外源草酸对不同年限狗尾草种子萌发及生理生化的影响
2022-09-29旷思萍蒋金娟罗富成祖艳群
徐 翠, 姜 娜, 旷思萍, 蒋金娟, 罗富成*, 祖艳群*
(1. 云南农业大学动物科学技术学院, 云南 昆明 650201; 2. 云南农业大学资源与环境学院, 云南 昆明 650201)
‘纳罗克’非洲狗尾草(Setariasphacelata‘Narok’)是禾本科(Gramineae)狗尾草属(Setaria)多年生草本植物,其牧草品质优良,干草产量可达16 500 kg·hm-2,不仅是亚热带、暖温带地区草业生产主要的牧草之一[1],还是水土保持、边坡植被恢复的地被植物[2]。‘纳罗克’种子休眠程度较深,种子休眠是植物在长期进化过程中抵抗不良环境所形成的一种生态适应性,从种族繁衍、种质保持方面来看,种子休眠是一种非常有利的特性[3],但作为播种材料,该种子要自然储藏1~2年后才具备较高的发芽能力[4],而种子活力水平往往与储藏时间呈反比关系,意味着随时间延长具有高活力水平的种子逐渐减少,这会给种子生产、人工草地建设带来极大的挑战[5],针对狗尾草种子的休眠学者展开了一系列研究,如用物理方法去除稃片增强种子的呼吸作用,或用化学方法强酸强碱腐蚀种皮突破萌发障碍[6]。但在实际生产过程中,除了存在复合休眠外[7],该牧草种子体积微小,导致休眠解除技术难以推广且不易操作。
草酸(Oxalic acid)是生物体的一种代谢产物,广泛分布于植物、动物和真菌体中,并在不同的生命体中发挥不同的功能[8]。研究表明,外源草酸在毫摩尔浓度水平具有极强的抗氧化性能[9],通过抑制蔬果的木质素代谢和提高抗氧化酶系统,果实细胞膜透性增加趋势被缓解,丙二醛含量降低,最终减缓果实成熟进程或延缓冷藏期内品质的下降[10],因此在适当用量范围内可以作为一种有效的抗氧化剂。而抗氧化剂和抗氧化酶的联合作用可以有效清除种子休眠过程中ROS对细胞产生的损伤[11]。草酸处理诱导提高果实抗氧化酶活性,降低ROS水平和膜脂过氧化程度,有助于维持细胞膜结构的稳定性[12]。庄正等[13]发现低浓度外源草酸具有促进杉木(Cunninghamialanceolata)种子萌发及幼苗生长的作用,且对杉木幼苗无明显伤害。而种子萌发过程也都伴随着活性氧(ROS)的产生和积累[14]。ROS的过量积累会导致膜脂过氧化不断加强,造成细胞膜结构破坏、电解质外渗和细胞膜受损,从而抑制种子的萌发[15-16]。丙二醛(MDA)是细胞膜质过氧化的产物,植物细胞中大量丙二醛加重细胞膜脂质过氧化,其含量越高,种子活力越低[17]。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)作为抗氧化酶系统中重要物质,能通过清除体内多余的ROS保持其平衡。抗氧化酶的活性也与种子的萌发表现出显著正相关性[18],随着抗氧化酶活性的提高,种子活力也会随之增加[19]。糖分子作为一种信号分子参与许多植物激素如乙烯、细胞分裂素、生长素等的信号转导途径,调控种子萌发和幼苗生长的生理过程,在拟南芥(ThaleCress)的种子和幼苗中,可以在有外源葡萄糖的情况下促进种子的萌发。目前外源草酸的添加主要应用于蔬果的保鲜,缓解植物根系重金属毒害等方面[20],关于休眠种子的报道较少。因此本试验预期将外源草酸作为一种抗氧化剂,研究外源草酸处理对‘纳罗克’非洲狗尾草种子萌发的影响,进一步探讨不同储藏年限种子经草酸处理后种子萌发及生理生化的变化,以期为外源物质对休眠种子的深入研究奠定理论基础,为揭示草酸提高种子萌发活力的机理提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为自然储藏0,1,2,3,4年的‘纳罗克’非洲狗尾草种子(Setariasphacelata‘Narok’),收获时间为每年的9—10月,试验室种子含水率分别为8.5%,7.4%,7.8%,7.9%,7.2%。采自于云南农业大学草业科学实践教学基地,原始材料经种子重力分离机(荷兰,Selecta Machinefabriek BV)清选,风力值为34 m·s-1,充分混匀后用作试验材料。试验于2020年1—6月完成,采用TTC(四氮唑)图像法[21]测定种子生活力分别为84.4%,81.4%,64.0%,53.2%和10.6%。
1.2 试验设计
储藏0,1,2,3,4年的‘纳罗克’非洲狗尾草种子用1.5%的硫酸铜消毒15 min,蒸馏水浸泡30 min,换水重复清洗3次[22],用吸水纸吸干种子表面水分备用。选择收获当年的种子用于筛选草酸处理参数,以不同草酸浓度设置5个梯度(0,5,10,15,20,25 mmol·L-1),不同处理时间设置4个梯度(0,5,15,25 min),根据不同浓度和不同处理时间设置组合,每个处理3 g种子,加入50 mL相应浓度的草酸进行试验,以浸入50 mL蒸馏水处理25 min的种子为对照。处理后的种子用蒸馏水清洗3次,并置于室内回干12 h,回干时室内温度为22.6℃,结合发芽试验确定最佳处理参数。
根据发芽指标选择最佳处理参数,用于处理储藏0—4年的‘纳罗克’非洲狗尾草种子,测定处理后种子的发芽率、发芽势、抗氧化酶活性、丙二醛含量和可溶性糖含量。待21 d种子发芽试验结束后,测定种子发芽指数、活力指数和幼苗的生长情况。
1.3 指标测定与方法
1.3.1发芽试验 发芽试验采用纸上发芽法[23]。对于处理组和对照组的种子,各取100粒种子,重复3次,并将种子置于内有湿润滤纸的培养皿中,于24 h光照的人工气候箱中(25±1)℃进行发芽试验,光照强度为180 Lx,逐日统计发芽数。分别在第7 d,第21 d统计种子的发芽势和发芽率。待21 d发芽试验结束后,将幼苗置于105℃的烘箱中杀青15 min,然后于80℃的烘箱中干燥24 h后,重复称其干重至恒重,并按照下列公式计算种子的发芽指数和活力指数[6]。
发芽指数(GI)=∑(Gt/Dt)
活力指数(VI)=GI×S
式中:Dt为发芽时间(d);Gt是与Dt相对应的每天发芽种子数;S表示发芽试验结束时正常幼苗的单株干重(g)。
1.3.2幼苗生长测定 21天发芽试验结束后,随机选取10株幼苗,游标卡尺测量根长和苗高,保留两位小数。
1.3.3种子生理指标测定 SOD活性采用NBT显色法[24];POD活性采用愈创木酚氧化法[24];CAT活性采用H2O2法[25];MDA含量采用TBA测定法[26];可溶性糖含量采用蒽酮-硫酸试剂法[26]。
1.4 数据分析
所得数据使用Microsoft Excel 2010整理分析、Origin 2018作图,采用SPSS 23.0进行单因素方差分析、线性回归分析、相关性分析和主成分分析,Duncan方法检验各处理水平在0.05水平的差异性。
2 结果与分析
2.1 不同草酸浓度与处理时间对当年种子萌发的影响
草酸处理后,狗尾草种子的萌发指标与对照相比均有显著提高(P<0.05)。随着草酸时间的延长,狗尾草种子的发芽率逐渐升高,15和25 mmol·L-1的草酸处理25 min的发芽率较高,其发芽率分别为74.0%和84.0%;种子的发芽势均在31.0%~65.0%之间,其中较高的两个处理组合参数与发芽率相同,即15和25 mmol·L-1的草酸处理25 min,发芽势分别为63.3%和64.3%;发芽指数与活力指数的变化相似,25 mmol·L-1的草酸处理25 min后种子的发芽指数,活力指数差异显著,两者分别为16.1和16.3,是对照的13.3倍和12.5倍(表1)。
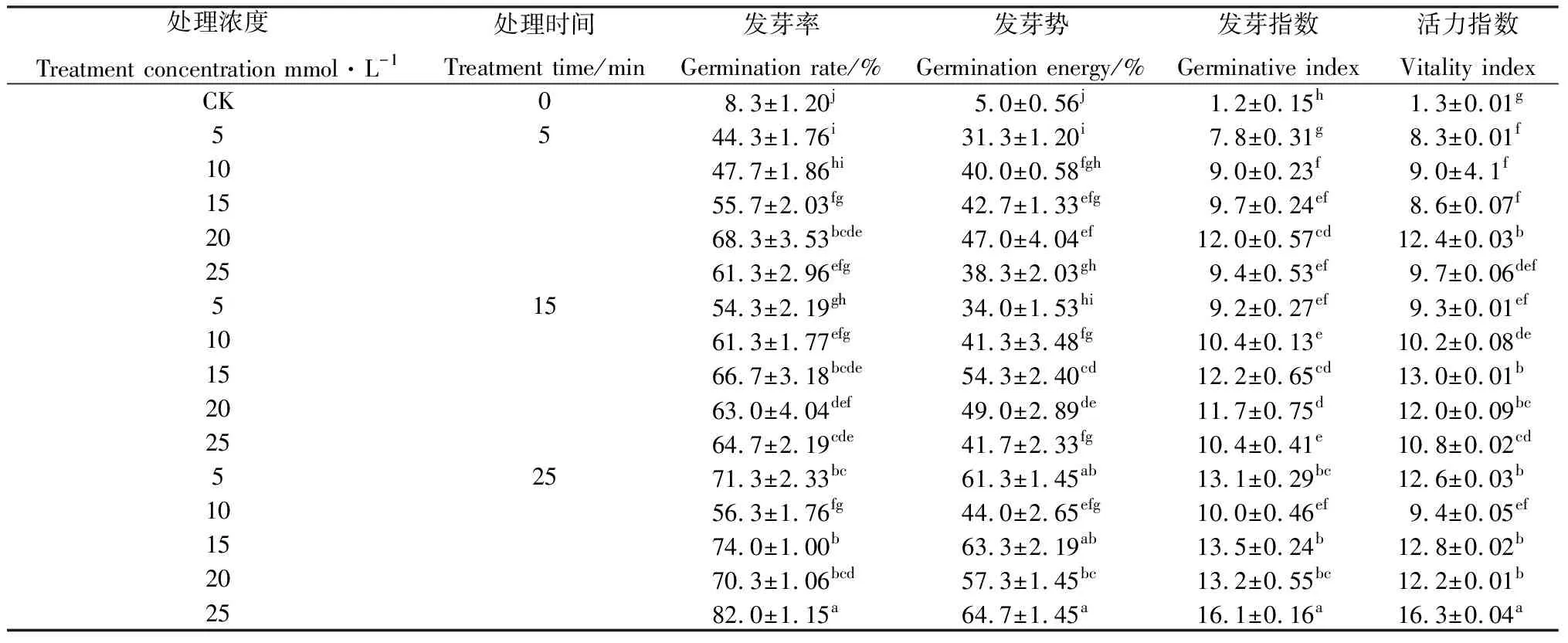
表1 不同的草酸处理组合对收获当年‘纳罗克’种子萌发指标的影响Table 1 Effects of different oxalic acid treatments on germination indicators of ‘Narok’ seeds in the year of harvest
对狗尾草种子萌发特性与草酸浓度与处理时间的关系进行了多元回归分析,处理时间是种子发芽特性解释率最大的因子。处理时间对种子的发芽率、发芽势、发芽指数和活力指数存在极显著正相关,而草酸浓度除与发芽指数呈显著正相关外,与发芽率、发芽势、活力指数也呈极显著正相关(表2)。

表2 狗尾草种子萌发和草酸浓度与处理时间的多元线性回归分析Table 2 Multi-linear regression analysis of seed germination and oxalic acid concentration and treatment time
2.2 不同储藏年份种子萌发对草酸处理的响应
草酸处理后,收获当年、储藏1,3,2,4年的种子,发芽率,发芽势,发芽指数和活力指数逐级递减,储藏0年的发芽指标均与其他储藏年份存在显著差异(P<0.05)。与CK相比,收获当年种子的发芽指标均得到显著提高,其发芽指标分别比CK提高了8.9,13.0,9.8和7.3倍。储藏1年和3年的种子除发芽势外,其他萌发指标无显著差异,但储藏1年时种子发芽势,发芽指数比CK提高3.4倍和1.1倍(表3)。
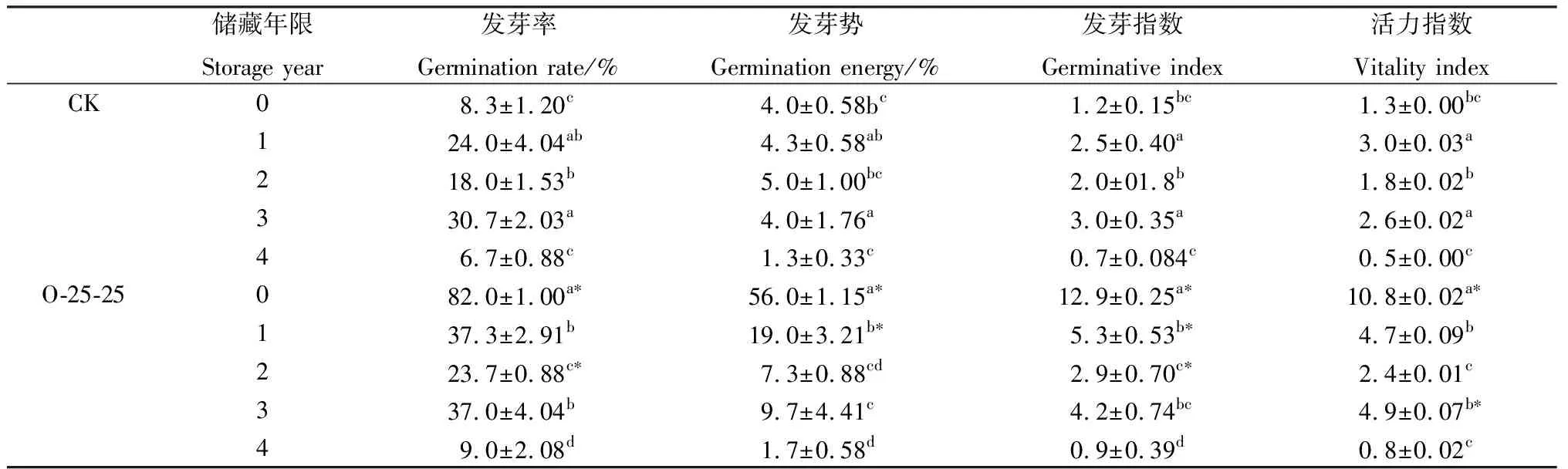
表3 草酸引发对不同储藏年份种子发芽指标的影响Table 3 Effect of oxalic acid initiation on germination indicators of seeds of different storage years
2.3 不同储藏年份种子幼苗生长对草酸处理的响应
随着储藏年份的延长,草酸处理后的幼苗根长呈单峰趋势变化,储藏0~2年种子的根长较长,分别较CK提高了37.3%,20.0%和226.0%,其中储藏2年时的种子幼苗提高效果最好,与其他储藏年份存在显著差异(P<0.05)。草酸处理后均能显著提高幼苗的苗高,较CK相比提高了7~19倍,处理后各苗高之间无显著差异,说明草酸处理能促进幼苗生长的一致性(图1)。
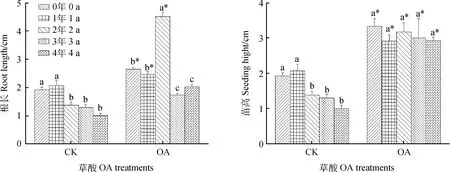
图1 草酸处理对幼苗根长及苗高的影响Fig.1 Effect of oxalic acid treatment on root length and seedling height注:“OA Treament”表示草酸处理Note:“OA Treament” indicates oxalic acid treatment
2.4 不同储藏年份种子丙二醛含量及抗氧化酶活性对草酸处理的响应
草酸处理后,不同储藏年限的种子MDA含量随储藏年份的延长呈逐渐上升趋势,收获当年的种子MDA含量显著低于其他储藏年份,与CK相比,储藏0~2年的MDA含量均有所下降,其中收获当年的与CK差异最大,较CK降低了35.4%。SOD活性随储藏年限的延长呈先上升后下降的趋势,储藏1年时的SOD活性可达到最大值2614.3 U·g-1,与其他储藏年限存在显著差异;与CK相比,草酸处理提高了不同储藏年限种子的SOD活性,其中变化最大的是收获当年的和储藏1年的种子,SOD活性较CK提高了87.3%,45.5%。草酸处理后,收获当年的种子POD活性显著高于其他储藏年份,比CK降低了18.5%,储藏3年和4年种子的POD活性比对照升高了5.9倍和1.4倍。草酸处理后,种子CAT活性随储藏年份的延长呈逐渐下降趋势,收获当年的种子CAT活性与其他储藏年份差异显著,并且收获当年的CAT活性与CK差异最大,较CK提高了1.7倍(图2)。
2.5 不同储藏年份种子可溶性糖含量对草酸处理的响应
草酸处理后,种子可溶性糖含量随储藏年份逐渐降低,收获当年和储藏1年的种子与其他储藏年限存在显著差异,最高可达6.5%,储藏1~3年时种子可溶性糖含量在3.4%~3.7%之间,各储藏年份之间无显著差异。与CK相比,草酸处理显著提高了收获当年和储藏1年种子的可溶性糖含量,分别比CK提高了154.1%,91.4%(图3)。
2.6 种子发芽指标与生理特征相关性分析及主成分分析
草酸处理后种子的POD活性与发芽指标具有极显著正相关性(P<0.01),与幼苗的苗高具有显著正相关性(P<0.05)。CAT活性与发芽率,发芽势具有极显著正相关性。除根长外,MDA含量与发芽指标、幼苗生长具有负相关性,其中与发芽势、发芽指数呈显著负相关(表4)。
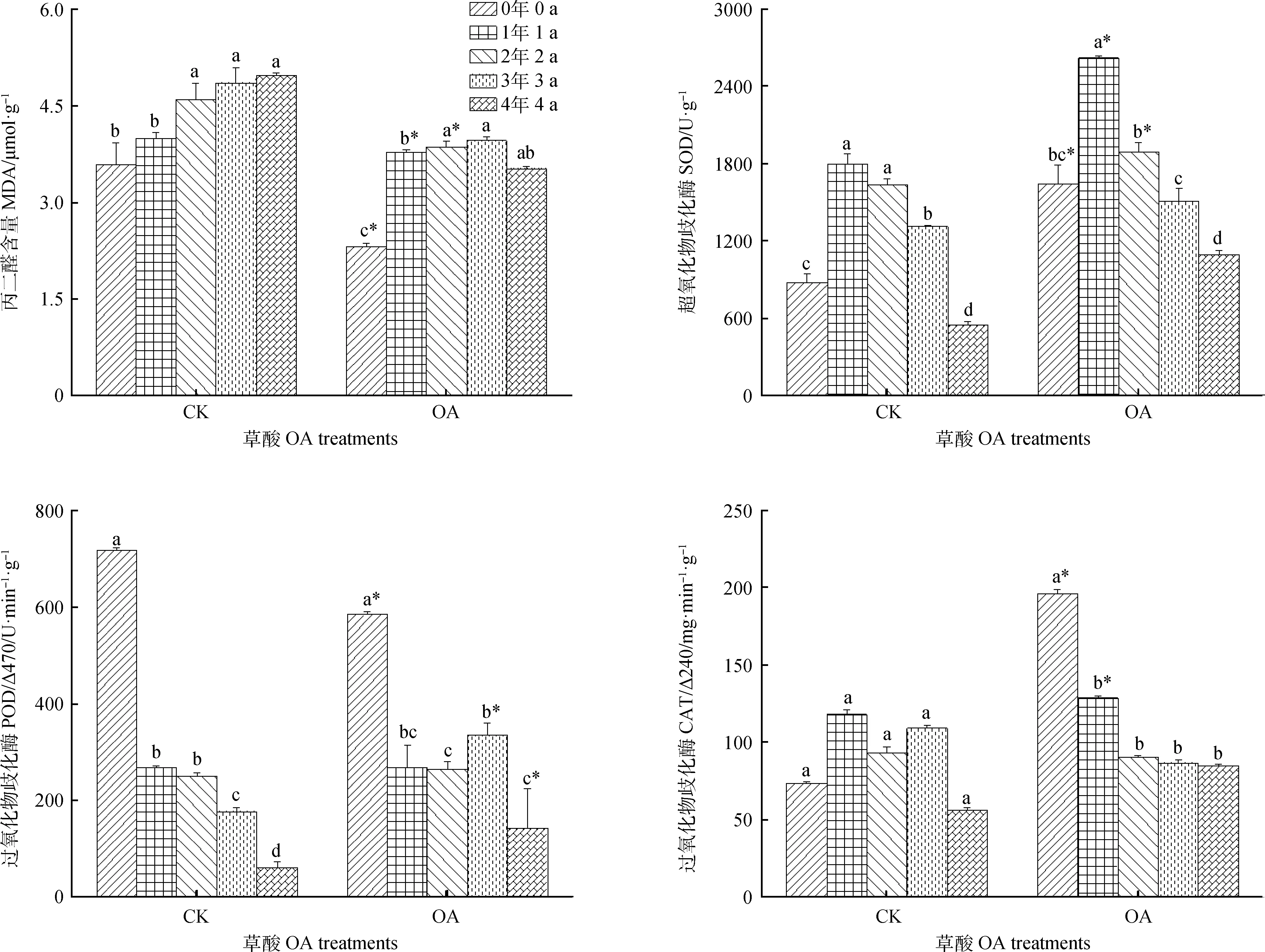
图2 草酸处理对不同储藏年份种子丙二醛含量及抗氧化酶活性的影响Fig.2 Effect of oxalic acid treatment on the MDA,SOD,POD activity and CAT content of S.sphacelata seeds of different storage years

图3 草酸处理对不同储藏年份种子可溶性糖含量的影响Fig.3 Effect of oxalic acid treatment on the soluble sugars content of S.sphacelata seeds of different storage years

表4 种子发芽指标与生理生化指标的相关性Table 4 Correlation of seed germination indicators with physiological and biochemical indicators
为了更好的比较外源草酸对不同储藏年份种子的处理效果,发芽率(X1)、发芽势(X2)、发芽指数(X3)、活力指数(X4)、根长(X5)、苗高(X6)、SOD(X7)、POD(X8)、CAT(X9)、MDA(X10)和可溶性糖(X11)采用主成分分析法做进一步分析。所检测的所有指标经标准化处理后,对种子的发芽指标和生理生化指标变化进行主成分方差贡献率分析(表5),以特征值>1为提取标准,得到3个主成分的累积贡献率分别为70.280%,83.351%,95.882%,说明3个主成分能较好的反映正眼数据的信息特征。

表5 主成分方差分析Table 5 PCA of variance
由成分载荷矩阵(表6)可知,发芽率、发芽势和发芽指数在第1主成分上有较高载荷,说明第1主成分主要反映种子萌发的信息,各指标贡献率大小顺序为发芽势>发芽指数>发芽率。SOD活性和MDA含量在成分2上载荷较高,且贡献率大小顺序为SOD>MDA,说明第2主成分主要反映种子抗氧化酶活性及脂质过氧化程度的信息。第3主成分根长和苗高的载荷量较高,说明主成分3是反映了幼苗的生长情况,其贡献率大小顺序为根长>苗高。
由表7可知,收获当年和储藏1年的种子综合得分较高,分别为3.25,0.32。5个不同年限的种子主成分分析得分高低顺序依次为:0年>1年>2年>3年>4年。

表6 成分载荷矩阵Table 6 Component matrix
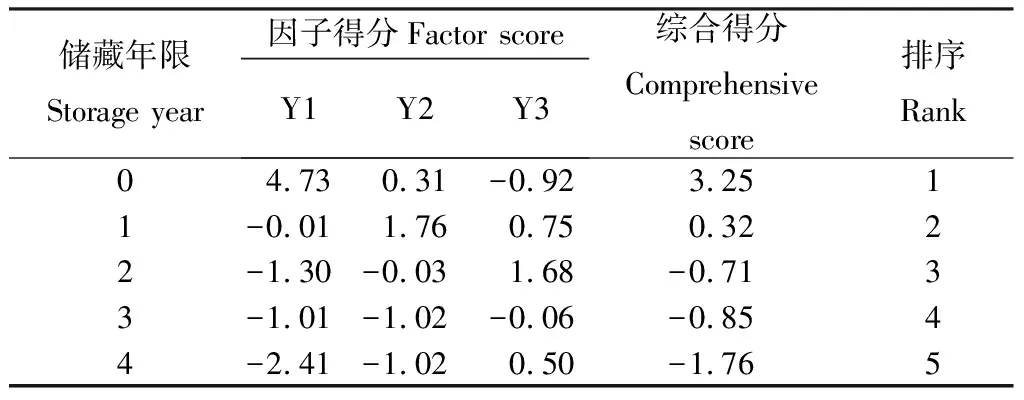
表7 草酸处理后不同储藏年限种子主成分综合得分及其排序Table 7 Scores and ranking of principal component on S.sphacelate seeds of different storage years after oxalic acid treatment
3 讨论
3.1 草酸处理对狗尾草种子萌发的影响
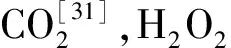
3.2 不同储藏年份狗尾草种子生理特征对草酸处理的响应
种子在萌发的过程中,为满足能量所需,种子内部的能量物质在萌发阶段水解通过乙醛酸循环、丙酮酸代谢等途径不断转化为小分子有机酸类物质[33]。但种子处在吸水阶段时,细胞不断吸水破裂导致有机酸外渗[34],导致能量物质不断降低。不同储藏年限的‘纳罗克’非洲狗尾草种子经外源草酸添加后,萌发指标均有一定程度的升高,收获当年的种子发芽率、发芽势、发芽指数和活力指数分别是对照组的8.8,13.0,10.0和7.3倍。可能是因为种子木质化程度影响外源物质的渗入,对于收获当年和储藏1年的种子,其种壳木质化程度较轻,有利于外源草酸的浸入,弥补种子发芽时有机酸等能量物质的损失。随着储藏时间的延长至3年时,种子老化[35]和种壳木质化程度加剧[36],其致密的种壳致使外源草酸难以进入种子内部,但此时种子的可溶性糖含量也有一定程度的累积,也可为种子萌发时提供能量所需[37]。
随着储藏年限的延长,通过草酸处理降低MDA含量的效果逐渐减弱,MDA含量可以表达细胞膜损伤的程度[38],MDA含量的降低不仅表示植物细胞膜修复的完整性,还能减少植物细胞壁木质化的合成。说明对于储藏年限短的种子来说,草酸处理可以降低木质素代谢、减少木质化的合成,有利于种子萌发过程中与外界进行水分和气体交换。本试验中,储藏年限较短(0~1年)的休眠种子SOD和CAT活性在25 mmol·L-1草酸处理25 min后都有显著提高,收获当年种子的SOD和CAT活性变化最大,分别比对照提高87.3%和164.4%,并且草酸处理后种子的CAT活性与萌发指标呈极显著正相关(P<0.01)。SOD作为抗氧化系统中的第一道屏障,通过歧化反应将活性氧(ROS)转化为H2O2,而H2O2在POD,APX和CAT等抗氧化物质的作用下可转化为H2O 和O2,降低多余H2O2产生的毒害作用[33]。说明外源草酸处理后,种子萌发时产生多余的ROS主要通过SOD和CAT的协同调控对氧化损伤起到防御作用[39],徐慧敏[40]在研究低分子有机酸对辣椒(Capsicumannum)生长发育及叶片活性氧代谢时也出现类似的结果。25 mmol·L-1的草酸处理提高了储藏3~4年种子的POD活性,比对照组提高了89.6%和137.2%,表现出随储藏年限延长而增加的趋势,但显著降低了收获当年种子的POD活性,这与沈玫等[41]的研究结果类似,草酸处理能够显著抑制去壳绿竹笋(Dendrocalamopsisoldhami)的木质素合成关键酶PAL,CAD,4-CL和POD的基因表达水平,而这4种酶的活性与其对应基因的表达是协同调控木质素的合成,能提高组织的木质化程度[42],说明POD活性的升高可能造成木质化程度的加剧,草酸处理通过抑制POD活性基因的表达,在减少POD活性的同时也降低了种子木质化程度。因此,综合分析评价可见,用25 mmol·L-1,25 min的草酸处理储藏0~1年狗尾草种子对促进种子萌发与幼苗生长较为适宜。
4 结论
5~25 mmol·L-1草酸处理能显著提高收获当年‘纳罗克’非洲狗尾草种子的萌发特性,其中收获当年的种子在草酸浓度25 mmol·L-1,时间25 min时,可以达到破除种子休眠的效果。草酸处理能够有效清除储藏年限较短种子的MDA含量,提高种子SOD,CAT活性和可溶性糖含量。但储藏年限超过1年后,外源草酸处理对种子的萌发效果越差。推荐采用25 mmol·L-1,25 min的草酸处理收获当年和储藏1年时的狗尾草种子。
