耐低pH值纳他霉素生产菌株的诱变选育
2022-09-28王鑫宇许倍铭刘陈梦吴康孙瑞杰贾士儒谭之磊
王鑫宇,许倍铭,刘陈梦,吴康,孙瑞杰,贾士儒,谭之磊
(天津科技大学生物工程学院,天津 300457)
纳他霉素是天然多烯大环内脂类抗生素,分子式为C33H47NO13,分子量为665.75[1],含有4个共轭双键的26元内酯环,可以与麦角甾醇基团结合,从而使真菌细胞膜变形[2],进而引起菌内维持细胞生长的物质渗出,最终致使细胞死亡,但对细菌没有抑制作用[3]。纳他霉素难溶于水和油脂,很难被人体消化吸收,大部分摄入的纳他霉素会随粪便排出,因此具有很高的安全性[4-5]。由于纳他霉素具有优良的抗菌性能,目前除在焙烤食品、乳酪制品、易发霉食品等食品行业上应用外[6-7],在医药工业、饲料工业等领域也得到了越来越多的重视,临床上现已用于治疗皮肤真菌感染疾病、真菌性眼角膜炎等[8-9]。
褐黄孢链霉菌是主要的纳他霉素生产菌之一,褐黄孢链霉菌在发酵前期pH值快速下降,但酸性条件不利于菌体生长和纳他霉素生产[10]。因此筛选耐受低pH值的菌株不仅有利于纳他霉素生产,也有利于在实际生产中减少耗碱量,降低生产过程中粗放操作对发酵产量造成的影响。Zhang等[11]等以干酪乳杆菌为出发菌,在pH4.3的酸性条件下适应性进化70 d得到的进化株,摇瓶发酵64 h,生物量较出发菌提高60%以上。Nyabako等[12]通过常压室温等离子体诱变结合酸适应进化,有效提高了嗜酸乳杆菌的耐酸能力。突变株LAartp-ale2在pH3.0和pH2.5的酸性条件下存活率显著提高。目前影响链霉菌耐酸性的相关报道较少。但在部分革兰氏阳性菌中已有研究表明[13],胞内质子(H+)的去除,细胞膜组分改变及胞内三磷酸腺苷(adenosine triphosphate,ATP)含量等都会影响菌株的耐酸能力,通过将胞内H+排出胞外可以维持胞内pH值稳态,从而保证菌体在酸性环境中的正常生长,而这一过程需要ATP提供能量。此外ATP通常作为微生物生长和生物合成有用物质所必需的能量来源,纳他霉素生物合成酶的作用的发挥需要ATP提供能量[14]。
本试验以褐黄孢链霉菌S.gilvosporeus TUST01为出发菌,通过诱变育种结合合理地筛选方式,获得耐低pH值突变菌株S.gilvosporeus Y-10,并对其耐酸机制进行研究。以期为降低纳他霉素生产成本,研究链霉菌耐酸机制提供参考。
1 材料与方法
1.1 主要试剂
甲醇(色谱纯)、氯仿(分析纯):康科德科技有限公司;氢氧化钠(分析纯):博欧特化工贸易有限公司;葡萄糖(分析纯)、磷酸盐缓冲液(分析纯)、ATP检测试剂盒:北京索莱宝生物科技有限公司。
1.2 菌株
纳他霉素原始生产菌株Streptomyces gilvosporeus TUST01:天津科技大学生化工程实验室保存。
1.3 培养基
种子培养基:淀粉10 g,葡萄糖10 g,黄豆饼粉10 g,蛋白胨 6 g,玉米浆 6 g,硫酸镁 1 g,磷酸二氢钾0.5 g,氯化钠2 g,碳酸钙5 g,加水定容至1 L,调节pH值为7.0。
发酵培养基:葡萄糖40g,淀粉30g,牛肉膏10g,黄豆饼粉15g,酵母抽提物0.3g,蛋白胨6.3g,MgSO4·7H2O 1.0g,NaCl2g,加水定容至1L,调节pH值为4.4和7.4。
斜面培养基:淀粉10g,葡萄糖10g,黄豆饼粉10g,酵母粉3 g,麦芽粉3 g,硫酸镁1 g,磷酸氢二钾0.5 g,氯化钠2 g,碳酸钙3 g,琼脂20 g,加水定容至1 L,调节pH值为7.0。
低pH值固体筛选培养基为斜面培养基,pH值调整为4.0。
以上培养基均使用121℃高压蒸汽灭菌20 min。
1.4 仪器与设备
BASIC紫外分光光度计:德国Eppendorf公司;SBA-400E生物传感分析仪:山东省科学院生物研究所;BIOTECH-5BG-7000A 5 L发酵罐:上海保兴生物设备工程有限公司;ARTP-Ⅱ常压室温等离子体诱变系统:北京思清源生物科技有限公司;Infinite 200PRO多功能酶标仪:帝肯贸易有限公司。
1.5 方法
1.5.1 菌株活化
在超净台中无菌条件下打开保藏菌种干粉的安瓿管,在其中加入1mL0.9%无菌生理盐水,使用移液枪吹吸使菌种干粉在生理盐水中混合均匀。吸取200 μL菌液加入到100 mL种子培养基中,平行3份。将种子培养基在28℃、220 r/min的摇床中培养48 h。
使用无菌接种环在菌种的复苏种子培养基中挑菌,在固体培养基平板上三区划线,平行3份,29℃恒温培养5 d~7 d直至菌种表面产生孢子。
1.5.2 常压室温等离子体诱变(atmospheric and roomtemperature plasma,ARTP)
用无菌生理盐水冲洗S.gilvosporeus TUST01斜面上生长良好的孢子,置于装有玻璃珠的250 mL三角瓶中,220 r/min振荡打散30 min后用滤膜过滤,调整孢子悬浮液浓度约为108CFU/mL。取10 μL孢子悬浮液于无菌金属载片上,放置于ARTP-Ⅱ常压室温等离子体诱变系统处理台座,在电源功率40 W,照射距离2 mm,气流量10 L/min条件下对菌悬液进行50 s的诱变处理[15],此时致死率为90%(正突变率最高)[16]。照射后,将金属载片放入装有1 mL无菌生理盐水的离心管中振荡1 min,再用生理盐水稀释后涂布于pH4.0的低pH值筛选平板,29℃培养5 d~10 d至突变株单菌落孢子丝生长良好。
1.5.3 耐酸菌株筛选
将pH4.0筛选平板上的突变株接种于24孔深孔板,每孔含3 mL种子培养基,用透气封板膜密封后28℃,220 r/min于摇床振荡48 h。之后取培养液以8%接种量接种于同样的装有3 mL发酵培养基的24孔深孔板中,摇床发酵96 h后吸取200 μL发酵液于96孔板中,4 000 r/min离心10 min,之后取40 μL上清液于新的96孔板中,以70%甲醇适当稀释后置于酶标仪测定OD303值[17],挑选OD303值较高的菌株进行平板验证,并进行摇瓶复筛及5 L发酵罐验证。
5 L发酵罐培养方法:初始装液量3 L,121℃高压蒸汽灭菌20 min,28℃,220 r/min种子培养48 h,种子接种量15%,初始搅拌转速200 r/min,罐压0.1 MPa,通气量1∶1,温度28℃,关联溶氧与转速以控制溶氧量在30%,流加80%葡萄糖以控制葡萄糖含量为20 g/L。定时取样测定发酵液的残糖[18]、生物量[19]和纳他霉素产量[20]。在此条件下通过控制不同的pH值条件以比较出发菌与筛选菌株的生长和纳他霉素生产情况[21]。
1.5.4 遗传稳定性
将复筛选出的菌株在斜面培养基上连续转接15代,挑取各代菌株进行摇瓶发酵试验,测定其生物量和纳他霉素发酵产量[22]。
1.5.5 胞内ATP含量测定
根据ATP检测试剂盒说明书的方法测定胞内ATP 浓度[23]。
1.5.6 细胞膜脂肪酸含量测定
参照Bo等[24]的方法,发酵液离心弃上清液后,用磷酸盐缓冲液反复洗涤菌体细胞两次,经液氮淬灭、提取液提取小分子物质、甲基化和硅烷化后,取上清液进行气相色谱-质谱 (gas chromatography-mass spectrometer,GC-MS)检测。
气相色谱条件:HP-5MS色谱柱(60 m×0.25 mm,0.25μm);升温程序为进样后70℃保持2min,以5℃/min的速度升温到290℃,保持3min,再降温到70℃。运行时间为52 min;载气为高纯氦气,恒定流速为1 mL/min;接口温度280℃。
质谱条件:电子轰击电离;离子源温度为250℃;离子电流为40 μA;电子轰击能量为70 eV;扫描质量范围 m/z为 50~800。
1.6 数据分析与统计
数据分析采用费舍尔最小显著差异法,P<0.05表示差异显著。
2 结果与分析
2.1 突变株的筛选
2.1.1 初筛
通过pH4.0筛选平板的筛选获得了420株突变菌株。按照1.5.3中的24孔深孔板筛选方法,根据纳他霉素产量筛选耐受pH4.4的菌株,耐低pH值菌株纳他霉素产量散点图见图1。
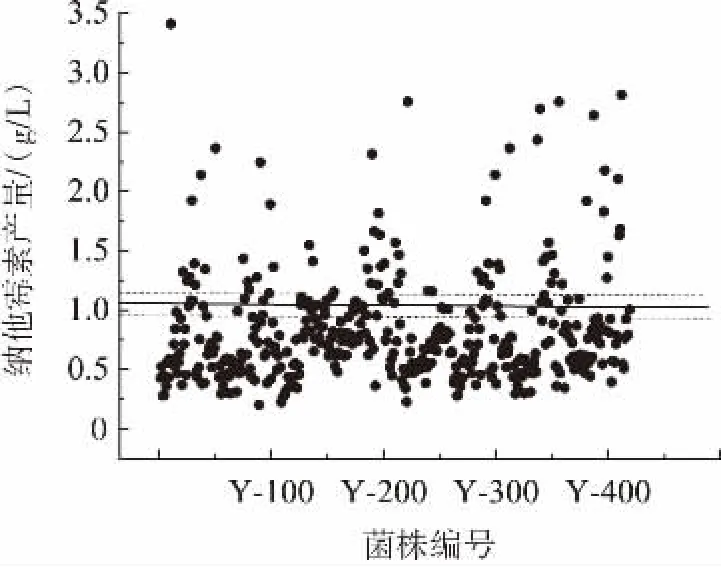
图1 耐低pH值菌株纳他霉素产量散点图Fig.1 Scatter plot of natamycin yield of strains with low pH resistance
由图1可知,206株突变株的纳他霉素与出发菌有显著差异,即与出发菌纳他霉素产量偏差超过±10%,突变率为49.05%。73株突变株较出发菌纳他霉素产量提高,正突变率达17.38%。其中6株突变株纳他霉素产量较高,菌株编号为Y-10,Y-226,Y-348,Y-354,Y-390以及Y-420,对6株菌株做进一步平板及摇瓶验证。
2.1.2 筛选菌株平板生长验证
对筛选菌株进行平板生长验证,如图2所示。
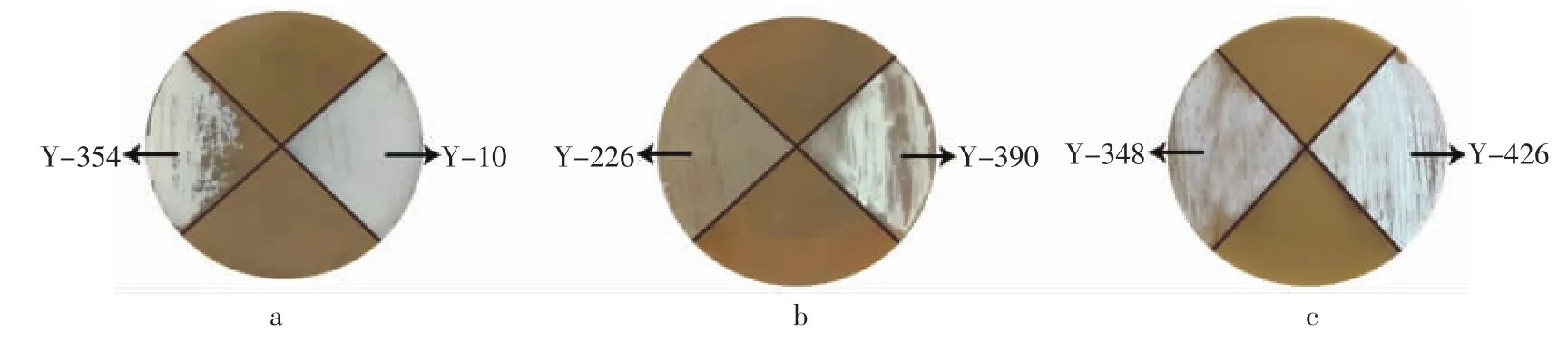
图2 筛选菌株平板验证图Fig.2 Plate verification of screened strains
由图2可知,筛选菌株中Y-10生长最为旺盛,此外Y-354,Y-390以及Y-420孢子发育良好,但菌落覆盖范围不完整。而Y-226与Y-348菌株外观形态异常,菌丝生长缓慢。其中Y-226只长菌但孢子完全未萌发,而Y-348仅有部分区域孢子萌发且发育较差。可见筛选菌株中Y-10耐酸效果最好,结合其较高的纳他霉素产量对Y-10进行进一步平板验证,观察其在pH4.4和pH7.4固体平板上不同培养时间的生长情况,结果见图3。
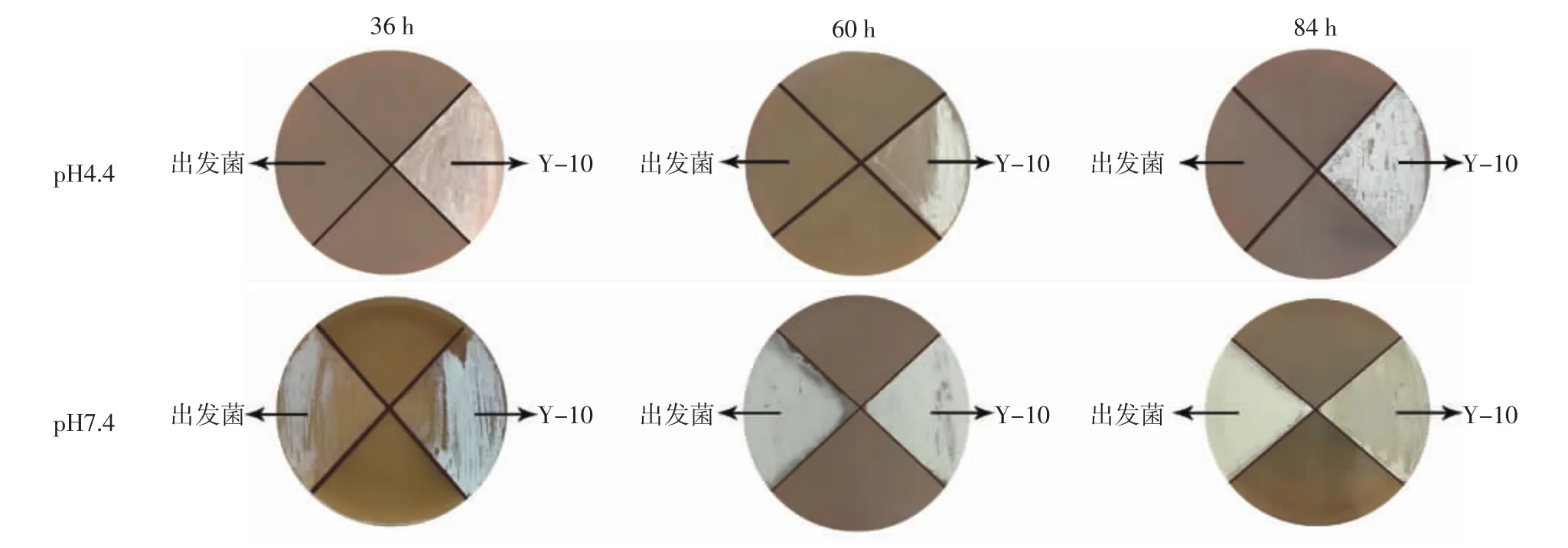
图3 Y-10与出发菌TUST01在不同pH值平板上的生长图Fig.3 Growth of Y-10 and TUST01 at different pH plates
由图3可知,出发菌在pH4.4的低pH值平板上不生长,而Y-10菌株生长旺盛,可见其具有较强的耐酸能力。此外,Y-10较出发菌在pH7.4的平板上培养前期生长更旺盛,到培养中期两株链霉菌孢子发育良好,而到后期Y-10菌株孢子变灰而出发菌孢子偏灰白色。
2.1.3 摇瓶复筛
2.1.3.1 pH4.4摇瓶复筛
6株突变株在24孔深孔板低pH值发酵培养基筛选条件下纳他霉素产量较高(图1),对6株突变株进行pH4.4摇瓶复筛,结果见图4。
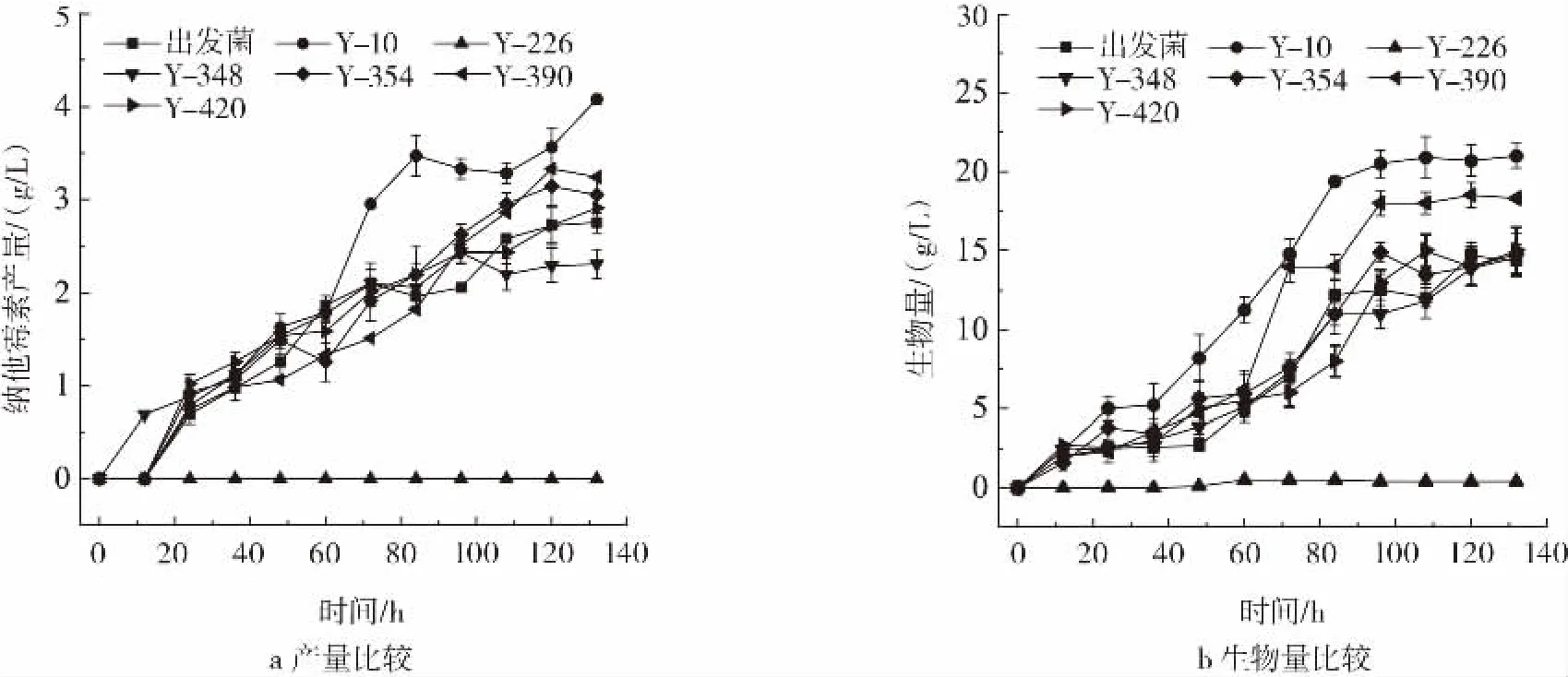
图4 耐低pH值菌株于pH4.4条件下纳他霉素产量和生物量验证Fig.4 Yield and biomass verification of the strain with low pH resistance at pH4.4 shaking flask
如图4a所示,Y-10在pH4.4条件下纳他霉素产量最高,于发酵132 h达最大值4.08 g/L,较出发菌TUST01提高48.36%。如图4b所示,Y-10于摇瓶发酵生物量最大值为20.89 g/L,较出发菌提高42.11%,与纳他霉素产量具有高度一致性。此外Y-226由于在平板上未产孢子,其在pH4.4摇瓶发酵条件下菌株难以生长,纳他霉素产量极低。因此选择Y-10作为优选耐酸菌株进行下一步研究。
将经过ARTP诱变及24孔深孔板初筛得到的在低pH值条件下纳他霉素产量较高的Y-10菌株与出发菌TUST01在pH4.4摇瓶发酵培养基中pH值和残糖进行比较,结果见图5。
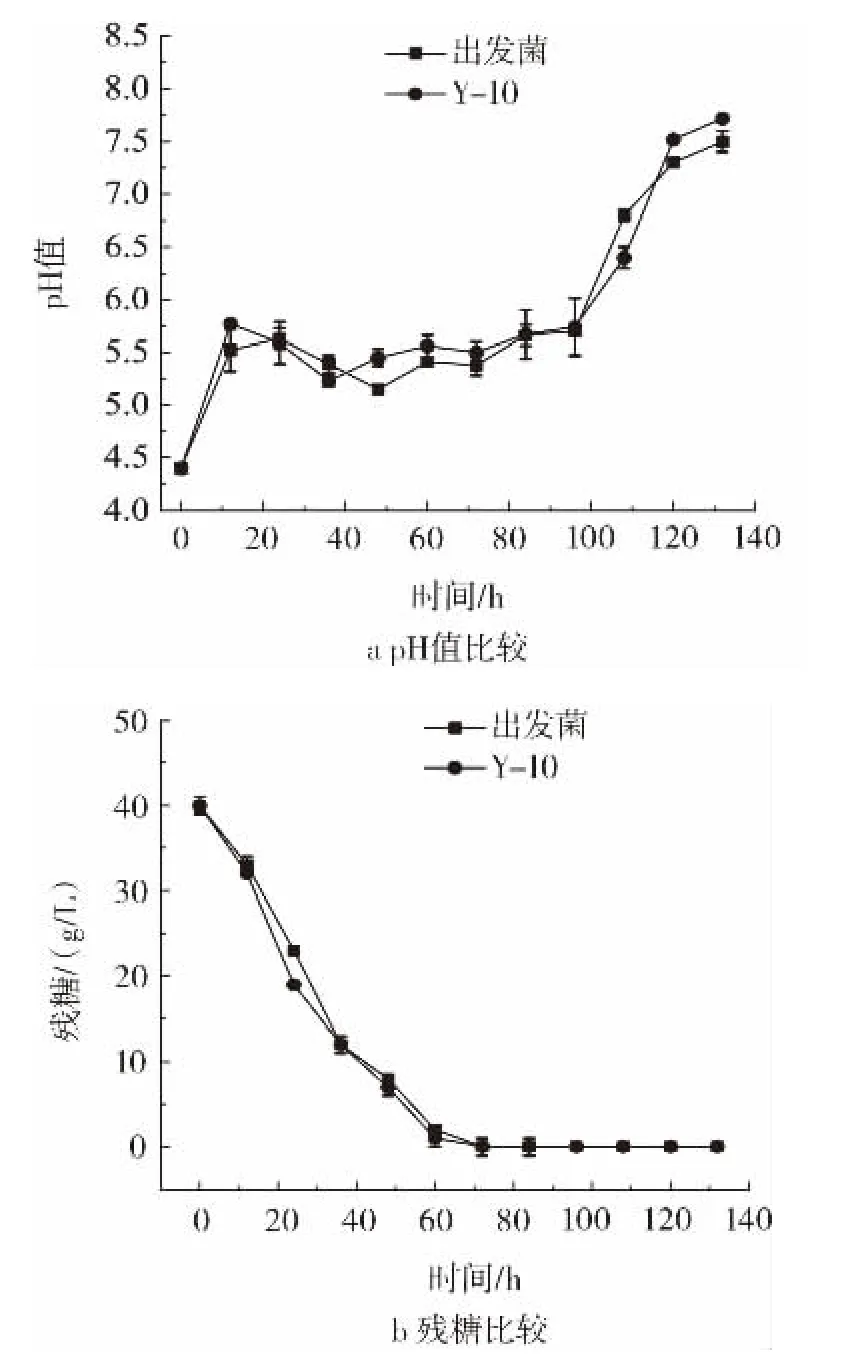
图5 Y-10与出发菌TUST01于pH4.4条件下摇瓶发酵参数比较Fig.5 Comparison of shaking flask fermentation parameters between Y-10 and the original strain at pH4.4
图5 a表明两株菌在低pH值条件下pH值都呈现出先升高后降低再升高的趋势,可能是由于发酵初期菌体出于满足细胞生长的要求,通过自身代谢调整pH值至近中性(5.63~5.77)。之后菌体生长产酸导致pH值降低,但是降低幅度很小。最后菌体自溶,碱性物质流出导致pH值上升。图5 b显示出两株菌的耗糖速度相当。由于Y-10较高的纳他霉素产量和生物量,确定Y-10可以耐受低pH值的条件,为了达到后期筛选耐受低pH值菌株以及减少发酵过程中碱的消耗的目的,还需对Y-10进行pH7.4的正常摇瓶发酵验证。
2.1.3.2 正常条件摇瓶复筛
对Y-10于pH7.4摇瓶条件下进行验证,比较Y-10与出发菌纳他霉素产量、生物量、pH值及残糖变化,结果见图6。
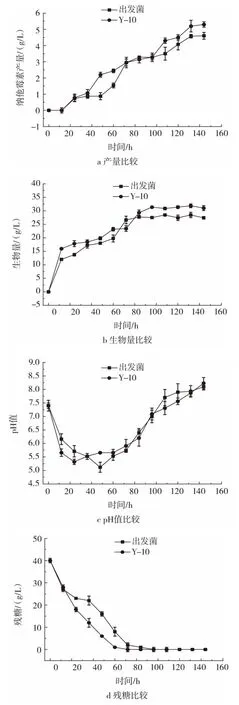
图6 Y-10与出发菌正常摇瓶发酵参数验证Fig.6 Verification of normal shaking flask fermentation parameters between Y-10 and the original strain
如图6 a所示,Y-10纳他霉素产量于发酵144 h达最高,为5.30 g/L,较出发菌株提高14.97%。如图6 b所示,Y-10发酵前期生物量增长速度较出发菌快,生物量于发酵96 h达最大,为31.85 g/L,较出发菌株提高11.96%。如图6 c所示,Y-10前期生物量快速提高,pH值随之快速降低,于发酵24 h即降至最低5.26。此外Y-10较出发菌耗糖速度加快。说明Y-10在正常条件下显示出了较强的生长活力。
2.2 5 L发酵罐验证
2.2.1 不控制pH值条件下5 L发酵罐验证
Y-10与出发菌28℃,220 r/min种子培养48 h后按照1.5.3中的5 L发酵罐培养方法,在不控制pH值条件下比较Y-10与出发菌pH值、生物量和纳他霉素产量变化,结果见图7。
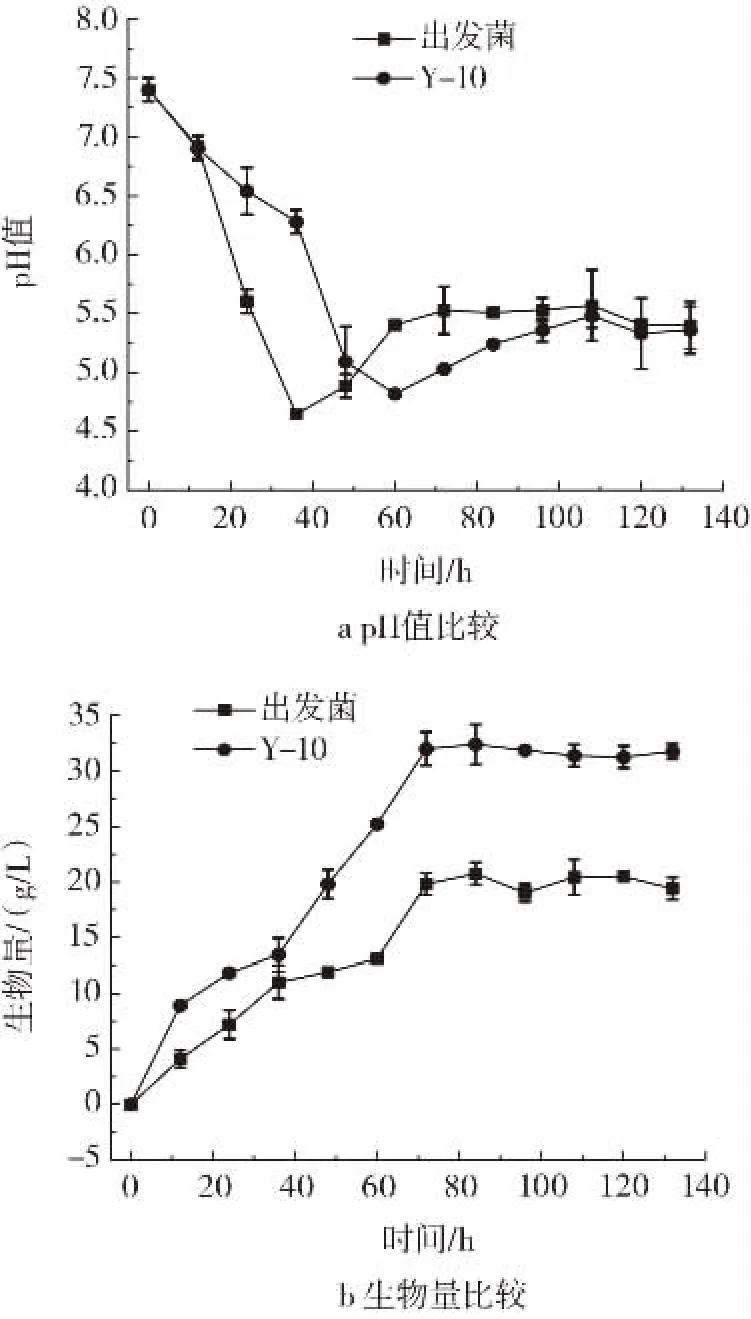
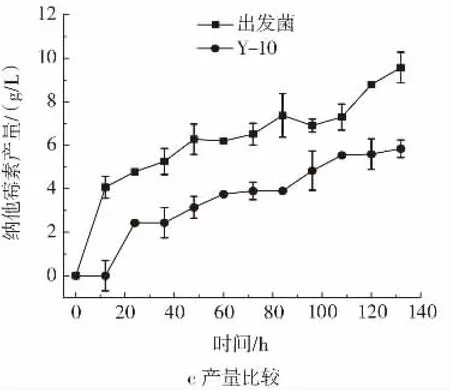
图7 Y-10与出发菌不控制pH值5 L发酵罐验证Fig.7 Verification of 5 L fermenter between Y-10 and the original strain without controlling pH value
如图7a所示,Y-10菌株pH值于发酵60 h自然降低至4.83,而出发菌于发酵36 h自然降低至4.64后开始上升,且整个发酵过程中Y-10整体pH值较出发菌更高,因此判断Y-10可有效减少碱的消耗。如图7b所示,Y-10在pH值自然变化条件下生物量明显提高,生物量最高达到32.40 g/L,较出发菌提高56.22%。但图7c所示纳他霉素产量较出发菌明显降低,纳他霉素最高产量仅为5.58 g/L。Y-10生物量的提高再次证明了其较好的耐酸特性。继续对Y-10控制pH6.3进行5 L发酵罐验证,观察其纳他霉素产量和生物量等的变化。
2.2.2 控制pH6.3条件下5 L发酵罐验证
对菌株Y-10与出发菌在初始pH7.4之后控制pH6.3条件下比较其纳他霉素产量与生物量,并计算其耗碱量,结果见图8。此外,Y-10在发酵过程中菌体异常上浮Y-10与出发菌正常条件下5 L发酵罐96 h取样图见图9。
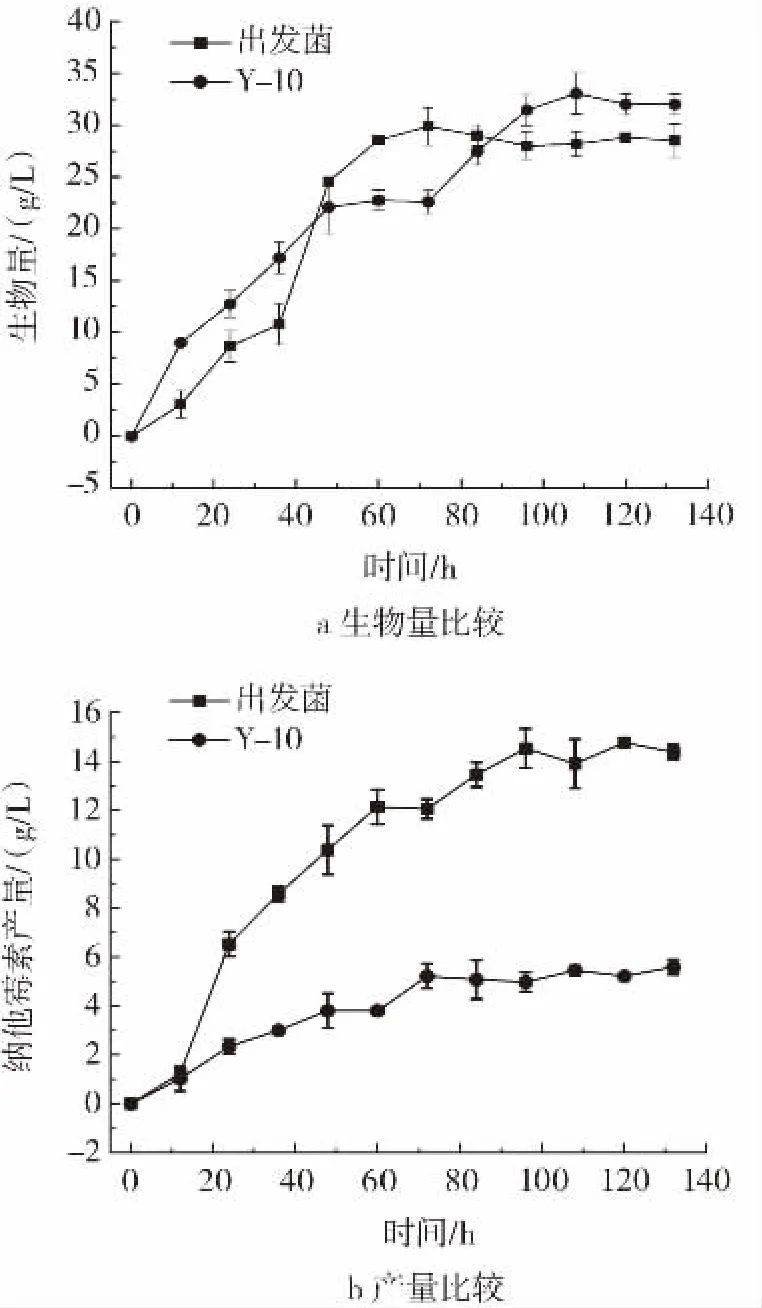
图8 Y-10与出发菌正常条件下5 L发酵罐验证Fig.8 Verification of 5 L fermenter between Y-10 and the original strain under normal conditions

图9 Y-10与出发菌正常条件下5 L发酵罐96 h取样Fig.9 Sample plot of Y-10 and the original strain at 96 h in the 5 L fermenter under normal conditions
如图8 a所示Y-10发酵前期,生物量增长速度较出发菌更快,且到发酵后期生物量最大值较出发菌株提高10.65%。但如图8 b所示,Y-10纳他霉素积累远低于出发菌且与不控制pH值的5 L发酵罐纳他霉素产量接近。原因可能是Y-10在发酵过程中菌体异常上浮(图9),导致菌体与培养基接触面积小,不能充分利用碳源和氮源。在发酵过程中,需要通过流加2 mol/L氢氧化钠维持pH值为6.3。整个发酵过程中,Y-10共耗碱290 mL,相较出发菌少消耗350 mL,说明Y-10可有效减少碱的消耗。
2.3 突变株Y-10的遗传稳定性分析
采用斜面传代方法将Y-10菌株进行连续15代的培养,再分别将活化的各代菌株在28℃摇床内发酵培养120 h,测定其纳他霉素产量及生物量,结果见图10。
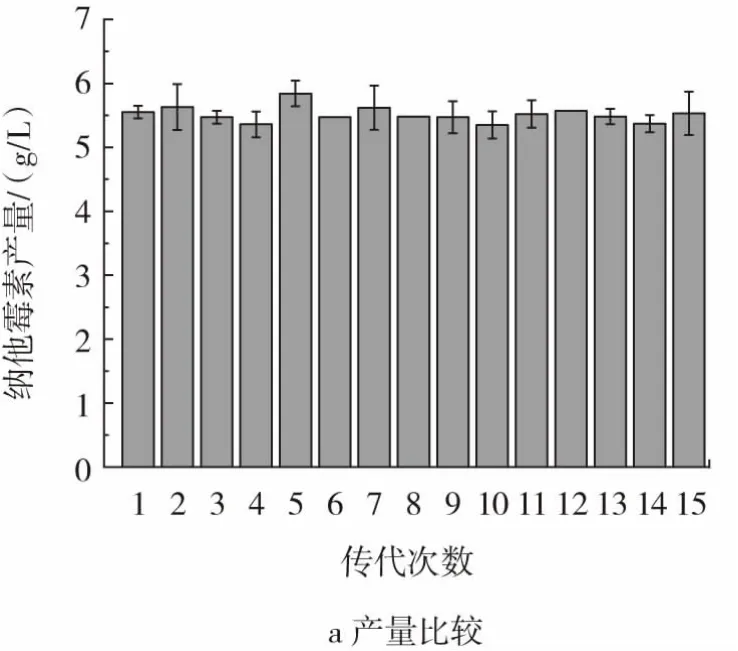
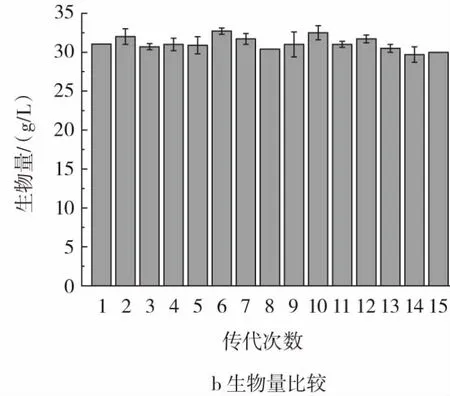
图10 Y-10遗传稳定性验证Fig.10 Verification of genetic stability of Y-10
如图10所示,Y-10纳他霉素产量在正常摇瓶条件下基本稳定在5.50 g/L左右(图10 a),生物量稳定在31.00 g/L左右(图10 b)。说明其生产性能稳定。
2.4 突变株Y-10耐酸机制
2.4.1 发酵过程中Y-10和出发菌胞内ATP比较
胞内ATP含量对于耐酸菌株的耐酸性能有重要影响,对Y-10与出发菌不同发酵时间的胞内ATP含量进行研究,结果见图11。
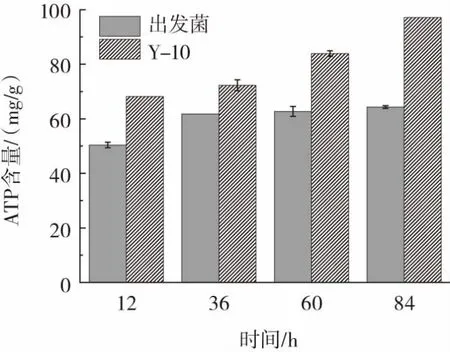
图11 Y-10与出发菌胞内ATP含量比较Fig.11 Comparison of intracellular ATPs between Y-10 and the original strain
如图11所示,出发菌在发酵12 h~36 h,胞内ATP含量明显提高,之后ATP水平趋于恒定。而Y-10在发酵过程中胞内ATP水平不断提高,且始终高于出发菌。较高的胞内ATP水平可以为胞内H+转运提供能量,并维持菌体在酸性环境下的正常生长。
2.4.2 菌株Y-10与出发菌脂肪酸含量比较
对Y-10与出发菌饱和脂肪酸和不饱和脂肪酸相对含量进行研究,结果见图12。
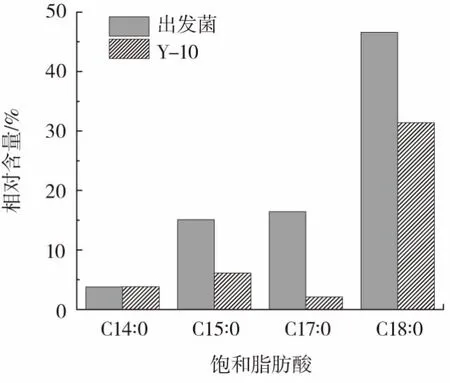
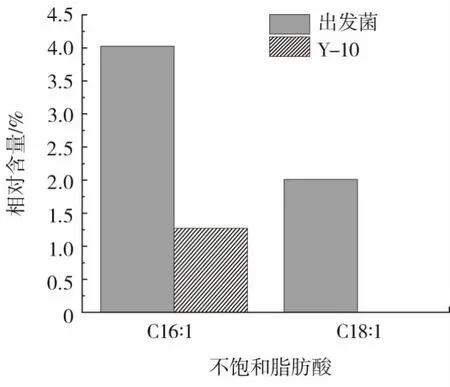
图12 Y-10和出发菌不同脂肪酸相对含量Fig.12 Relative content of different fatty acids in Y-10 and the original strain
图12结果表明Y-10饱和脂肪酸和不饱和脂肪酸的相对含量较出发菌少,且Y-10不饱和脂肪酸比例也比出发菌更低。Y-10细胞膜脂肪酸较低的不饱和度,影响了细胞膜的流动性,进而影响细胞膜物质运输的功能。纳他霉素作为胞外代谢产物,Y-10纳他霉素产量较低可能是受到细胞膜流动性的影响。
3 结论
本试验通过对纳他霉素原始生产菌Streptomyces gilvosporeus TUST01采用ARTP诱变处理,并采用低pH值平板筛选结合24孔深孔板筛选获得耐酸菌株Y-10。Y-10在pH4.0平板上生长良好且在不同的摇瓶发酵条件下,纳他霉素产量和生物量都有明显提高,说明Y-10具有较好的耐酸能力。在不控制pH值和控制pH6.3的正常5 L发酵罐条件下,Y-10都表现出较高的生物量,但纳他霉素产量偏低。较高的生物量同样证明了Y-10具有较好的耐酸能力。对Y-10的耐酸机制进行研究,通过比较Y-10与出发菌的胞内ATP水平发现,Y-10较出发菌株有更高的胞内ATP水平,保证了其在酸性条件下的正常生长。而通过气相色谱-质谱技术研究发现Y-10细胞膜脂肪酸的不饱和度更低,可能影响了Y-10的纳他霉素发酵生产。本文对耐酸菌株筛选及耐酸机制的部分研究可为进一步优化耐酸菌株筛选方法及揭示耐酸机制提供一定的参考。
