墨吉明对虾高血糖激素基因FmCHH-I的分子鉴定和表达特征
2022-09-28雷易果陈兆明
雷易果,陈兆明,王 伟
(广东海洋大学水产学院,广东 湛江 524088)
在甲壳动物中,高血糖激素(Crustacean Hyperglycemic Hormone,CHH)是一类由眼柄中的X 器官-窦腺-复合体合成并分泌的神经肽类激素[1]。CHH 家族神经肽可分为I 型和II 型两大类[2],CHH-I型神经肽有二硫键结构,信号肽与成熟肽之间由一段前导肽(CHH precursor-related peptide,CPRP)连接,而CHH-II 型神经肽结构中的信号肽直接与成熟肽相连。CHH-I 型神经肽仅包括经典的CHH 类激素,主要参与调节血淋巴中的葡萄糖水平;CHHII 型神经肽种类多样,主要包括蜕皮抑制激素(Molt-inhibiting hormone,MIH)、大颚器官抑制激素(Mandibular organ-inhibiting hormone,MOIH)以及性腺抑制激素(Gonad-inhibiting hormone,GIH)[3]。CHH 家族神经肽在甲壳动物的脂质代谢、蜕皮、应激反应、性腺发育等多个生理过程中均发挥重要调节作用[4-5]。
我国是对虾养殖大国,凡纳滨对虾(Litopenaeus vannamei)是人工养殖的主要虾种,但其种质资源和亲虾严重依赖于进口,当前的对虾行业还存在养殖环境恶化、种质退化、病害频发等问题[6],这些都制约着对虾养殖产业的可持续发展。因此,开发凡纳滨对虾的替代或补充物种十分必要。墨吉明对虾(Fenneropenaeus merguiensis)是浅海的暖水性虾类,在我国东南沿海均有分布,若其人工养殖技术得以突破,可在一定程度上缓解目前存在的对虾养殖问题[7]。在墨吉明对虾基础生物学方面已有一系列研究,史黎黎等[8]研究了墨吉明对虾C 型溶菌酶基因的克隆及表达,周婷婷等[9]研究了胰岛素样促雄性腺激素基因(FmIAG)的分子特征,杨世平等[10]研究了丁香酚对墨吉明对虾的麻醉效果,为其人工繁殖方法的开发提供了参考。但是,目前墨吉明对虾的大规模人工养殖技术还有待开发和完善,其基础生物学知识还比较匮乏。有关对虾CHH 家族神经肽的种类和功能,研究者已进行了一些探索,但目前对于墨吉明对虾的CHH 家族基因信息了解的还比较有限,眼柄中的X器官对CHH家族基因的调控作用以及CHH 家族神经肽在墨吉明对虾性腺发育中的功能演化仍有待于阐明。本研究利用分子生物学技术和生物信息学分析,获得了墨吉明对虾CHH-I基因(FmCHH-I)cDNA 序列,探究FmCHH-I的组织表达情况以及卵巢发育不同时期FmCHH-I的表达规律,分析生产上常用的单侧眼柄切除法对FmCHH-I表达的影响,为探明FmCHH-I的功能奠定基础,并为甲壳动物CHH神经肽的相关研究提供参考。
1 材料与方法
1.1 材料
实验用墨吉明对虾购自广东省湛江市霞山区水产品批发市场,在室内循环水养殖系统中进行暂养,系统由供气系统、给回水系统、生物滤池等组成,调节室温为26~28 ℃,每天定时定点定量投喂3次饲料,对水体进行连续充气并定期换水,维持良好水质,观测对虾的活动状态是否异常。挑选活力较好的对虾[体质量(30.5±0.8)g,体长(13.5±1.2)cm]进行后续实验。
1.2 实验试剂
TransZol Up Plus RNA Kit(RNA 提取试剂盒)、TransScript One-Step gDNA Removal 反转录试剂盒购自TransGenBiotech 公司,Primer ScriptTM1st Strand cDNA Synthesis Kit 逆转录试剂盒购自日本TaKaRa公司,SYBR PrimeScript RT-PCR Kit购自SangonBiotech公司,SMARTerTMRACE cDNA Amplification Kit 购自美国Clontech 公司,M5 HiPer Ptopo-TAVector、DH5α 大肠杆菌感受态细胞菌种购自全式金生物试剂公司。
1.3 总RNA 提取与FmCHH-I 基因cDNA 序列全长获取
使用RNA 提取试剂盒,用Trizol 方法提取墨吉明对虾总RNA。后通过反转录试剂盒将其反转录为cDNA。
根据本课题组前期获取的墨吉明对虾转录组序列信息(GenBank 序列号ON646230)设计特异性引物(表1),以墨吉明对虾总RNA逆转录产物cDNA为模板,克隆出FmCHH-I基因的中间序列。采用Primer 5.0 软件设计3′RACE、5′RACE 特异性引物(表1),利用RACE 技术扩增FmCHH-I基因的5′-UTR 和3′-UTR,从而得到FmCHH-I的cDNA 全长。PCR 产物通过琼脂糖凝胶电泳检测,对目的片段进行纯化回收。通过连接、转化后,按照实验操作,提取出菌液后,用此菌液作为模板,选用通用引物M13(-47)和M13(-48),进行菌液PCR 扩增,通过琼脂糖凝胶电泳检测反应产物。分装200 μL 菌液,送广州艾基生物技术有限公司进行测序。

表1 PCR引物Table 1 PCR primers
1.4 生物信息学分析
通过NCBI 数据库中BIASTX 在线程序(http://blast.Ncbi.nlm.nih.gov/Blast.cgi) 对墨吉明对虾FmCHH-I基因进行同源性分析,利用SignalP 4.1 Server(www.cbs.dtu.dk/services/SignalP/)预测该氨基酸序列的信号肽以及具体位置;利用在线软件Expasy(http://web.expasy.org/compute_pi/) 预 测FmCHH-I 蛋白分子质量和等电点;使用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)比对墨吉明对虾与其他物种CHH基因的氨基酸序列;用MEGA 7.0邻接(NJ)法构建系统进化树。
1.5 FmCHH-I基因的组织表达
取健康墨吉明对虾的脑、眼柄、鳃、肝胰腺、肠、肌肉、卵巢等7种组织,进行RNA提取以及cDNA转录(方法见1.3)。用半定量RT-PCR方法检测FmCHH-I在墨吉明对虾不同组织中的转录情况。以EF1α 为内参引物,RT-PCR 扩增条件为95 ℃3 min;95 ℃30 s,58 ℃30 s,72 ℃30 s,35个循环;72 ℃10 min。
1.6 不同性腺发育时期FmCHH-I的表达
参考文献[14]的方法,根据卵巢的外观形态和性腺发育指数(GSI)区分墨吉明对虾卵巢的不同发育阶段。选择处于卵巢成熟期与未发育的雌性墨吉明对虾各3 尾,取其眼柄组织,进行RNA 提取并反转录为cDNA。以反转录的cDNA 为模板,设计定量PCR 引物qFm-CHH-IF/ qFm-CHH-IR,进行实时荧光定量PCR 检测,EF1α 为内参引物(表1),目的基因和内参基因各做3个平行扩增。反应条件为95 ℃5 min;95 ℃15 s,55 ℃60 s,75 ℃5 s,40个循环。用2-△△CT法比较FmCHH-I基因在不同发育时期的眼柄组织中的表达量。利用SPSS 软件对数据进行单因素方差分析,用Duncan 方法进行多重比较,显著性水平设为0.05。
1.7 单侧眼柄切除对FmCHH-I基因表达的影响
随机取处于性腺成熟期的雌性墨吉明对虾3尾,用镊烫法切除单侧眼柄。眼柄组织RNA提取和基因表达量检测方法见1.6。比较切除及未切除眼柄的雌性墨吉明对虾眼柄组织的CHH-I表达量。
2 结果与分析
2.1 FmCHH-I基因cDNA序列及其结构
克隆得FmCHH-I基因的cDNA 全长599 bp,其中5′非翻译区(UTR)长度为37 bp,3′-UTR 长度为193 bp,开放阅读框(ORF)长为369 bp,编码123 个氨基酸(图1),其中在第1~20 位包含一个信号肽(图2),随后有一段由26个氨基酸组成的前导肽,末端的成熟肽具有74个氨基酸,无跨膜区。
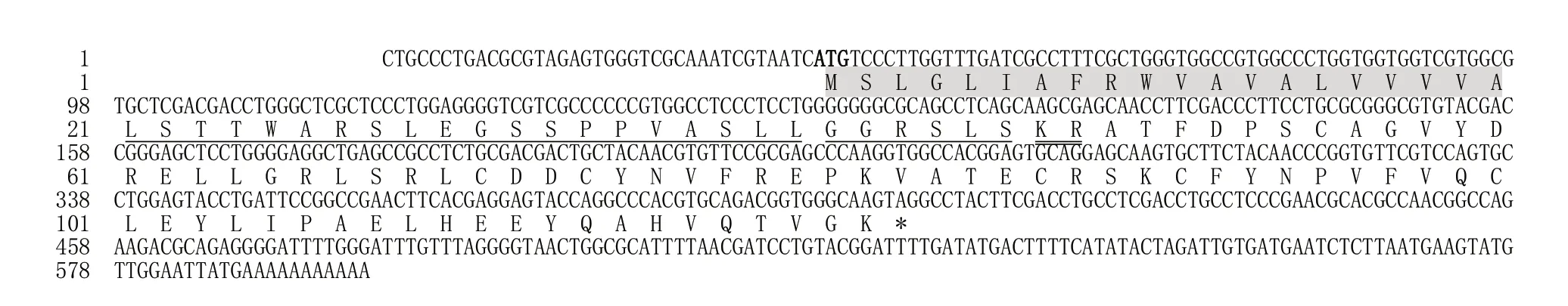
图1 墨吉明对虾FmCHH-I基因的cDNA和氨基酸序列Fig.1 Amino acid and cDNA sequence of gene FmCHH-I in Fenneropenaeus merguiensis

图2 墨吉明对虾FmCHH-I蛋白信号肽预测Fig.2 Signal peptide prediction of FmCHH-I in Fenneropenaeus merguiensis
2.2 FmCHH-I基因的理化性质分析
经预测,FmCHH-I基因编码的蛋白分子质量为13.5 ku,理论等电点为7.56。脂溶指数为93.44,不稳定系数为64.58,被划分为不稳定蛋白。总平均亲水性系数为0.157,最高亲水位在第15 位为3.622,最高疏水位在第112 位为-1.789,即此蛋白为亲水性蛋白(图3)。

图3 FmCHH-I蛋白疏水性Fig.3 Hydrophobicity for the FmCHH-I protein
2.3 FmCHH-I基因的同源性
FmCHH-I与GenBank 上部分物种的CHH-I同源序列比对结果见图4。CHH-I 蛋白序列在不同甲壳动物之间较为保守,FmCHH-I与对虾科物种相似性较高,与斑节对虾(Penaeus monodon)的同源性最高,与热液口虾(Rimicaris kairei)的同源性最低,序列中6个半胱氨酸残基在不同物种间高度保守(图4),系统发育分析(图5)表明,FmCHH-I 与斑节对虾聚在一起,与对虾科物种聚为一支,与斑节对虾、凡纳滨对虾(Litopenaeus vannamei)、日本囊对虾(Marsupenaeus japonicus)和刀额新对虾(Metapenaeus ensis)等物种的CHH基因相似性较高。这些同源基因在所选物种氨基酸序列的后半部分均有相同的功能结构域(图5)。

图4 墨吉明对虾FmCHH-I蛋白的多序列比对Fig.4 Multiple sequence alignment of the FmCHH-I protein of Fenneropenaeus merguiensis

图5 墨吉明对虾FmCHH-I基因的系统进化分析和蛋白结构域预测Fig.5 Phylogenetic analysis of FmCHH-I gene and protein structural domain prediction
2.4 FmCHH-I基因的表达特征
图6 表明,FmCHH-I基因在墨吉明对虾眼柄中表达量最高,在肝胰腺、肠道中表达量次之,在其他组织中无明显表达或表达量极低。

图6 墨吉明对虾CHH-I的组织表达Fig.6 Tissue distribution of FmCHH-I
图7(a)显示,FmCHH-I基因在雌性对虾卵巢未发育期的表达量与性腺成熟期的表达量无显著差异。处于性腺成熟期的墨吉明对虾在切除眼柄后,FmCHH-I基因的表达量远远高于未切除眼柄对虾的表达量(P<0.01)(图7(b))。
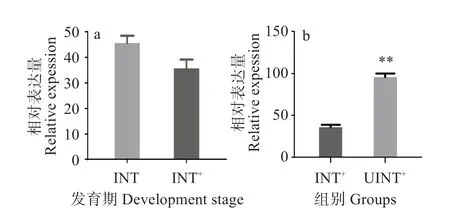
图7 性腺发育及切除眼柄对FmCHH-I表达量的影响Fig.7 Effect of gonad development and eyestalk abloation on FmCHH-I expression
3 讨论
甲壳类高血糖激素(CHH)家族神经肽参与调控甲壳动物多种生理过程。许多学者研究了该超基因家族在甲壳动物中的功能。在日本沼虾(Macrobrachium nipponense)[11]、斑节对虾(Penaeus monodon)[12]、罗氏沼虾(Macrobrachium rosenbergii)[13]等虾蟹类中克隆出了CHH家族基因的序列,通过对比发现,不同物种间该基因结构存在差异,但总体保持着CHH家族基因功能结构域的保守性,如均有信号肽、成熟肽中均有6个半胱氨酸残基等,这些特征也被证明是CHH基因超家族的典型特征[14]。
本研究首次在墨吉明对虾中克隆到CHH 家族神经肽成员FmCHH-I基因的cDNA全长,为599 bp,其中5′-UTR 37 bp,3′-UTR 193 bp,ORF为369 bp,编码123 个氨基酸,在第1~20 位包含一个信号肽,中间有一个由26个氨基酸组成的前导肽,随后是一段74个氨基酸组成的成熟肽,无跨膜区,含有6个半胱氨酸残基,成熟肽中第12位甘氨酸残基缺失,且有二盐基切割位点KR,为I型CHH神经肽的典型特征。
FmCHH-I 与CHH-II 蛋白氨基酸序列比对可见,CHH 神经肽中半胱氨酸的含量高,且序列高度保守。系统进化分析发现,CHH-I基因在对虾科中极为保守。FmCHH-I与斑节对虾、凡纳滨对虾、日本囊对虾和刀额新对虾的CHH 同源基因均有很高的相似性,且序列中的半胱氨酸高度保守,6 个半胱氨酸残基形成3 个二硫键结构,起稳定肽链空间结构的作用。由此推断,半胱氨酸在CHH神经肽的生物学功能保守性方面扮演着重要角色。
本研究的半定量RT-PCR 检测结果显示,FmCHH-I基因在墨吉明对虾眼柄中表达量最高,在肝胰腺、肠道中也有表达,这与罗氏沼虾[13]的结果一致。FmCHH-I在眼柄中高表达可能是因为,该基因主要由眼柄中的X 器官窦腺复合体合成和释放,再通过血淋巴传递到其他组织而发挥功能[17]。但Liu 等在凡纳滨对虾的组织中发现,CHH基因在其心脏中的表达量最高,推测可能由物种间的差异造成[15]。Sedl-Meier等[16]研究发现,CHH-I基因能调节甲基法尼脂的分泌,并影响肝胰腺分泌消化酶,调控中肠释放淀粉酶,由此解释了在肝胰腺、肠道中有CHH基因表达的现象。
研究发现,在一些物种中CHH-I在雄性精巢中的表达量远高于卵巢,由此推测CHH-I基因可能与雄性的发育密切相关[18]。对虾养殖人员在生产上常常采取切除单侧眼柄的方法促进卵巢成熟。本研究中,雌性对虾性腺成熟期FmCHH-I基因的表达量与未发育期无显著差异;处于性腺成熟期的墨吉明对虾在切除眼柄后,FmCHH-I基因表达量远高于未切除眼柄对虾中的表达量,说明CHH-I基因不仅与对虾体内血糖调节有关,还可能参与调控卵巢成熟过程。推测主要有两方面原因。首先,在许多物种中CHH-I基因都有多种形式的可变剪切体或亚型,如美洲龙虾(Homarus americanus)CHH-I基因就存在着CHH-A和CHH-B两种形式,只有CHH-B对卵巢发育有促进作用[19]。Kleijn 等[20]在雌性美洲龙虾生殖周期的研究中发现,X 器官/窦腺复合体中的CHH-A只在卵黄发生前期表达量较高,CHH-B则在卵黄发生前期与成熟期表达量较高。而本研究的FmCHH-I基因可能为CHH-I型基因中的一个亚型,因此在雌性对虾性腺成熟期与未发育时期的表达量无显著差异。
此外,对虾性腺成熟期需要积累大量营养物质,这一时期对虾所需要的营养物质远高于生长期[21]。切除单侧眼柄的目的是为了刺激对虾性腺发育和繁殖,而移除储存和分泌CHH 神经肽的X 器官/窦腺复合体后,对虾体内的营养物质不能及时吸收和转移,当体内能量不足时血糖含量会下降,此时需要大量的CHH-I 蛋白发挥作用,刺激机体产生更多的葡萄糖,从而满足自身能量需求。
综上,本研究首次在墨吉明对虾中克隆得到FmCHH-I基因的cDNA 全长,多序列比对、系统进化和基因表达分析表明,FmCHH-I基因的功能较为保守,在对虾的发育过程中可能起调节血糖以及参与性腺发育调控的作用,但此作用过程还需进一步的研究验证。
