日本血吸虫重组蛋白TMEM66P的表达和多克隆抗体的制备
2022-09-23李旭欣GiRi刘静宜刘军涛夏天奇杜鹏飞李慧民程国锋
李 雪,李旭欣,GiRi B R,刘静宜,刘军涛,夏天奇,杜鹏飞,李慧民,李 舜,程国锋
(中国农业科学院上海兽医研究所 农业农村部动物寄生虫学重点实验室,上海 200241)
血吸虫病是由血吸虫尾蚴感染引起的一种人畜共患寄生虫病,对宿主产生严重病理性损害,是WHO发布的六大重点热带病之一[1],也是仅次于疟疾的全球第二大寄生虫病[2]。据估计,全球有超过2.3亿人曾患有血吸虫病[3]。目前,血吸虫病的防控主要依赖化学药物吡喹酮[4],单一药物长期大量使用有可能导致耐药虫株的产生。由于血吸虫的多阶段发育生活史比较复杂,虫体诱发宿主的免疫应答机制以及逃避机制不甚清晰,使获得良好保护效果的血吸虫病疫苗研究被制约。因此,寻找更有效的疫苗抗原候选分子或许成为研发抗血吸虫病疫苗的一种重要途径。
跨膜蛋白位于生物膜,贯穿膜的脂质双层结构,连接膜的内外环境,具有介导信号转导、分子运输、能量转换等多种重要功能[5]。据估计,膜蛋白约在所有编码蛋白中占30%。目前,在药物研发领域约有70%的药物靶点为膜蛋白[6]。血吸虫体被直接与宿主相接触,被视为虫体与宿主信息交流的重要界面。研究表明位于体被的膜蛋白也是抗血吸虫疫苗候选的重要分子。开展体被膜蛋白的功能研究对日本血吸虫疫苗的研发有益。
本文对日本血吸虫TEME66P蛋白进行了初步探索,利用生物信息学分析 TMEM66P,并对其编码cDNA进行了克隆和原核表达,制备了抗血清,并通过Western blot对抗血清的特异性进行了检测。
1 材料与方法
1.1.生物材料 昆明雄性小鼠(8周龄)购自上海杰思捷实验动物有限公司;小鼠通过腹部贴片法感染日本血吸虫尾蚴,在感染后42 d,通过肝门静脉灌注法收集虫体,收集的虫体置于液氮中保存。
1.2 引物合成和DNA测序 引物合成由上海赛默飞生物有限公司合成;重组质粒测序由上海派森诺生物科技股份有限公司完成。
1.3 试剂与耗材 异丙醇购自上海旭庆工贸有限公司;三氯甲烷购自上海科敏生物科技有限公司;HRP标记小鼠抗His标签抗体购自北京百奥莱博科技有限公司;RNAiso Plus购自上海皓嘉科技发展有限公司;感受态细胞均购自上海少辛生物科技有限公司;DNA凝胶回收试剂盒购自擎科生物技术有限公司;一步法快速克隆试剂盒购自翌圣生物科技有限公司;HiScript®Ⅲ RT SuperMix for qPCR(+gDNA wiper)购自南京诺唯赞生物科技有限公司;Omega质粒小提试剂盒购自上海拜尔生物科技有限公司。
1.4 生物信息学分析 通过DNAStar软件查找TMEM66P序列的阅读框,翻译为氨基酸,预测蛋白的大小。利用TMHMM网站(http://www.cbs.dtu.dk/services/TMHMM-2.0/)分析TMEM66P有无跨膜区。在NCBI网站上利用在线工具Blast(http://blast.ncbi.nlm.nih.gov/Blast.cgi)查找TMEM66P在其他物种中的同源序列,使用ClustalW对TMEM66P及其同源序列进行多重序列比对分析[7],使用MEGA 7.0软件,通过邻接法,采用默认参数,构建系统发育树[8-9]。
1.5 重组蛋白的构建、表达与纯化 提取日本血吸虫的总RNA,采用HiScript®Ⅲ RT SuperMix for qPCR(+gDNA wiper)进行逆转录,制备日本血吸虫的cDNA。以此cDNA为模板,根据TMEM66P(GenBank登录号:CAX69776.1)的cDNA序列,使用翊圣无缝克隆引物设计软件(http://122.112.245.84:8080/hieff-clone/)设计包含KpnI和HindⅢ限制性酶切位点的特异性引物,进行PCR扩增(F:5'-GACAGCCCAGATCTGGGTAC CATGTTATTATATTTTGTTTTGA-3';R:5'-TGC TCGAGTGCGGCCGCAAGCTTTTATCTCCGAGAT GTACCACCG-3')。将扩增产物与双酶切的载体通过Hieff Clone®Plus One Step Cloning Kit构建重组质粒,将测序正确的重组质粒转化到DH5α感受态中,置于37℃培养,挑单菌落进行测序验证。将测序正确的菌液利用FastPure® Plasmid Mini Kit提取重组质粒,转化到BL21感受态中,诱导重组蛋白表达。表达产物通过SDS-PAGE凝胶电泳进行分析。将成功表达的重组蛋白进行可溶性鉴定,通过切胶法纯化蛋白。用SDS-PAGE凝胶电泳鉴定纯化效果。用抗His标签抗体作为一抗进行Western blot对重组蛋白进行鉴定。
1.6 抗血清的制备 免疫抗原的制备:将蛋白与Montanide ISA 206 VG佐剂同时放在32℃水浴锅中平衡30 min,然后按体积比例(佐剂∶蛋白=54∶46)在涡悬震荡蛋白时逐滴滴加206佐剂,混合后水平5000 ×g震荡5次,每次1 min,待油相与水相完全融合,25℃条件下静置1 h,观察是否有水油分离及在水相中溶解情况。将免疫原在小鼠颈背部多点注射,50 μg/只,一共免疫3次,每次间隔14 d。在免疫前收集空白血清,采用眼眶静脉丛采血的方法,每只小鼠采集约200 μL。在第3次免疫后第10 d采集血清进行Western blot检测。稀释于1%脱脂牛奶的PBST溶液中室温孵育1 h,PBST洗膜4次,每次10 min。将PVDF膜置于HRP底物ECL发光液中避光孵育3-5 min,用凝胶成像系统拍照。
2 结果
1.7 抗血清的Western blot分析 抗血清对日本血吸虫全虫蛋白中的TMEM66P进行Western blot鉴定。全虫蛋白在10%SDS-PAGE中电泳,将蛋白通过湿转法转移至PVDF膜,在5%脱脂奶粉的PBST溶液室温封闭2 h,一抗为血清,以1∶80稀释于1%脱脂奶粉的PBST溶液中4℃过夜,用PBST洗膜4次,每次10 min。二抗为HRP标记的山羊抗鼠IgG,1∶1500
2.1 生物信息学分析 生物信息学分析表明日本血吸虫TMEM66P蛋白1-148位氨基酸位于膜表面,149-168位氨基酸之间形成一个典型的跨膜螺旋区(图1),序列与曼氏血吸虫同源性为67.79%,与小鼠同源性为44.54%。通过邻接法构建系统发育树,TMEM66P与曼氏血吸虫距离最近,与多房棘球绦虫距离较远(图2)。

图1 TMEM66P的跨膜区预测Fig.1 Prediction of TMEM66P transmembrane region

图2 TMEM66P的系统发育树分析Fig.2 Phylogenetic tree analysis of TMEM66P
2.2 重组蛋白构建、表达、纯化与验证 根据日本血吸虫TMEM66P基因序列设计引物,PCR扩增其CDS序列,并构建pET32a(+)-TMEM66P的原核重组表达质粒。以日本血吸虫成虫的cDNA为模板进行PCR扩增,用1.0%琼脂糖凝胶对扩增产物进行鉴定(图3),结果表明扩增到约1000 bp的DNA片段,与预期大小一致。利用HindⅢ和kpnI限制性内切酶,将扩增产物亚克隆至表达载体pET32a(+),并转化到DH5α中,在含氨苄的培养基中培养过夜,挑取数个单菌落扩大培养,将阳性菌落送测序,进行序列比对。测序结果表明成功构建了重组质粒表达质粒pET32a(+)-TMEM66P。将重组质粒pET32a(+)-TMEM66P转化到BL21感受态细胞中表达,SDS-PAGE分析含有重组质粒的菌体表明rTMEM66P蛋白能在BL21菌中表达(图4A)。经可溶性鉴定发现其为包涵体蛋白(图4B)。通过蛋白切胶法纯化获得重组蛋白,分子量约为52 kDa(图4C)。

图3 TMEM66P编码cDNA的PCR扩增Fig.3 PCR amplification of TMEM66P encoding cDNA

图4 rTEM66P重组蛋白的表达及纯化Fig.4 Expression and purification of rTMEM66P
2.3 抗血清的Western blot分析 Western blot分析表明抗血清能够识别出全虫蛋白中的TMEM66P,大小为38 kDa(与重组蛋白相比缺少Trix-S-his标签),大小与生物信息学预测一致,说明该抗血清具有良好的特异性(图6)。
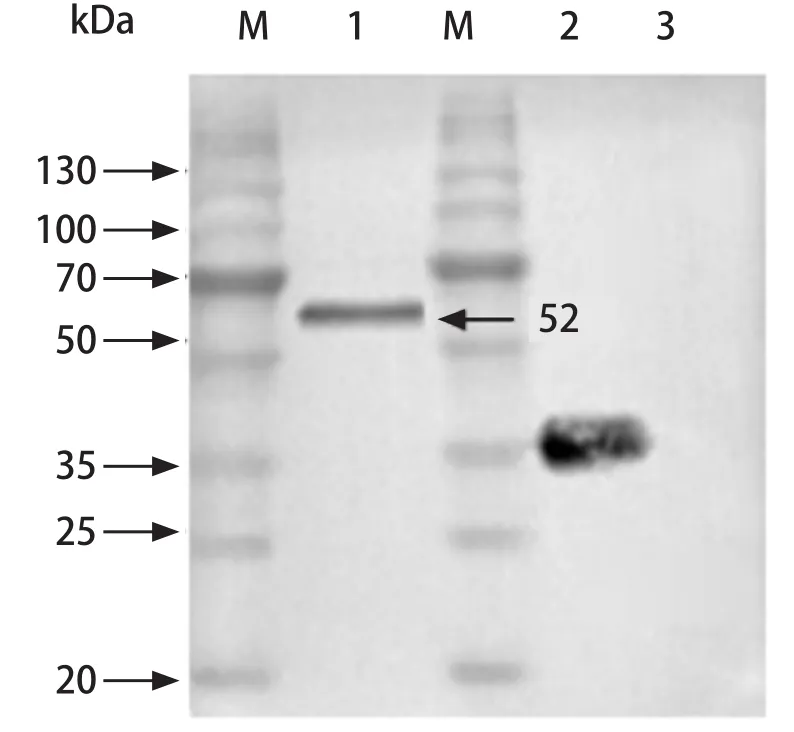
图5 Western blot鉴定重组蛋白Fig.5 Identification of rTMEM66P by Western blot

图6 Western blot鉴定抗血清的特异性Fig.6 Western blot analysis of the specificity of antiserum
3 讨论
跨膜蛋白广泛存在于细胞膜,如内质网、溶酶体、和高尔基体等,对机体健康具有重大意义。研究发现在消化系统中,跨膜蛋白对恶性肿瘤的产生发挥重要作用[10];跨膜蛋白43的编码基因突变会导致肌营养不良和心率失常[11];干扰素诱导跨膜蛋白存在于病毒侵入细胞的主要靶点,如质膜、溶酶体等,能够通过抑制宿主细胞与病毒膜融合来抵抗病毒侵染[12];牛支原体膜蛋白对宿主的细胞和体液免疫应答产生有刺激作用[13];乳脂球膜蛋白能够抑制癌细胞生长、对冠心病自闭症等疾病有重要作用[14];四跨膜蛋白Tspan1、Tspan8、CD82和CD151与肝癌的产生密切相关[15]。许多膜蛋白被作为血吸虫疫苗候选分子进行研究,例如:钙激活中性蛋白激酶、血吸虫副肌球蛋白、谷胱甘肽-S-转移酶、Mr23 kDa膜蛋白、脂肪酸结合蛋白、磷酸丙糖异构酶、日本血吸虫14-3-3信号转导蛋白等[16]。这些膜蛋白能够参与细胞信号传导,是抗血吸虫病疫苗研究中理想的免疫靶标。
抗血清是一种含有多克隆抗体的血清。通过注射抗血清可以传递被动免疫,治疗许多疾病。目前唯一能够治疗埃博拉病毒患者的方法就是通过注射前一个患者的抗血清到体内。抗血清的作用在于其接种注射后直接增加动物体机体内的抗体含量,产生防治效果快而显著,而且在一个地区,往往发病的血清型一致,因而特异性很强,效果显著。
本文对rTMEM66P进行纯化并制备了抗血清,为TMEM66P在日本血吸虫中的功能研究奠定基础。
