鸢尾糖基转移酶ItUGT797基因的克隆与表达分析*
2022-09-29黄梓璐陶无恙陈嘉杰段礼新季爱加
姚 薇,李 静,黄梓璐,陶无恙,陈嘉杰,段礼新,季爱加
(广州中医药大学中药学院国际中医药转化医学研究所 广州 510000)
鸢尾(Iris tectorumMaxim.)别名紫蝴蝶、蛤蟆七,性寒味辛、苦,是鸢尾属植物,遍布于各个国家,植株可入药也可观赏[1]。在中国,鸢尾较多分布于陕西、甘肃、青海等西北地区以及四川、贵州、云南等西南地区[2]。鸢尾的干燥根茎可加工为中药川射干,可活血祛瘀以及治疗咽喉肿痛等,因此被称为“治疗喉痹咽痛之要药”。川射干主要药效成分为异黄酮及其糖苷类化合物[3],此类化合物在解除疼痛、治疗炎症、过敏以及癌症等方面效果良好[4-6],其生物合成更是备受关注。
糖基化反应是糖苷类化合物生成途径上涉及到的重要修饰步骤,该反应依赖于糖基转移酶[7]。其过程主要涉及的供体分子是活化后的糖,它在转移酶的催化作用下转移到特定的受体上,从而形成不同的糖苷类化合物,同时苷元的稳定性、亲水性以及生物活性都能得到改善[8-10]。据报道糖基转移酶依据序列相似度和催化特异性可分为99 个家族,其中尿苷二磷酸-糖基转移酶(UGT)家族与植物有效活性成分的合成联系最为密切。UGT 家族作为参与植物发育以及代谢调控的重要糖基转移酶家族,广泛分布于各个物种中,模式作物拟南芥中(120)[11]、水稻(180)[12]以及大豆(182)[13]中的UGT 家族成员已经被鉴定出来。另外,药用植物柴胡、地黄中均有10 个UGT 基因片段被发现[14],有待克隆和功能验证。三七中被鉴别出来的PnUGT82 可以催化黄酮类产物的生成[15],来自红花的CtGT3 可催化7 种化合物进行糖基化[16];黄芩中也有3种新颖的O-糖基转移酶被鉴定出来,既可催化形成黄芩的主要糖苷类成分,也能够催化外源的黄酮底物发生O-糖基化反应[17];金花茶中的CnUFGT14 则有促进多酚合成的作用[18]。底物宽泛性是UGT 家族成员的特性之一,除此外,该家族的成员在C-末端均具有一个PSPG(Plant Secondary Product Glycosyltrans-ferase box)盒子,是包含44 个氨基酸的高度保守区域,而N端则变化多样。UGT 家族成员的结构特征是其行使催化功能、作用底物不一的重要基础[19]。
川射干中的主要活性物质鸢尾苷(tectoridin)是通过异黄酮代谢支路合成。在查尔酮异构酶(chalcone isomerase, CHI)的催化作用下,异甘草素(isoliquiritigenin)合成甘草素(liquiritigenin)。随后在异黄酮合酶(isoflavone synthase,IFS)及2-羟基异黄烷酮脱水酶(2-hydroxyisoflavanone synthase, HID)的催化作用下合成异黄酮骨架,合成鸢尾苷之前还需要在其C-6 位加上甲氧基最后再糖基化。与其他植物例如大豆中异黄酮具有较为显著的雌激素样作用不同,射干类药材异黄酮药理活性更偏向于明显的抗菌消炎等作用[20]。通过两者结构比较发现,大豆异黄酮C-5 位多数没有羟基而射干类药材中的异黄酮则具有C-5 位羟基的结构,从鸢尾中异黄酮结构就可以很明显的发现这一差别,另外鸢尾中的异黄酮多数含有甲氧基。可以推测由于两者结构上的差别造成其在活性上的不同。而糖基化修饰作为关键环节参与了异黄酮结构的形成,可预测该过程对于异黄酮的药理活性也有一定的影响。因而解析川射干中糖基转移酶的修饰功能以及机制对其有效活性成分异黄酮类物质的合成及药理活性具有重大意义。
目前植物中已经被表征功能的UGT 成员很多,涉及到黄酮、萜类以及多种植物激素的生物合成,能够间接或者直接调控植物的生长发育以及品质。而鸢尾作为药用并观赏两用的植物拥有较高的市场价值,其中的UGT 家族成员鲜见报道,因而本研究着重于挖掘鸢尾中的可能参与有效成分鸢尾苷生物合成的UGT 家族成员。利用已知功能的同源基因从鸢尾转录组数据blast 筛选并克隆出一条糖基转移酶基因ItUGT797[21],并对其编码蛋白进行了特征解析,通过同源重组的方法将其克隆到pET-32a 载体上并进行体外原核表达,成功表达出可溶性蛋白,实时荧光定量PCR 实验显示该基因在不同部位均有不同程度表达。这些结果为进一步探索ItUGT797的具体功能及其在鸢尾苷的生物合成中的作用奠定了基础。
1 方法与试剂
1.1 鸢尾总RNA提取以及cDNA的获得
实验材料植物鸢尾(Iris tectorumMaxim.)于2019年6 月采自北京中国科学院植物研究所,后在广州中医药大学栽种。选取生长年限2年,生长健壮,处于盛花期的鸢尾植物,洗净,取须根、根状茎、叶片和花等新鲜组织后立即置于液氮中速冻。参照北京华越洋生物科技有限公司RNA提取试剂盒说明书(Lot:2016#08),提取各组织部位RNA。RNA 提取液各取1μL 进行琼脂糖凝胶电泳检测看是否具备完整条带,其浓度则用岛津BioSpec-nano 核酸定量仪测定。确认合格的mRNA 用于合成川射干的cDNA 链,具体操作按照TakaRa 公 司 的 PrimeScriptTMII 1st Strand cDNA Synthesis Kit反转录试剂盒中的说明进行。
1.2 川射干ItUGT797基因的克隆及表达载体构建
以已知功能的类黄酮7-O-UGT 作为Query 序列,以川射干转录组的pep 文件为Library 进行本地BLASTp,综合基因在根状茎、叶子以及花中的FPKM值与系统发育进化树结果筛选到一条潜在的糖基转移酶基因,命名为ItUGT797。根据其CDS 序列信息,使用Vector NTI设计引物(表1),以鸢尾各部位的混合cDNA 为模板,在冰上按照表2 顺序加入体系,轻弹混匀后按照表3程序进行PCR扩增反应。获得的扩增产物参照生工生物工程(上海)股份有限公司的SanPrep柱式PCR 产物纯化试剂盒说明书进行回收。原核表达载体pET32a 采用双酶切线性化(酶切位点为EcoRI和XhoI)的手段获得具有粘性末端的pET32a 骨架,通过南京诺唯赞生物科技有限公司的同源重组酶将线性化的载体与胶回收产物连接。同源重组的体系按照ClonExpress®ⅡOne Step Cloning Kit 的说明书进行配置。同源基因片段和线性化载体后将其转化至大肠杆菌DH5α,涂布于含100 mg·L-1浓度的氨苄青霉素钠抗性的平板上,置于37℃培养箱过夜培养。挑取大小适中的单克隆进行PCR 鉴定,选取拥有单一且明亮的条带的阳性菌液送测序,测序比对正确后提取质粒,获得pET-32a(+)-ItUGT797重组载体。

表1 引物序列

表2 高保真酶PCR 扩增反应体系

表3 高保真酶PCR 扩增反应程序
1.3 生物信息学分析
将测序得到的ItUGT797序列用各类在线工具解析(表4)。首先利用NCBI 在线工具ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder)搜寻扩增候选糖基转移酶基因ItUGT797序列的开放阅读框;在线工具ProtParam(https://web.expasy.org/protparam/)、CELLO v2.5(http://cello.life.nctu.edu.tw/)和SignalP-5.0(http://www.cbs.dtu.dk/services/SignalP/)被分别用来预测该蛋白的理化性质、亚细胞定位和蛋白信号肽;蛋白跨膜区和蛋白的二、三级结构则分别使用在线工具TMHMM(http://www. cbs. dtu. dk/services/TMHMM/)、SOPMA (https://npsa-prabi. ibcp. fr/cgi-bin/secpred_sopma.pl)和SWISS-MODEL(https://swissmodel.expasy.org/interactive)来预测。DNAMAN 软件被用来进行序列比对,NCBI-cdd 和pfam 网站被用来预测蛋白的结构域范围,最后使用MEGA 7.0 软件中的Neighbor-Joining 法预测基因与其他已被验证功能的糖基转移酶基因的进化关系。

表4 功能表征UGT来源
1.4 川射干ItUGT797 基因Real-time PCR 分析
采用QuantStudio5 实时荧光定量PCR 仪对ItUGT797在各部位的表达情况进行检测,以eEF 基因为内参[21]。鸢尾各组织部位的mRNA 则参照TakaRa公司的反转录试剂盒(RR047A)中的操作说明合成cDNA 链用作模板。按照以下体积配置10 μL 反应原液:2×SYBR Green Pro Taq HS Premix(艾科瑞公司),5.0 μL;正反向引物各0.2 μL;荧光染料0.2 μL;稀释20 倍的cDNA 模板1 μL;超纯水3.4 μL。反应条件:95℃,30 s;40个循环(95℃,5 s;64℃,34 s)。每个样品三次重复,使用2-△△Ct法检测ItUGT797基因在根状茎、叶片和花等部位中的表达水平。
1.5 原核表达
将重组质粒pET-32a(+)-ItUGT797转入宿主菌BL21(DE3)感受态,PCR 鉴定选取阳性克隆菌,接种于1 mL 带有氨苄抗性的LB 液体培养基中,置于37℃摇床,200 rpm 转速培养,至菌液浑浊,以1:100 的比例接种于500 mL 带有氨苄抗性的LB液体培养基中。转速与温度同前,菌液培养至OD 值达到0.5-0.6 的程度即可。18℃,静置30 min,分别加入终浓度0.1 mM、0.3 mM、0.5 mM 的IPTG 进行三个平行实验以摸索最佳的诱导剂浓度。在18℃,180 rpm 条件下诱导培养18 h。后续则在4℃下收集菌体,弃上清。重悬菌体时加入50 mM Tris-HCl,蛋白破碎前加入1 mM PMSF。超声破碎蛋白条件为:间隙时间5 s,超声时间2 s,全程时间18 min,超声功率30%,以上重悬至超声破碎过程均在冰水浴中进行。4℃,12000 rpm,离心30 min,分离上清及沉淀。最终的蛋白表达结果使用SDS-PAGE检测。
2 结果分析
2.1 鸢尾各组织部位RNA的获取
经过1%的琼脂糖凝胶电泳检测,从鸢尾当中提取的各部位的28S 和18S 条带均完整无弥散(图1),凝胶条带亮度越高表明提取RNA 的浓度越高,其中花亮度最高,表明其浓度最大,根状茎的浓度次之,叶片浓度最低,提取的各部位RNA 质量均较好,均可用于接下来反转录实验。

图1 RNA提取凝胶电泳图
2.2 鸢尾ItUGT797基因的克隆以及表达载体构建
以转录组中ItUGT797的CDS 序列为基准,设计带有同源臂的特异性引物,扩增模板为鸢尾的各部位的混合cDNA。电泳结果显示PCR 扩增出的条带处于1500 bp左右(图2),符合ItUGT797基因实际大小。参照生工上柱胶回收之后,通过同源重组的方法将带有同源臂的ItUGT797基因连接到线性化的pET-32a(+)表达载体上,转化后挑取单克隆进行验证,后将阳性菌液测序,测序结果显示与转录组序列一致。而经在线工具预测ItUGT797开放阅读框长度为1455 bp,编码484个氨基酸。

图2 鸢尾ItUGT797基因的克隆
2.3 ItUGT797的生物信息学以及序列分析分析
2.3.1 ItUGT797的理化性质、跨膜区及信号肽预测
ItUGT797 的分子量大小(Molecular,MW)、理论等电点(Theoretical pI)和不稳定系数(Instability index,Ⅱ)等均是通过ProtParam 在线网站预测,结果分别为53.80 kDa,4.94以及52.52。不稳定系数评价结果显示ItUGT797 属于稳定蛋白。ItUGT797 经预测可能定位于细胞质。从在线工具TMHMM 预测的结果来看,ItUGT797 不含有跨膜螺旋区,属于膜外蛋白(图3)。SignalP-5.0 网站显示ItUGT797 没有信号肽,属于非分泌蛋白(图4)。推测ItUGT797 可能在细胞质中发挥功能。

图3 鸢尾ItUGT797的跨膜区预测
图4 鸢尾ItUGT797的信号肽预测
2.3.2 ItUGT797蛋白的二级与三级结构预测
在线工具SOPMA 预测ItUGT797 的二级结构如下:无规则卷曲组成占比最大,为44.42%,较为均匀地分布于各处;其次是占比35.95%的α 螺旋,延伸链占比为13.43%,转角占比最少为6.20%。结果表明ItUGT797 主要由无规则卷曲和α 螺旋组成(图5)。SWISS-MODEL预测ItUGT797基因编码蛋白的三维结构表明,该蛋白与氢醌葡萄糖基转移酶具有26.92%的序列相似性,同源建模以该酶蛋白(2vg8.1)A 链为模板,建模范围为9-470 个氨基酸,GMQE 值(全球性模型估测值)为0.59,三维建模质量良好(图6)。

图5 鸢尾ItUGT797基因的蛋白二级结构预测
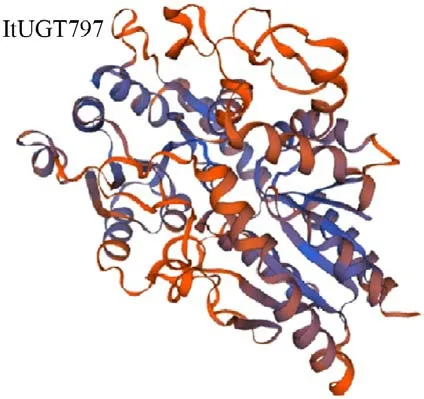
图6 鸢尾ItUGT797基因的蛋白三级结构预测
2.4 ItUGT797的系统发育与结构域分析
为了预测筛选出来的ItUGT797的功能,将其与已被表征过功能的拟南芥、葡萄、大豆、蒺藜苜蓿、葛根等19 种植物UGTs 进行建树。从pfam 网站预测的结果来看,这些UGTs 包括ItUGT797均具有UDPGT 这一关键结构域。根据文献报道,黄酮类化合物的UGTs具有明显的底物偏好性,包括糖受体(黄酮醇、花色素、黄烷酮、黄酮、异黄酮和黄烷醇等苷元)和糖供体(UDP-葡萄糖、UDP-半乳糖、UDP-鼠李糖、UDP-阿拉伯糖和UDP-葡萄糖醛酸)的多样性和特异性[8,22]。根据催化位点的不同,主要可分为O-糖基转移酶(O-UGT)和C-糖基转移酶(C-UGT)两大类。黄酮类化合物的O-糖基转移酶可进一步分为3-OUGT、5-O-UGT、7-O-UGT 和4-O-UGT。系统发育进化树结果表明ItUGT797聚类在Cluster 1,与UGT89C1、Nt7GlcT 等具有7-O-糖基化的活性的UGTs 聚在一支上(图7)。Cluster 2 聚类的则是有异黄酮5-O 位的糖基化活性的UGTs,Cluster 3 均是3-O-UGTs。从建树结果可推测ItUGT797 可能是一种类黄酮7-O-UGT 糖基转移酶。
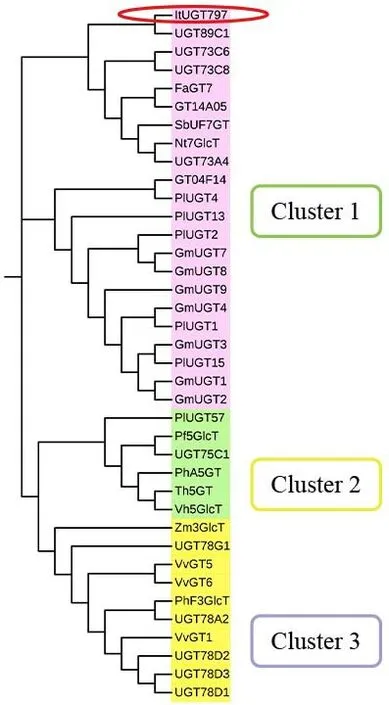
图7 鸢尾ItUGT797与其他物种来源UGTs的系统进化分析
2.5 ItUGT797基因的原核表达
将构建好的表达载体pET-32a(+)-ItUGT797转化至宿主菌BL21(DE3)中,使用菌液PCR 验证是否转化成功。结果表明,目的条带大小与预期一致(图8),取阳性菌液进行IPTG诱导的原核表达。

图8 重组子质粒转入宿主菌BL21(DE3)菌液PCR电泳图
经过改造的pET-32a(+)载体携带20.4 kDa大小的融合标签,旨在促进蛋白的可溶性表达。IPTG 能够诱导重组质粒在大肠杆菌体系中的异源表达,最终目的蛋白以融合蛋白的形式经由SDS-PAGE 表现。相较于空载对照组,pET-32a(+)-ItUGT797 蛋白大小理论值为75.82 kDa,实际条带符合理论值(图9)。改变其IPTG 蛋白诱导剂终浓度这一变量,摸索目的蛋白表达的较优条件。在诱导温度为16℃条件下,分别加入终浓度为0.1 mM、0.3 mM、0.5 mM 的IPTG 诱导剂诱导表达12 h,最终发现诱导剂IPTG 终浓度为0.1 mM 时,上清液可溶性目的蛋白含量最高,表达情况较优(图9)。

图9 改变诱导剂浓度后鸢尾pET-32a-ItUGT797蛋白原核表达SDS-PAGE胶图
2.6 ItUGT797基因在不同组织的转录水平分析
实时荧光定量检测ItUGT797基因在鸢尾根状茎、叶片和花中的表达结果以根茎为准进行归一化(图10)。花中ItUGT797表达最高,其次是叶片,根状茎中水平最低,基因在植物不同部位的表达情况与其功能存在一定的联系[23-25],本研究中ItUGT797基因在花中的高表达可能暗示其更多地在鸢尾花中发挥功能。
3 讨论
类黄酮是广泛存在于蔬菜水果以及观赏性植物中的多酚类化合物,参与植物生长发育以及抗逆过程。类黄酮包含了黄酮、花色苷、黄烷醇、黄酮醇、黄烷酮和异黄酮等6 大类物质。据已有文献报道,在黄酮类化合物中氧苷是最常见的糖基化类型,其中类黄酮主链(苷元)通过黄酮中存在的多种羟基(-OH)基团与糖基(糖供体)相连[26]。通常,黄酮、异黄酮和二氢黄酮多在7-OH 上糖基化形成糖苷键;二氢黄酮醇和黄酮醇则多在3-,7-,3’-,4’-OH上形成单糖苷键,或者会在3,7-二OH,3,4’-二OH 及7,4’-二OH 形成双糖苷。目前在豆科植物中对异黄酮类化合物的糖基化已经研究的比较广泛,主要集中在大豆、葛根和甘草中,大豆中GmUGT1-GmUGT7和GmUGT9均已被鉴定具有异黄酮7-O 位的糖基化活性[27]。葛根中PlUGT1、PlUGT2、PlUGT4、PlUGT13 和PlUGT15 通过克隆和功能表征,发现均可以大豆苷元及染料木素为底物进行7-O 位的糖基化。在黄蓍草中表征的PaUGT1 和PaUGT2 对多种黄酮类化合物具有7-O-糖基化的活性[28]。在植物中,UGTs 通常定位于细胞质中,具有多种生物学功能,参与植物天然产物的生物合成,如类黄酮、苯丙素、萜类和类固醇,以及植物激素的调节[29]。具备稳定天然产物(游离型苷元)、增强水溶性、改变反应性和生物活性等作用。另外还参与了生物活性天然产物的合成、贮存、转运,植物激素和细胞稳态调节以及外源性药物的减毒的过程,具有广泛且重要的价值和意义[30-32]。利用糖基化可以改变和优化药物化合物的潜在用途,丰富植物次生代谢产物的多样性等。
鸢尾中含有丰富的异黄酮化合物及其糖苷,暗示其中存在丰富的(异)黄酮糖基转移酶系统。鸢尾中的糖基转移酶UGT73CD1 被报道能够在鸢尾苷元的7-OH 位置进行糖基化产生鸢尾苷,此外该酶还能催化产生氧苷和氮苷[21]。除此之外鸢尾科植物中少有UGT 基因被研究功能。因此,本研究着重于解析挖掘到的糖基转移酶基因ItUGT797的全貌,包括蛋白结构在内的各项生物信息学指标以及基因的进化关系均被完善分析。QPCR 结果显示从鸢尾转录组中筛选出的糖基转移酶基因ItUGT797在花中表达最高,并且可能是7-O-UGT。鸢尾的花呈现青紫色,富含花青素这一类黄酮化合物,推测ItUGT797可能参与花中包括花青素在内等类黄酮化合物的生物合成中的糖基化修饰。具体的功能验证需要后期进行体外酶活实验确定。且ItUGT797在根状茎和叶中也有表达,鸢尾苷和鸢尾苷元主要分布于根状茎部位,这暗示着它也有可能参与根状茎中鸢尾苷的合成。已有研究表明,参与有效活性成分生物合成的基因的表达模式与次生代谢物的积累模式类似[23,33-34]。例如,常见药用植物丹参的药理活性成分丹参酮以及丹酚酸主要在根部积累,而许多调控这两种化学成分生物合成的转录因子也在根中高表达[24-25,35]。在除此之外,对于植物体而言,异黄酮类化合物参与植物生长发育和抵抗胁迫的过程,通过提高植物对环境紫外的抵抗和对病菌和昆虫的抵御能力来增强自身的保护[36-37]。因此,ItUGT797作为可能参与异黄酮类化合物糖基化修饰的成员,在植物生物发育、次生代谢以及防御抗逆过程中的功能值得进一步发掘研究。

图10 鸢尾ItUGT797基因在不同部位的表达情况
4 总结以及展望
鸢尾作为著名的药用和园艺植物,市场需求极大。其主要活性成分为鸢尾苷、鸢尾新苷B、野鸢尾苷等多种异黄酮类化合物[3]。异黄酮糖苷类化合物的生物合成依赖于糖基转移酶(UGTs)的修饰。本研究从鸢尾转录组中鉴定出ItUGT797这一糖基转移酶基因,网站预测结果表明其为稳定的膜外非分泌蛋白,且主要由无规则卷曲组成;ItUGT797在叶片中表达最高。使用大肠杆菌为媒介的原核表达结果表明该基因可以表达出较少的可溶性蛋白。系统发育进化树结果暗示ItUGT797 可能是一类主要以类黄酮为底物进行催化过程的糖基转移酶。已有很多物种的糖基转移酶被验证可以提高植物有效活性成分含量和参与抵御生物胁迫[38-39]。然而,关于鸢尾中糖基转移酶的研究较少,本研究后续将结合体外酶活实验验证ItUGT797的功能。挖掘鸢尾中的UGTs 既为其可能参与植物生长发育、抗逆等解析了机制也为实现天然药物的生物合成提供更多工具酶,最终达到提高药用植物品质的目的。
