合保障和技术推广中心,内蒙古 兴安盟 37600)
【研究意义】钾是植物生长不可或缺的大量营养元素之一,占植物总干物质的2%~7%。甘薯作为典型喜钾作物,钾素有调节甘薯细胞渗透压[1]、膜内外电荷平衡[2]、参与酶活反应[3]、提高光合反应速率[4],促进植株内糖分的运输和蛋白质合成等作用。研究表明通过栽培技术等手段可以提高甘薯对钾的利用效率[5],进而促使甘薯产量的提升[6],而有关甘薯钾离子吸收分子机制的研究还未见报道。因此,研究提高甘薯对K+的吸收和转运成为当下的热点方向。【前人研究进展】植物通过钾离子通道和转运体两大系统来完成植物对K+的吸收和转运,根据前人在拟南芥和水稻中的相关研究[7],已经确定K+转运体主要分为KUP/HAK/KT、KEA、Trk/HKT及CHX四大家族[8]。其中KT/HAK/KUP是在大肠杆菌中被首次鉴定到,称为KUP转运体,负责钾离子吸收,在土壤酵母中发现则称为HAK1转运体,对钾离子有较高的亲和力[9]。水稻的OsHAK5基因在过表达的情况下,能通过K+从地下部分到地上部分的转运,增加其耐盐性;在干旱胁迫胁迫下ABA可以拟南芥的KUP6基因表达量迅速上调;拟南芥AtHAK5和AtKUP11可以通过对K+的转运响应盐胁迫。【本研究切入点】至今在拟南芥[10]、水稻[11]、玉米[12]、小麦[13]、大豆[14]、苹果[15]中KUP/HAK/KT基因家族均已被报道,且有部分KUP/HAK/KT基因已被克隆和功能验证。甘薯是我国第四大的粮食作物[16],但在甘薯中相关研究尚未开展。【拟解决的关键问题】本研究中以甘薯品种‘泰中6号’全基因测序结果为基础,通过基因组学和转录组学对数据进行挖掘,对甘薯KUP/HAK/KT家族基因进行鉴定,分析其家族蛋白质的理化性质、亲缘关系和表达模式,为进一步深入分析甘薯KUP/HAK/KT基因功能奠定基础。
1 材料与方法
1.1 甘薯KUP/HAK/KT家族成员鉴定
首先通过甘薯基因组网站(https://ipomoea-genome.org/)得到甘薯全基因组蛋白序列、CDS和注释文件。其次以KUP/HAK/KT结构域(PF02705)的隐马尔科夫模型为条件,通过hmmsearch对甘薯蛋白质数据库进行搜索,筛选含有该结构域的序列。最后利用NCBI-CDD和Pfam数据库检测所得序列结构域的完整性,去除重复及不完整序列。在cello(http://cello.life.nctu.edu.tw/)进行亚细胞定位;ExPASy(http://web.expasy.org/protparam/)进行蛋白质的理化特性分析[17];利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)进行跨膜结构域预测[18]。
1.2 系统发育进化分析
从TRAI网站(https://www.arabidopsis.org/)和MSU数据库(http://rice.uga.edu/)分别下载拟南芥和水稻KUP/HAK/KT蛋白序列。利用MUSCLE进行多序列比对,利用MEGA 7.0软件比邻法(Neighbor-joining method,NJ)构建系统发育树,bootstrap值设为1000[19]。通过MEME服务器(http://meme-suit/org/)分析蛋白序列的保守基序(motifs),结合进化树分析结果整合制图。
1.3 启动子顺式作用元件和GO注释分析
利用eggNOG-mapper数据库(http://eggnog-mapper.embl.de/)对甘薯KUP/HAK/KT蛋白进行GO注释分析[20],使用WEGO 2.0(http://wego.genomics.cn/)进行可视化[21],对KUP/HAK/KT基因进行功能注释。利用PlantCARE软件对顺式元件进行分析。
1.4 染色体定位和基因复制事件分析
利用基因组注释文件,提取甘薯KUP/HAK/KT基因的位置信息,通过blastn比对甘薯基因组具有潜在重复关系的基因对,并用TBtools进行可视化[22]。使用KaKs_calculator计算甘薯KUP/HAK/KT基因Ka/Ks比值[23]。
1.5 表达模式分析
利用NCBI-SRA数据库获取的‘徐薯18’不同组织和逆境胁迫的转录组文件,使用hisat2和ballgown进行转录组数据分析,计算转录本TPM表达量,并进行KUP/HAK/KT基因家族在不同组织和逆境胁迫下的热图分析。
2 结果与分析
2.1 甘薯KUP/HAK/KT基因家族成员鉴定
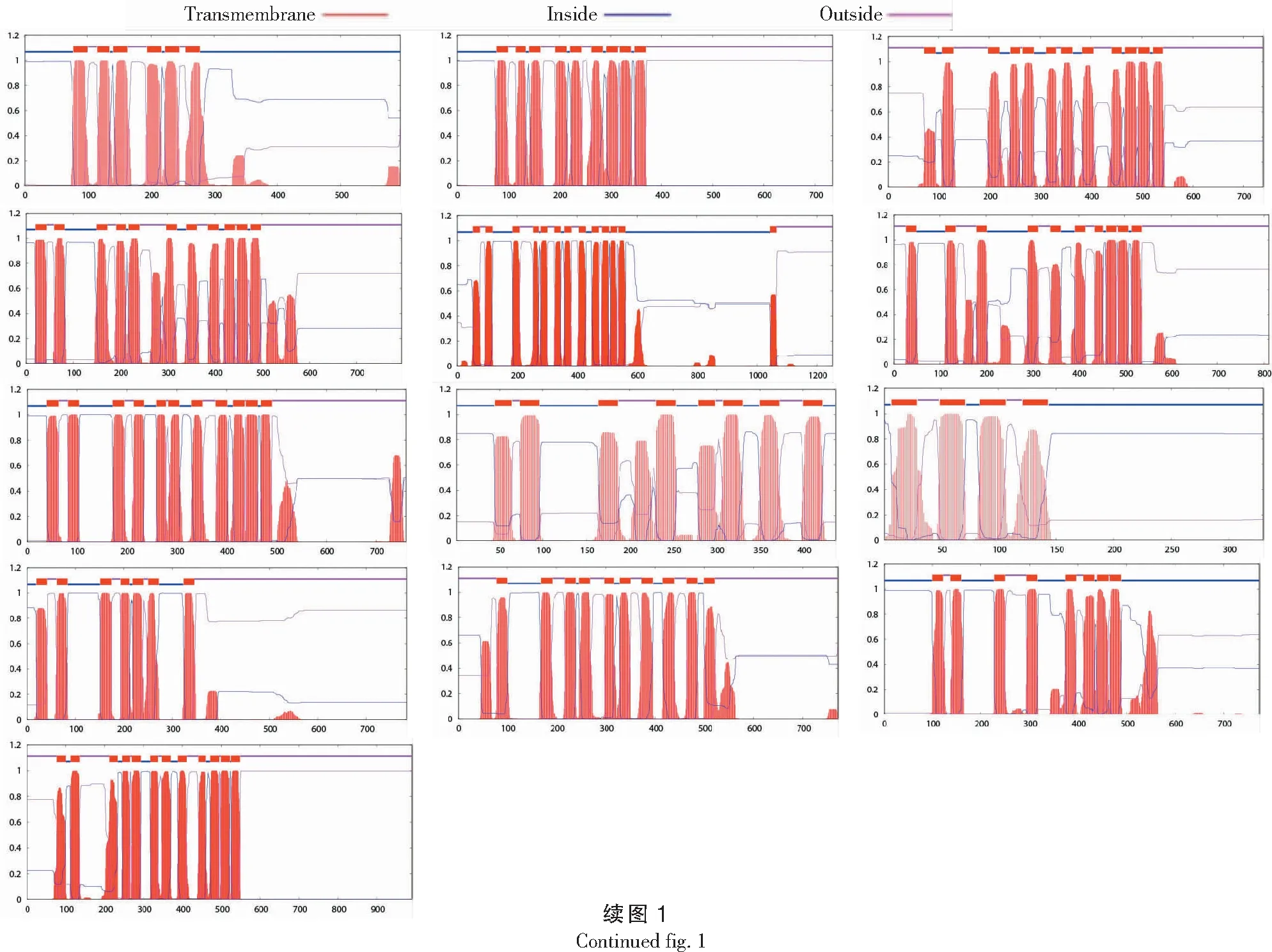
由表1可知,通过相关筛选和验证,在甘薯基因组中最终得到的22条甘薯KUP/HAK/KT候选蛋白序列。对其蛋白质理化特性分析发现,甘薯KUP/HAK/KT蛋白序列长度为227~1252 aa,平均长度为726.4 aa;分子量为18 297.95~140 640.46 u,平均值为78 242.8 u;等电点为5.54~9.31,平均值为8.18。基因的亚细胞定位对于其功能具有借鉴作用,结果表明所有的甘薯KUP/HAK/KT基因都定位于细胞质膜上,这和其主要作用是参与K+转运是一致的。蛋白完整的二级结构均具有4~13个跨膜区(图1)。
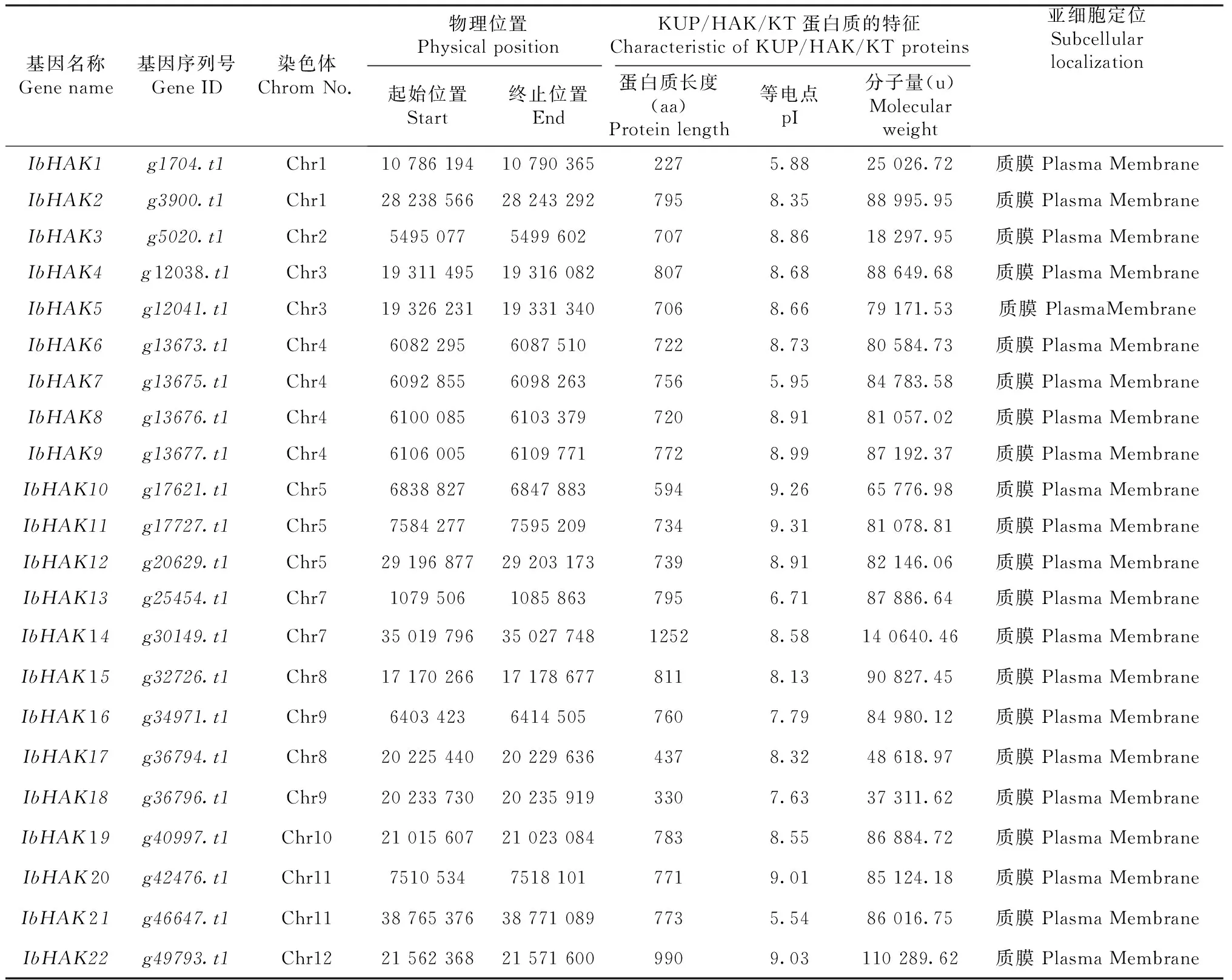
表1 甘薯KUP/HAK/KT家族基因的特征

纵坐标代表跨膜结构概率The ordinate represents transmembrane helices probability图1 甘薯KUP/HAK/KT蛋白的跨膜结构分析Fig.1 Transmembrane topology analysis of sweetpotato KUP/HAK/KT proteins
2.2 系统发育树的构建和蛋白质保守结构域分析
对13个拟南芥,27个水稻,22个甘薯KUP/HAK/KT全长蛋白质序列整合,利用MEGA 7软件中的NJ法构建系统发育进化树。如图2所示,甘薯KUP/HAK/KT基因家族分为4个进化簇,依次命名为ClusterⅠ、ClusterⅡ、ClusterⅢ和ClusterⅣ,又进一步细分为Ⅰa、Ⅰb、Ⅱa、Ⅱb、Ⅲ和Ⅳ 6个亚类。其家族基因数量依次为ClusterⅡ的成员数目最多(10),其次是ClusterⅠ(7)、再次是ClusterⅢ(3),ClusterⅣ的成员数目最少(2),表明甘薯KUP/HAK/KT家族基因的分布情况与拟南芥和水稻相似,均表现为ClusterⅠ和ClusterⅡ的成员数目远多于ClusterⅢ和ClusterⅣ。根据系统进化树,KUP/HAK/KT基因在水稻,拟南芥,甘薯中的同源性很高,表明KUP/HAK/KT基因物种分化过程中非常保守。
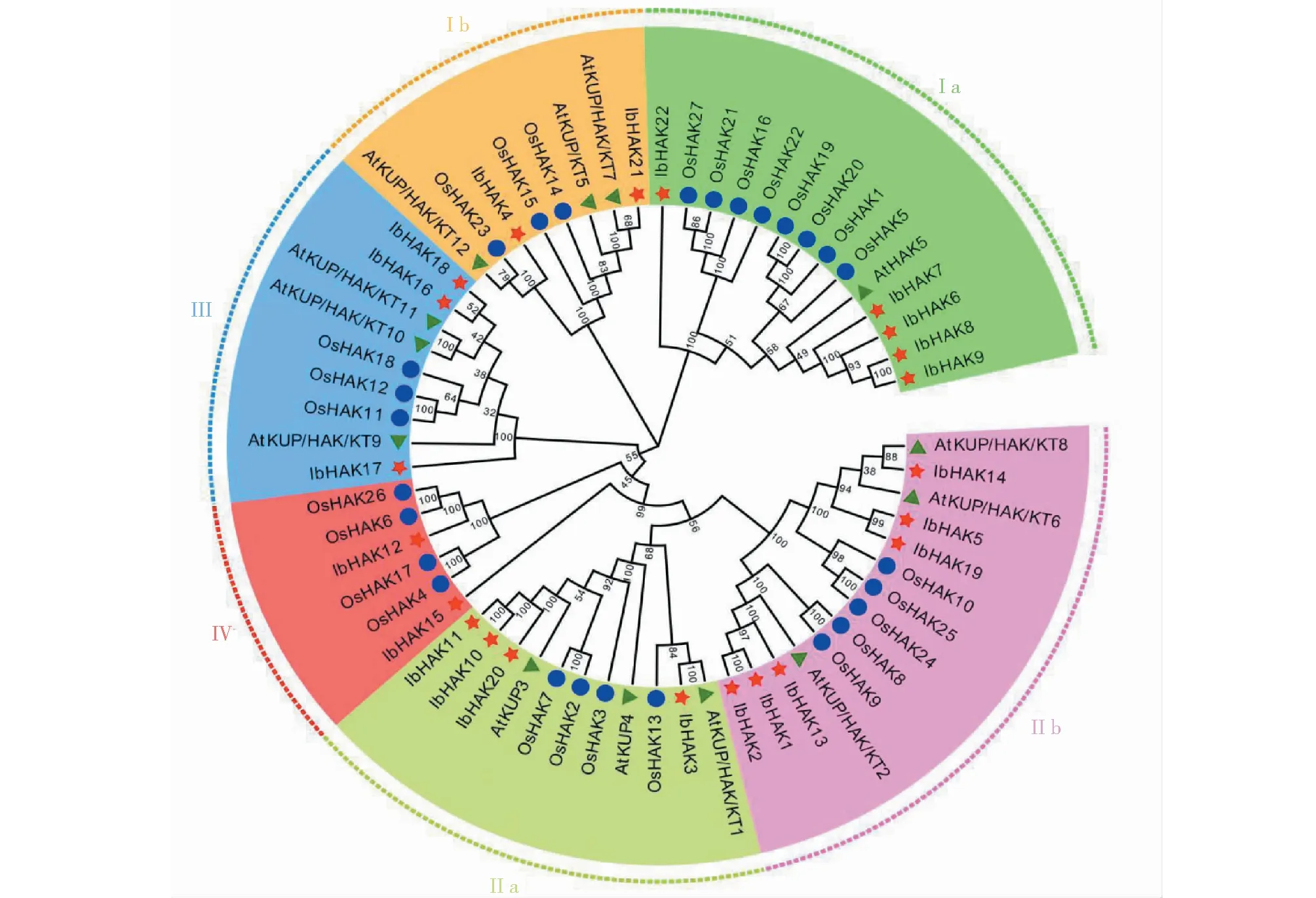
图2 甘薯、水稻和拟南芥KUP/HAK/KT基因家族系统进化树Fig.2 Phylogenetic tree of KUP/HAK/KT genes in sweetpotato, rice and Arabidopsis
如图3所示,在甘薯KUP/HAK/KT基因家族中共找到10个基序。进一步分析发现ClusterⅠ家族成员均含有基序4、基序5和基序8,ClusterⅡ均含有基序2、基序3、基序4、基序6和基序10,ClusterⅢ均含有基序8、基序9和基序10,ClusterⅣ均含有基序8和基序9。这表明,KUP/HAK/KT在进化过程中,存在着家族内部的分化,进一步可能导致各家族基因功能的分化。

图3 甘薯KUP/HAK/KT基因家族的保守基序Fig.3 Conserved motifs of KUP/HAK/KT genes in sweetpotato
2.3 GO注释和启动子顺式作用元件分析
从图4可以看出,GO注释主要包括了细胞成分(Cellular Component)、分子功能(Molecular Function)、生物过程(Biological Process)三部分。对甘薯KUP/HAK/KT基因进行GO分析;在细胞成分类别中,所有的KUP/HAK/KT蛋白在细胞膜的合成过程中发挥作用,25%的蛋白在细胞器的合成方面起作用。在分子功能类别中,100%的KUP/HAK/KT蛋白具有转运蛋白活性,来发挥其在植物体内的生物学功能;仅有少于10%的蛋白起催化作用。在生物过程类别中,发现有100%的蛋白在细胞进程中发挥作用、另有少于20%的KUP/HAK/KT蛋白可在生长发育过程和细胞组成生物合成过程中起到调控作用。通过上述结果表明甘薯KUP/HAK/KT蛋白在细胞膜组成过程和跨膜转运中发挥重要作用。

图4 IbKUP/HAK/KT蛋白的基因本体(GO)注释分析Fig.4 Gene Ontology annotation of IbKUP/HAK/KT proteins
顺式作用元件位于基因的旁侧,可以与功能基因一起发挥作用。本研究截取了IbHAK基因上游1500 bp的序列,分析基因的顺式作用元件。如图5所示,在IbHAK启动子区域共鉴定到12种顺式作用元件,大致可分为四大类:调控植物生长发育相关元件、光响应元件、逆境响应元件和激素响应元件。其中22个IbHAK基因都含有TATA和CAAT控植物生长发育相关元件以及光响应元件;22个IbHAK基因含有1~3种逆境响应元件,包含响应干旱、低温、厌氧、物理伤害的顺式作用元件;22个IbHAK基因都含有1~4种激素响应元件,包含响应茉莉酸甲酯、脱落酸、生长素、赤霉素和水杨酸顺式作用元件。这表明IbHAK基因对逆境和激素响应中起到至关重要的作用。
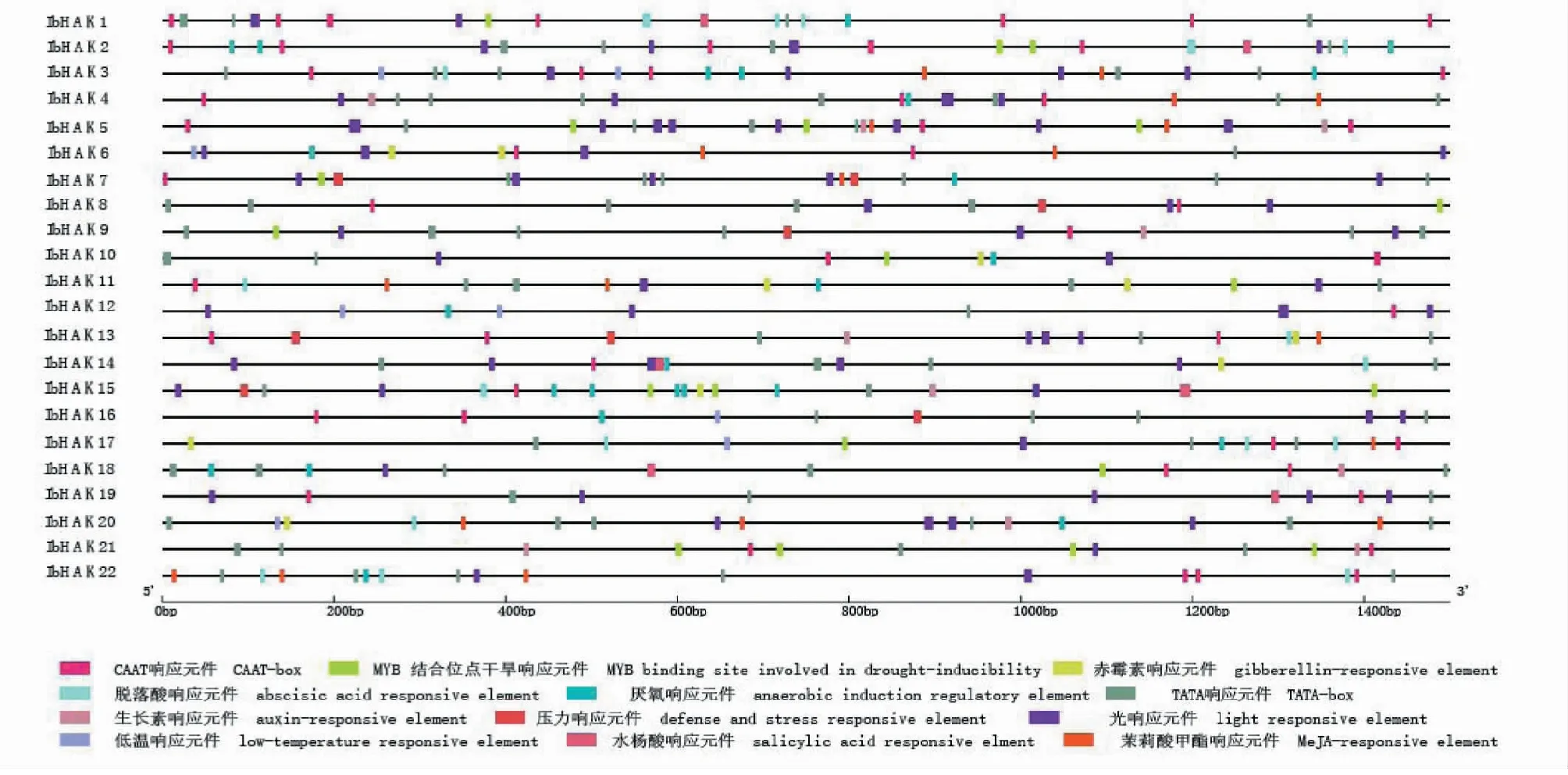
图5 甘薯KUP/HAK/KT启动子顺式作用元件分析Fig.5 Cis-elements in the promoter regions of KUP/HAK/KT genes in sweetpotato
2.4 染色体定位、基因复制分析
根据甘薯基因组gff3注释文件,将鉴定得到了IbHAK基因进行染色体定位。如图6所示,22个基因分布在甘薯的11条染色体上。其中Chr4染色体上含有IbHAK基因最多(4个);Chr5染色体上分布3个IbHAK基因;而Chr6、Chr13、Chr14和Chr15 4条染色体上没有IbHAK基因;此外的9条染色体上分布1~2个IbHAK基因。通过对甘薯IbHAK复制事件的分析,共发现3对大片段复制事件(IbHAK1/IbHAK2、IbHAK2/IbHAK13和IbHAK10/IbHAK11)和2对串联复制事件(IbHAK7/IbHAK8和IbHAK8/IbHAK9)。为了研究IbHAK基因在进化过程中对受到何种选择,计算了非同义突变频率(Ka)和同义突变频率(Ks)的比值,发现IbHAK基因的并联复制和串联复制基因对的Ka/Ks值分别为0.75和0.893,比值均小于1,说明所有IbHAK复制事件在甘薯进化过程中都受到了纯化选择(表2)。
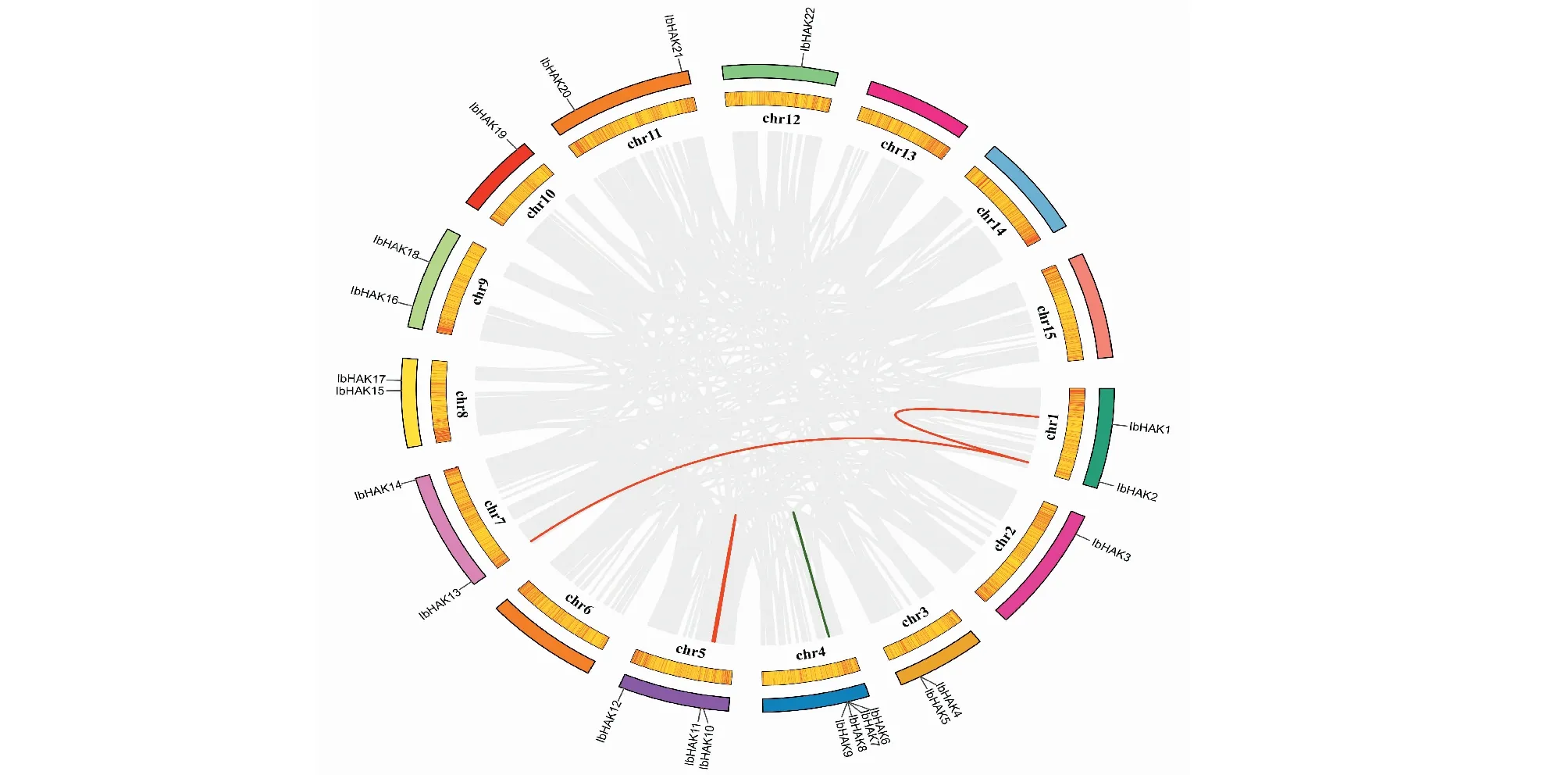
图6 甘薯KUP/HAK/KT基因的基因定位和甘薯基因组内复制基因对Fig.6 Genomic locations of IbKUP/HAK/KT genes and duplicated gene pairs in sweetpotato

表2 甘薯KUP/HAK/KT基因复制事件类型、同义替换率、非同义替换率及其比值
2.5 表达模式分析
为深入研究IbHAK基因在甘薯生长发育过程中潜在的生物学功能,通过‘徐薯18’的转录组数据对甘薯KUP/HAK/KT家族基因在根、茎和叶中的表达情况进行分析(图7-A)发现:IbHAK基因表现出了明显的组织表达特异性,一些组织特异性表达的基因主要分布在Ⅱa、Ⅲ、Ⅱb和Ⅰa,其中亚家族Ⅱa和Ⅲ全部成员,Ⅱb大部分成员均在茎中呈现高表达,在叶和根中低表达;亚家族Ⅰa的大部分成员在根中表达量较高,在叶和茎中表达量很低。Ⅰa中IbHAK22、IbHAK6、IbHAK8和IbHAK9在根中的表达量远高于茎和叶,这几个IbHAK基因在甘薯根的生长发育过程中起至关重要的作用。
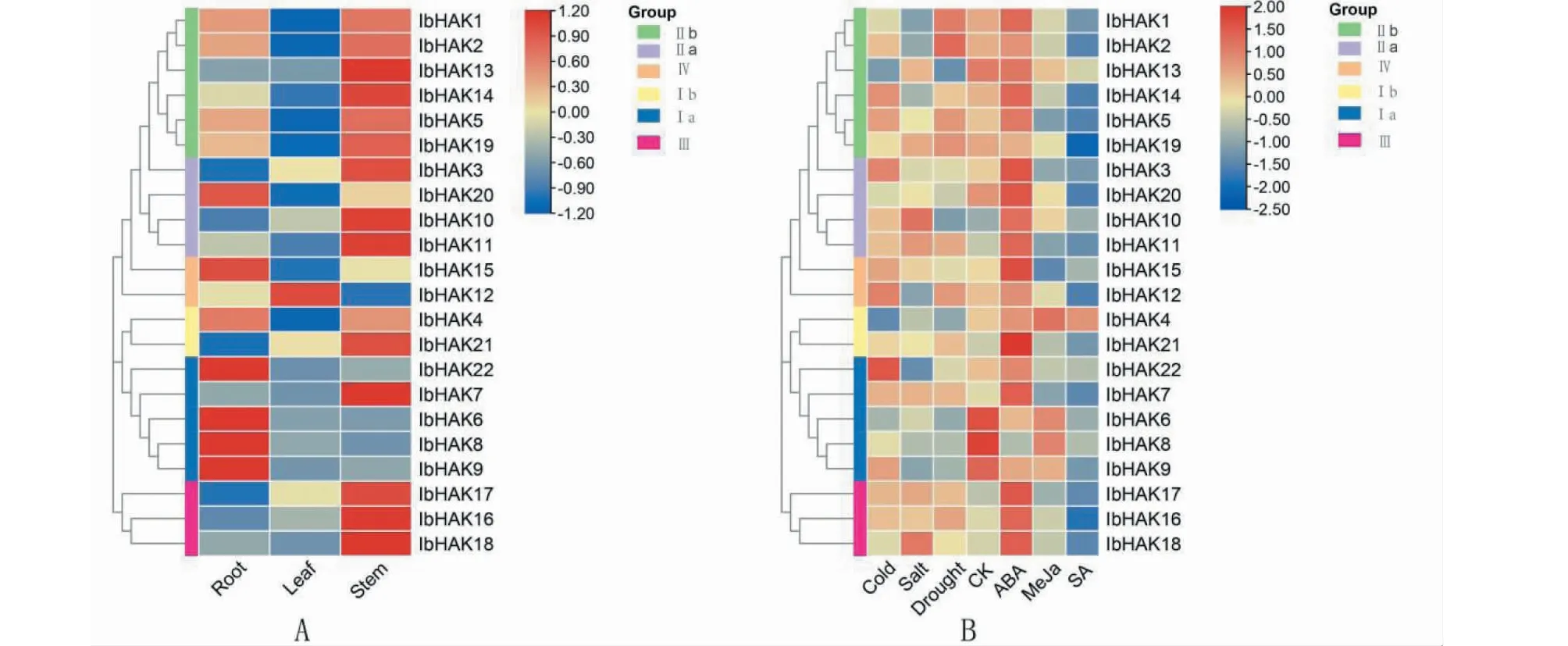
图7 甘薯KUP/HAK/KT基因在不同组织(A)和逆境胁迫(B)中的表达Fig.7 Expression profiles of sweetpotato KUP/HAK/KT genes in different tissues (A) and under adversity(B)
同时利用‘徐薯18’在干旱、盐、冷、水杨酸、脱落酸和茉莉酸甲酯胁迫下的转录组数据,解析IbHAK基因在逆境胁迫下的生物学功能。如图7-B所示,在干旱胁迫下50%(11/22)的KUP/HAK/KT基因表达量上调,IbHAK1和IbHAK2表达量明显上调;在盐胁迫下36.4%(8/22)的IbHAK基因表达量上调,其中IbHAK10和IbHAK18表达量明显上调;在冷胁迫下59.1%(13/22)的IbHAK基因表达量上调,其中IbHAK22表达量显著上调;在脱落酸处理下81.8%(18/22)的IbHAK基因表达量均上调;在水杨酸和茉莉酸甲酯处理下除IbHAK4表达量略有上调外,其余IbHAK基因均呈现表达量下调。
3 讨 论
甘薯是世界上重要的粮食和经济作物,中国是世界上最大的甘薯生产国[16]。而钾作为植物生长所需的三大营养元素之一,在植物生长发育过程中发挥重要的作用,因此研究甘薯K+吸收机制和培育钾高效品种对甘薯的生产具有重要意义[24-25]。本试验利用甘薯基因组信息和生物信息学手段,对甘薯KUP/HAK/KT基因家族进行分析,在甘薯全基因组内共发现22个甘薯IbHAK基因,数量相比水稻(27)、大豆(30)和玉米(27)相差不多。所有的IbHAK基因定位结果都显示位于质膜,与GO注释显示的所有IbHAK基因都具有跨膜转运功能相对应,这与前人对小麦和大豆研究结果一致[26]。此外对家族成员的系统进化关系分析来看,甘薯KUP/HAK/KT基因家族可分为4个进化簇(ClusterⅠ~Cluster Ⅳ),各进化簇的成员分布差异较大,其中77.3%的IbHAKs来源于ClusterⅢ和ClusterⅣ,只有22.7%的IbHAKs来源于ClusterⅠ和ClusterⅡ。在22个甘薯IbHAK基因中共发现3对片段重复的同源物(IbHAK1,IbHAK2;IbHAK2,IbHAK13;IbHAK10,IbHAK11),说明基因复制是导致甘薯HAK/KUP/KT基因家族扩张的主要因素。
KUP/HAK/KT钾离子转运蛋白家族开始于细菌中被发现[27],后续研究表明此家族仅在植物、真菌和细菌中存在,在原核生物和动物中未曾发现[28]。IbHAKs基因表达具有组织特异性,其中IbHAK6,IbHAK8,IbHAK9和IbHAK22在根中超优势表达,其他组织表达量很低,这与其大豆GmHAK30[14]和拟南芥AtHAK5基因[29]的表达模式一致。此外前人研究表明KUP/HAK/KT基因广泛参与植物多种逆境胁迫的响应[30]。其中,拟南芥AtHAK5、AtKUP4和AtKUP11等多个基因在植物受到盐胁迫中高度表达[31];水稻OsHAK5基因在烟草中的过表达,可以通过提高K+在地下部分到地上部分的运输能力,进一步提高耐盐性[32];芦苇PhaHAK2基因在耐盐材料和对照中的转录组数据存在明显差异[33]。在生长激素的处理下,水稻OsHAK5的K+/H+协同转运活性得到了显著提升[34]。本研究通过对‘徐薯18’干旱、盐、冷、水杨酸、脱落酸和茉莉酸甲酯胁迫下的表达模式分析表明,IbHAKs基因在甘薯中存在组织表达特异性,同时在逆境胁迫下也有表达量的变化,说明了IbHAKs积极参与甘薯的生育进程以及和在逆境胁迫下的响应。
4 结 论
甘薯全基因组共鉴定到22个IbHAK基因,全部定位于质膜上,系统进化将其分为4个进化簇,22个IbHAK基因定位在11条染色体上。各成员均含有调控植物生长发育、光响应、逆境响应和激素响应等相关元件,并在不同组织和逆境胁迫下表现出特异性。这为进一步分析甘薯KUP/HAK/KT基因功能提供了生物学依据。
