低温胁迫对军曹鱼幼鱼血清生化指标、肝脏抗氧化酶活性及凋亡相关基因表达量的影响
2022-09-28黄建盛陈有铭温震威欧光海黄鉴鹏蒋鑫涛郭潮安
李 豫,黄建盛,陈有铭,温震威,欧光海,黄鉴鹏,蒋鑫涛,郭潮安,马 骞,陈 刚,3
(1.广东海洋大学水产学院,广东 湛江 524088;2.广东蓝粮种业有限公司,广东 湛江 524000;3.广东省水产动物病害防控与健康养殖重点实验室,广东 湛江 524088)
温度是影响鱼类生长、发育和繁殖的主要环境因子。温度骤降对鱼类正常生命活动产生不利影响,甚至导致鱼类大量死亡。尼罗罗非鱼(Oreochromis niloticus)[1]、杂交石斑鱼(Epinephelus moara♀×Epinephelus lanceolatus♂)[2]、点带石斑鱼(Epinephelus coioides)[3]、鲤(Cyprinus carpiovar.wuyuanensis♂×Cyprinus pellegrini pellegrini♀)[4]等在低温胁迫下,葡萄糖(GLU)、甘油三酯(TG)和谷丙转氨酶(ALT)等血清生理生化指标均发生显著变化。此外,低温胁迫还会导致鱼体内源性活性氧(ROS)增加,造成氧化损伤[5]。将斑马鱼(Danio rerio)和欧洲绵鳚(Zoarces viviparus)直接转入低温环境中导致脑和肝脏发生氧化损伤[6-7],长期处于低温胁迫的滨岸护胸鲇(Teleostei callichthyidae)和欧洲舌齿鲈(Dicentrarchus labrax)中也出现类似情况[8-9]。鱼类通过抗氧化防御系统中的过氧化氢酶(CAT)、超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GPx)等抗氧化酶消除ROS。当ROS含量超出机体清理能力范围时,将引起氧化应激反应,导致细胞膜损伤、酶失活以及遗传物质等重要细胞成分受损[10-11]。氧化应激可进一步诱导细胞凋亡[12]。大多数凋亡信号转导过程与caspase家族、p53、mdm2、bax和bcl-2等凋亡相关基因有关,关键凋亡相关基因表达量的变化可降低细胞活力,增加细胞凋亡比例[13-14]。
军曹鱼(Rachycentron canadum)营养丰富,肉质鲜美,经济价值高,是近海浮动式网箱和深海网箱的重要养殖鱼种[15]。据报道,军曹鱼适宜生长水温为25~32℃,水温低至21 ℃时摄食量明显降低,18 ℃时静止于水底[16]。近年来,我国南方地区冬春季节寒潮天气频发,对军曹鱼养殖业造成一定的经济损失。本实验室前期开展了低温胁迫影响军曹鱼幼鱼脂代谢的相关研究,发现军曹鱼通过显著提高肝脏和腹腔脂肪的多不饱和脂肪酸比例来响应低温胁迫[17]。本研究测定低温胁迫下军曹鱼的血清和肝脏生理生化指标,检测肝脏中凋亡相关基因表达量的变化,为军曹鱼健康养殖提供参考。
1 材料与方法
1.1 实验材料及实验设计
实验用军曹鱼幼鱼由广东海洋大学海洋生物研究基地自繁自育,实验在广东恒兴饲料股份有限公司863 基地进行。实验开始前将幼鱼暂养两周,养殖设备为24 h 的流水养殖系统,每个水槽300 L,溶氧水平≥5 mg/L,氨氮≤0.02 mg/L,pH 7.6±0.2,温度(28±1)℃、盐度29±1。暂养结束后挑选健康、规格一致、平均体质量(39.11±1.01)g、平均体长(19.71±0.73)cm 的军曹鱼进行实验。实验设置(18±0.5)、(21±0.5)、(25±0.5)℃3 个温度处理组和1个对照组(28±1)℃,每组3个重复,每个重复20尾鱼。降温程序参考文献[18-20]并适当调整,水温高于23℃时以6 h/℃匀速降温,降至23℃后以12 h/℃匀速降温。温度处理组,将冰块置于4层虾苗袋,密封,放入水槽,水槽上方覆盖薄膜,并在水槽周围粘贴隔热棉以减少温度波动;使用气石连续充气保证水体温度均衡;实验期间24 h连续监测温度值(每2 h测量一次水温),当水温偏离0.3 ℃以上时即调整冰块数量。实验期间低温处理组静水养殖,每天在8:00和17:00投喂饲料(广东东腾饲料有限公司),每天更换50%的等温海水,除水温外,其余水质条件与暂养期间保持一致。
1.2 样品采集
当温度处理组水温降至指定温度(25±0.5、21±0.5、18±0.5)℃后,在0、4 和7 d 时每组随机取鱼15尾(每个重复5 尾),使用40 mg/L 的丁香酚麻醉,用1 mL 注射器尾静脉处采血,离心(3 500 r/min,15 min,4℃)取上层血清置-80 ℃冰箱中保存,用于检测生理生化指标;采血后每组随机取12尾,6尾鱼肝脏用于酶活测定,6 尾鱼肝脏用于基因表达分析。所取组织经液氮速冻后置于-80 ℃冰箱冻存备用。
1.3 血清生理生化指标及肝脏酶活测定
血清葡萄糖、甘油三酯、总胆固醇(T-CHO)、总蛋白(TP)、谷丙转氨酶、谷草转氨酶(AST)、碱性磷酸酶(AKP)指标使用全自动分析仪测定(Chemray 240,深圳雷杜生命科技),所用试剂盒购自南京建成生物工程有限公司。肝脏过氧化氢酶、超氧化物歧化酶、谷胱甘肽过氧化物酶活性和丙二醛(MDA)含量均用南京建成生物工程有限公司试剂盒,参照试剂盒说明书测定。
1.4 基因表达分析
肝脏总RNA 提取、质量检测、反转录操作流程均参照文献[21]。根据军曹鱼已有的全基因组数据(NCBI BioProject ID:PRJNA634421)查找目的基因序列,使用Primer Premier 6.0 设计实时荧光定量PCR(qRT-PCR)引物(表1)。以获得的肝脏cDNA为模板,β-actin为内参基因,通过qRT-PCR 分析bax、bcl-2、caspase-9、caspase-3、p53、mdm2基因在不同温度条件下的表达变化。qRT-PCR 反应程序:94 ℃30 s;94 ℃5 s、58 ℃15 s、72 ℃10 s,40循环。用2-ΔΔCt法计算基因的相对表达量。

表1 荧光定量引物Table 1 Primers used for quantitative real-time PCR
1.5 数据分析
实验数据采用单因素方差分析和Duncan's 法多重比较进行显著性差异分析,P <0.05 表示差异显著,采用GraphPad Prism 8软件作图。
2 结果
2.1 低温胁迫对军曹鱼幼鱼血清生化指标的影响
图1 可见,胁迫0 d 时,18 ℃组血清GLU 含量显著高于其他组(P <0.05);4、7 d 时,28 ℃组GLU含量显著高于温度处理组(P <0.05)(图1(A))。18 ℃组血清TG 含量在各个时间点均显著高于28 ℃和25 ℃组(P <0.05)(图1(B))。21 ℃和18 ℃组T-CHO 含量在各个时间点均显著低于28 ℃和25 ℃组(P <0.05)(图1(C))。0、4 d 时,组间血清TP含量无显著性差异(P >0.05);7 d时,21℃和18℃组TP 含量均显著低于28℃和25℃组(P <0.05)(图1(D))。

图1 低温胁迫对军曹鱼幼鱼血清葡萄糖、甘油三酯、总胆固醇和总蛋白含量的影响Fig.1 Effects of low-temperatures on contents of glucose(GLU),triglyceride(TG),total cholesterol(T-CHO)and total protein(TP)in serum of juvenile cobia
2.2 低温胁迫对军曹鱼幼鱼血清酶活性的影响
图2 可见,胁迫0、4 d 时,18 ℃组血清ALT 活性显著高于其他组(P <0.05);7 d 时,18 ℃组ALT 活性下降且组间无显著差异(P >0.05)(图2(A))。0 d时,28 ℃组血清AST活性显著低于温度处理组(P <0.05);4 d 时,21 ℃和18 ℃组AST 活性均显著高于28 ℃和25 ℃组(P <0.05),而在7 d 时,仅21 ℃组显著高于其他组(P <0.05)(图2(B))。0、4 和7 d时,血清AKP 活性在28 ℃和25 ℃组较稳定,21 ℃和18 ℃组分别在4 d 和7 d 达到峰值并显著高于其他组(P <0.05)(图2(C))。

图2 低温胁迫对军曹鱼幼鱼血清谷丙转氨酶、谷草转氨酶和碱性磷酸酶活性的影响Fig.2 Effects of low-temperatures on activities of alanine aminotransferase(ALT),aspartate aminotransferase(AST)and alkaline phosphatase(AKP)in serum of juvenile cobia
2.3 低温胁迫对军曹鱼幼鱼肝脏抗氧化酶活性的影响
图3 可见,在0、4、7 d 时,28 ℃和25 ℃组肝脏SOD 活性较稳定且显著高于21 ℃和18 ℃组(P <0.05)(图3(A))。0 d 时,28 ℃组肝脏CAT 活性显著高于温度处理组(P <0.05);4、7 d 时,28 ℃和25 ℃组肝脏CAT 活性显著高于21 ℃和18 ℃组(P <0.05)(图3(B))。0 d 时,18 ℃组肝脏GPx 活性显著高于其他组(P <0.05);21 ℃和18 ℃组GPx 活性在4 d 时显著高于28 ℃和25 ℃组,而在7 d 时,则显著低于后者(P <0.05)(图3(C))。7 d时,21 ℃和18 ℃组肝脏MDA 含量均显著高于28 ℃和25 ℃组(P <0.05)(图3(D))。
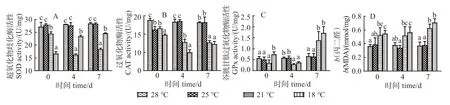
图3 低温胁迫对军曹鱼幼鱼肝脏超氧化物歧化酶、过氧化物酶、谷胱甘肽过氧化物酶活性及丙二醛含量的影响Fig.3 Effects of low-temperatures on activities of SOD,CAT,GPx and contents of MDA in liver of juvenile cobia
2.4 低温胁迫对军曹鱼幼鱼肝脏凋亡基因相对表达量的影响
图4 可见,在0、4 和7 d 时,21 ℃和18 ℃组肝脏caspase-3和caspase-9基因表达量均显著高于28 ℃和25 ℃组(P <0.05)(图4(A,B))。28 ℃组肝脏p53和bax基因表达量均显著低于温度处理组(P <0.05)(图4(C,F))。胁迫0 d 时,25 ℃和18 ℃组肝脏mdm2基因表达量显著高于28 ℃和21 ℃组(P <0.05);胁迫4、7 d 时,21 ℃和18 ℃组mdm2基因表达量显著高于28 ℃和25 ℃组(P <0.05)(图4(D))。胁迫0、7 d 时,25 ℃组肝脏bcl-2基因表达量显著高于其他组,18 ℃组显著低于其他组(P <0.05);胁迫4 d 时,25 ℃组显著高于21 ℃和18 ℃组(P <0.05)(图4(E))。

图4 低温胁迫对军曹鱼幼鱼肝脏组织凋亡相关基因caspase-3、caspase-9、p53、mdm2、bcl-2和bax表达量的影响Fig.4 Effect of low-temperatures on relative expression of genes caspase-3,caspase-9,p53,mdm2,bcl-2 and bax in liver of juvenile cobia
3 讨论
3.1 低温胁迫对军曹鱼幼鱼代谢水平的影响
血液是鱼体内循环系统的重要组成部分,具有生理调节、生理防御和物质运输等功能,其生理生化指标直接反映鱼类的营养状况和代谢水平[22]。GLU 为机体主要的能源物质,是鱼类维持正常生命活动的能量直接来源[23]。本研究中,军曹鱼幼鱼血清GLU 含量在18 ℃和21 ℃水温胁迫下呈先升高后降低的趋势,这与鲤(Cyprinus carpio)[4]的低温胁迫结果相似,其原因可能是低温处理初期鱼体内大量肝糖原转化为GLU以满足机体的代谢需求,而后大量GLU 分解成ATP 为机体提供能量以抵御低温环境[4]。TG 和T-CHO 作为血脂的主要成分[24],为鱼体提供能量并维持体温,可作为衡量脂代谢的指标[25]。在18 ℃和21 ℃水温胁迫下幼鱼血清TG 含量显著高于对照组,与本实验室前期开展的军曹鱼低温胁迫研究结果一致,推测低温胁迫会诱导鱼体合成更多的脂肪酸以抵御寒冷[17]。低温胁迫下鱼体肝脏会受到损伤,导致与合成T-CHO 相关的酶活性受抑制,使T-CHO 合成速度减慢,同时肝脏损伤导致其代谢功能障碍,T-CHO 不能正常通过肝肠循环被肝脏重新吸收,进而引起血清T-CHO含量降低[22]。本研究中,军曹鱼幼鱼在低温胁迫后血清T-CHO 含量降低,推测与低温导致其在幼鱼体内合成速度下降及其在肝肠循环中代谢异常有关。
血清蛋白质水平可反映鱼体健康状态,TP具有稳定血液胶体渗透压和酸碱度的作用[26]。褐篮子鱼(Siganus fuscescens)[27]和七带石斑鱼(Epinephelus septemfasciatus)[28]的血清TP 含量随着低温作用强度的增强呈降低的趋势。本研究中,军曹鱼幼鱼血清TP 含量在18℃和21℃水温胁迫到7 d 时显著低于对照组,推测长时间低温胁迫可能造成肝脏功能受损,蛋白质合成速率下降[28];此外,低温胁迫后期幼鱼摄食量明显减少,也可能导致血清蛋白水平下降。
3.2 低温胁迫对军曹鱼幼鱼肝功能及免疫的影响
鱼类血清酶活性变化可作为判断组织器官的状态和代谢水平的依据[29]。ALT 和AST 是可用于指示肝细胞损伤及其程度的转氨酶。机体处于正常状态时肝细胞结构完整,仅少量转氨酶进入血液,当肝细胞损伤时细胞膜通透性改变,大量ALT和AST 被释放到血液中,导致其活性在血液中迅速升高[30]。在云纹石斑鱼(Epinephelus moara)的低温胁迫实验中,血清ALT 和AST 活性显著高于对照组[31],符合上述规律。本研究中,低温胁迫引起军曹鱼幼鱼ALT 和AST 活性显著升高,胁迫7 d 时降至对照组水平,可能是低温胁迫前期肝脏合成转氨酶过程没有受到抑制,肝脏受损后大量转氨酶进入血清,而在胁迫7 d 幼鱼响应低温刺激时合成转氨酶的过程受到抑制,使血液中转氨酶含量呈下降趋势[2]。
AKP 是鱼类重要的防御因子,广泛参与到免疫活动中,血清AKP 活性可作为评价机体免疫能力、代谢水平的指标[32]。机体代谢异常时AKP 通过淋巴道和肝窦释放到血液中,造成血清AKP 活性升高[28]。本研究中18 ℃和21 ℃水温胁迫7 d 时军曹鱼幼鱼血清AKP活性显著高于对照组,推测低温胁迫对军曹鱼幼鱼的免疫机制或代谢造成损伤。
3.3 低温胁迫对军曹鱼幼鱼抗氧化防御系统的影响
生物机体在环境刺激下会产生大量ROS,而过量的ROS 则会造成DNA、蛋白质和脂质的功能受损[33]。鱼体抗氧化防御系统中,抗氧化酶SOD、CAT、GPx 和部分非酶类抗氧化剂可相互协调清除ROS,防止或修复氧化损伤[34]。研究表明,鱼类抗氧化系统易受温度影响,大口黑鲈(Micropterus salmoides)长期处于温度胁迫状态会造成应激损伤并降低免疫水平[35];对中华鲟(Acipenser sinensis)进行急性低温胁迫后其氧化酶活性改变并对肝脏产生一定程度的损伤[36];点篮子鱼(Siganus guttatus)幼鱼肝脏抗氧化酶活性在低温胁迫(14 ℃)后显著降低[37];在云纹石斑鱼[38]和尼罗罗非鱼[39]的研究中发现,低温胁迫导致鱼体内MDA 浓度升高,从而对鱼体造成伤害。本研究中,18 ℃和21 ℃组军曹鱼幼鱼肝脏SOD 和CAT 活性在胁迫期间均显著低于对照组,其原因可能是低温胁迫造成肝功能受损,抗氧化防御能力降低,从而导致SOD 和CAT 活性下降。而GPx 活性在低温胁迫至7 d 时显著高于对照组,可能是胁迫后期CAT 活性下降诱导GPx活性上升以清除过量的H2O2[40]。MDA 含量在胁迫期间均显著高于对照组,表明低温胁迫下军曹鱼幼鱼抗氧化防御能力遭到破坏,机体无法及时清除过量的ROS,体内ROS 浓度上升使脂质过氧化程度加深。
3.4 低温对军曹鱼幼鱼肝脏组织凋亡基因相对表达量的影响
细胞应对刺激有不同的保护机制,但超出细胞承受范围的刺激强度会导致细胞信号中断、DNA 结构破坏和细胞凋亡[12]。研究表明,氧化应激会改变caspase家族、p53、mdm2、bax和bcl-2等凋亡相关基因表达水平,进而诱导细胞凋亡[13]。细胞应激产生的ROS 会改变p53表达水平,进而启动细胞周期停滞或凋亡程序[41]。p53为信号网络中心枢纽,可调控细胞的活性,其中p53-mdm2信号通路是主要的调控途径[42]。mdm2在调控p53表达水平、维持机体的稳态发挥着重要作用,p53被激活后促进mdm2表达,mdm2表达产物对p53具有负调控作用[43]。本研究中,18 ℃和21 ℃水温胁迫导致军曹鱼幼鱼p53表达量显著高于对照组,提示p53可能在低温诱导幼鱼细胞凋亡中起关键作用,其表达量变化可能启动细胞凋亡程序,同时mdm2的激活可能是受p53表达上调的影响。
p53可通过上调bax和下调bcl-2表达诱导细胞凋亡。bax和bcl-2均属bcl2家族,但基因功能完全不同[44]。研究表明,低温胁迫导致四川华鳊(Sinibrama taeniatus)细胞凋亡,其凋亡相关基因p53和bax表达上调以及bcl-2表达下调,本研究检测的基因表达趋势与此结果相似,推测低温胁迫下军曹鱼发生了细胞凋亡[45]。
细胞凋亡可由多种刺激触发,但凋亡途径主要有外源性死亡受体途径和内源性线粒体途径[46]。内源性线粒体途径通过激活caspases-9启动凋亡程序,caspases-9表达上调可以激活下游caspases-3促使细胞凋亡,caspases-3是两种凋亡途径的“执行者”。本研究中,在18 ℃和21 ℃水温胁迫下军曹鱼幼鱼肝脏caspases-9和caspases-3表达显著高于对照组,说明低温胁迫下最终激活caspases-3表达,进而激活内源性线粒体途径引发细胞凋亡[47]。此结果与低温胁迫下暗纹东方鲀(Takifugu obscurus)[48]和点带石斑鱼[3]caspases-9和caspases-3表达显著上调结果相似。鱼类受到外来刺激时,会造成体内ROS 代谢紊乱,细胞组分及生物合成机制发生异常,进而可能诱发细胞凋亡[49]。推测可能是低温胁迫军曹鱼幼鱼过程中,抗氧化防御能力降低,细胞不断受到氧化损伤,从而导致细胞凋亡[11]。
3.5 军曹鱼对低温的敏感性
低温胁迫显著影响鱼类生理过程,降温速率对于鱼类响应低温胁迫过程的形态、生理、基因表达水平的变化有决定性作用;因此,需依据目标物种的温度适应性等特征选取合适的降温速率。例如,花鲈(Lateolabrax maculatus)[20]、七彩神仙鱼(Symphysodon aequifasciatus)[19]、四带无须鲃(Puntius tetrazona)[50]等分别选取1 ℃/h、1 ℃/d及2 ℃/d 的降温速率开展低温胁迫处理。军曹鱼广泛分布在水温较高的海域[51],对低温较为敏感,过快的降温速率(2 ℃/h)会导致军曹鱼不摄食[17],故本研究首先以6 h/℃匀速降温至23 ℃后,再以12 h/℃匀速降温。此外,上述研究中低温处理组的首次取样均选取在温度降至目标温度后开始,因此,本研究也采取相似的取样策略,以保证研究结果的可比性。
4 结论
本研究表明,低温胁迫会显著抑制军曹鱼幼鱼代谢水平,降低其抗氧化相关酶活性,造成氧化损伤,从而诱导凋亡相关基因表达,促进细胞凋亡。
