不同光质对福鼎大白茶叶片叶绿素荧光参数和呈味氨基酸积累的影响
2022-09-23王加真刘义富曾一霞周玲艳
王加真, 刘义富, 肖 尧, 何 佳, 曾一霞, 周玲艳, 赵 蛾
(1.遵义师范学院生物与农业科技学院,贵州遵义 563000; 2.遵义师范学院茶叶研究所,贵州遵义 563000)
茶叶中游离氨基酸种类和含量的高低不仅影响茶汤的口感和风味,使茶汤具有一定的香气和甜爽味,而且是茶保健功能的重要因子。范培珍等研究发现,谷氨酸(Glu)、甘氨酸(Gly)、丙氨酸(Ala)、脯氨酸(Pro)、茶氨酸(The)是茶叶中的鲜爽因子,丝氨酸(Ser)、Gly、Ala、Pro是甜味类,组氨酸(His)、精氨酸(Arg)、缬氨酸(Val)、蛋氨酸(Met)、异亮氨酸(Iso)、亮氨酸(Leu)、苯丙氨酸(Phe)是苦味类因子,天冬氨酸(Asp)和Glu是鲜味决定因子。张丹丹等通过对白茶的研究认为,不同白茶品种中鲜味氨基酸种类存在差异,在其研究中发现Pro、Gly、Asp和苏氨酸(Thr)对白毫银针贡献甜味,而苦味则受Leu、His和Met影响;在贡眉中贡献甜味的则是Val和Ala代替了Gly和Thr,影响苦味的只有Leu相同,其余的则是Phe、Iso和Val。大量研究一致认为,The占茶叶游离氨基酸含量的50%以上方可调节人体的免疫功能,且对心血管疾病、神经性疾病具有一定的疗效,是茶叶保健功能的重要因子。因此,高氨基酸含量是提高茶叶品质的重要课题之一。方开星等研究218份茶树资源的氨基酸发现,存在高氨基酸(总量≥5.00%)的茶树资源,可选育出高氨基酸含量的茶树品种。通过70% 遮阳处理、春夏季施以一定量尿素、梨茶间植以及冬深耕+重施氮肥+遮阳+喷灌等方法,均能显著提高茶叶氨基酸含量。在茶青加工过程中,采用摊凉萎凋 2~3 h、90~100 ℃烘焙、80~90 ℃提香30 min的工艺可使茶叶中氨基酸含量提高。
育种和栽培措施从材料的层面解决了氨基酸含量的高低,但育种方法需要相应的种质资源、较长的时间和较大的投入,短期难见成效。近年来通过控制光照调节植物生长和生理成为了研究热点。茶树光处理试验显示遮阴可增加氨基酸含量,不同光质照射处理的茶树茶鲜叶中氨基酸含量也不同,红光处理会降低氨基酸含量,蓝光和红蓝光处理使氨基酸含量升高。光质主要是通过影响光合效率而影响氨基酸含量,光合效率可用叶绿素荧光参数来表示。叶绿素荧光参数主要有实际光量子产量[Y (Ⅱ)]、潜在最大相对电子传递效率()、光能的利用效率()和强光耐受能力()等,它们可用来表述植物对光能的利用及耐受效率和能力,并以此来判断植物的抗性、生长和生理状况。叶绿素荧光参数可利用便携式分析仪原位测定,具有快速、无损伤的特点,能够真实反映植物的光合能力,从而能较准确地分析生长环境对植物的影响。本研究以福鼎大白茶一年生幼苗为材料,利用叶绿素荧光测定技术分析其叶片在不同光质下叶绿素荧光参数及叶片呈味氨基酸含量差异,探讨福鼎大白茶叶片对不同光质的利用及其能量分配,以及光质对茶树叶片呈味氨基酸含量的影响,以期为光质调控茶叶功能成分的生产应用提供理论依据。
1 材料与方法
1.1 试验材料
福鼎大白茶由贵州湄潭国家农业科技园区茶树良种繁育中心提供,选取生物量一致的盆栽福鼎大白茶一年生幼苗[株高(20±0.5) cm]进行试验。
光质精准可调人工气候箱(安徽昂科丰光电科技公司);高精度快速光谱辐射计(HAAS-200,杭州远方光电信息股份有限公司);LI-250A光度计(LI-COR,美国);MINI-PAM-Ⅱ脉冲调制式叶绿素荧光仪(德国WALZ公司);200目钢丝网筛;L-3000 型全自动氨基酸分析仪(苏州华美辰仪器设备有限公司)。
荧光灯 T8 L36W/865 O-D 6500K 白光;LED红光灯;LED蓝光灯;LED红蓝混合灯。
1.2 试验设计与方法
1.2.1 光源设置 以荧光灯和LED灯为光源,利用高精度快速光谱辐射计测量光源光谱参数。分别置于白光(荧光灯,对照,W)和红光(645 nm,R)、蓝光(440 nm,B)、红蓝组合光(RB,光质比 1.4 ∶1)下,用LI-250A光度计(LI-COR,美国)采用9点法测定茶苗顶部受光面平均光量子密度,调整茶苗放置高度,将4种光源光照度统一调整为(85±2) μmol/(m·s)。
1.2.2 茶苗栽培设置 试验于2019年12月在贵州省遵义市遵义师范学院茶树光生物学实验中心进行,光周期12 h,温度恒定在(25±1) ℃,相对湿度为(85±3)%。每种光照下设置10盆栽培盆,每盆栽植4株茶苗,每隔3 d浇1次20 mL去离子水。
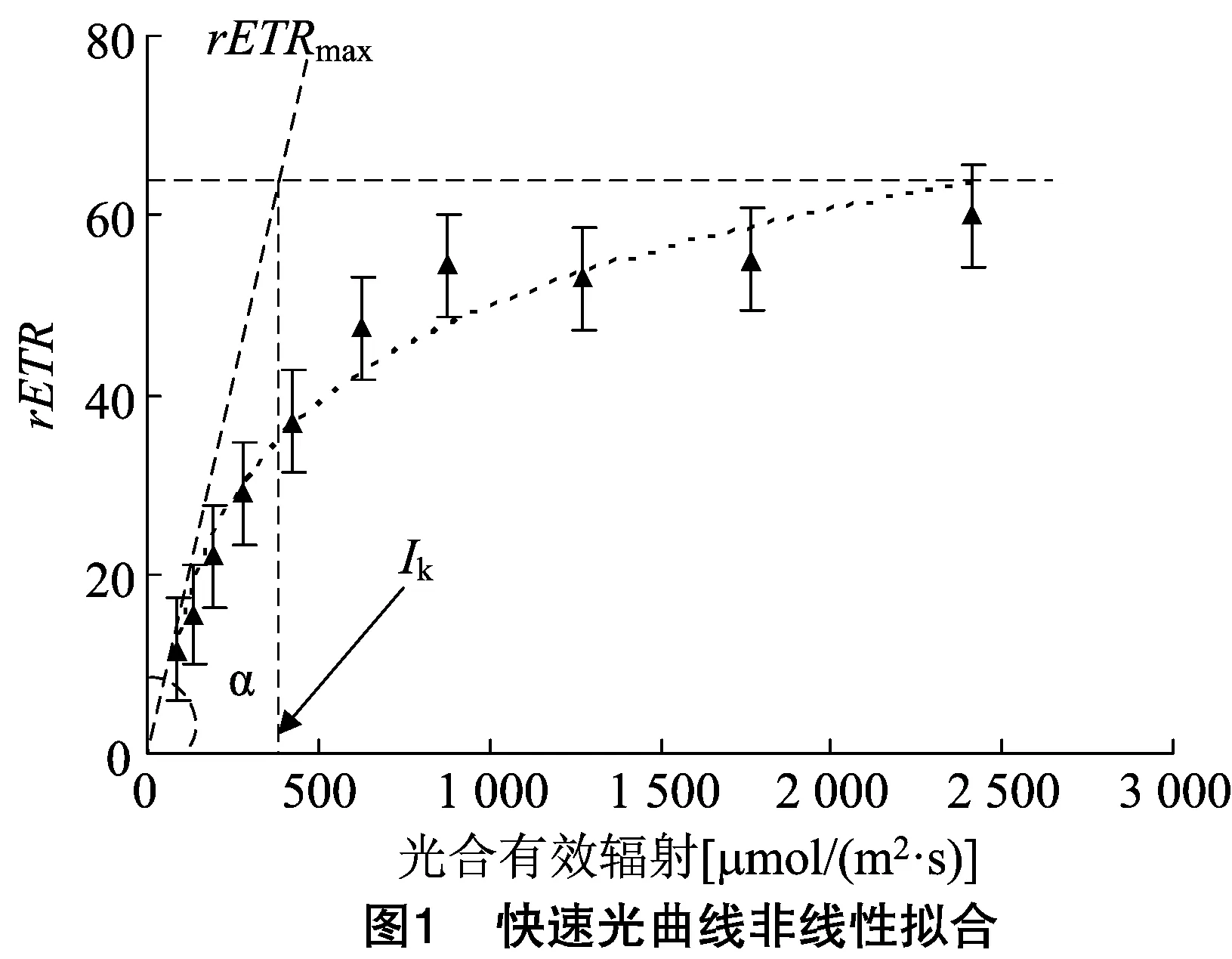
1.2.3 叶绿素荧光仪测定 每株选择3个萌发芽长出的第1张叶。分别在光照0、1、2、3 周对第1张叶暗适应12 h后用MINI-PAM-Ⅱ脉冲调制式叶绿素荧光仪测定光系统Ⅱ快速光曲线,对光曲线进行非线性曲线拟合后,曲线初始斜率为光能利用效率(),曲线的最大值为潜在最大相对电子传递效率(),表示方法如图1所示。同时测定第1张叶在光适应下的实际光化学效率(YⅡ)和耐受强光的能力()。每株3张叶的平均值作为该株的参数。
1.2.4 叶片氨基酸含量测定 于光照处理3周后,每处理摘取测叶绿素荧光参数的对应第1张片,用100 ℃水蒸汽杀青3 min,80 ℃下烘干至恒质量,粉碎过200目筛后测氨基酸含量。18种氨基酸组分含量的测定采用氨基酸分析仪。
1.3 数据统计分析
测定数据利用Excel 2013进行平均值、标准差等的分析及排列,然后利用SPSS 18.0 软件进行单因素方差分析。
2 结果与分析
2.1 光源光谱特征分析
试验用光源经HAAS-200测量分析获得如表1所示的光谱参数。
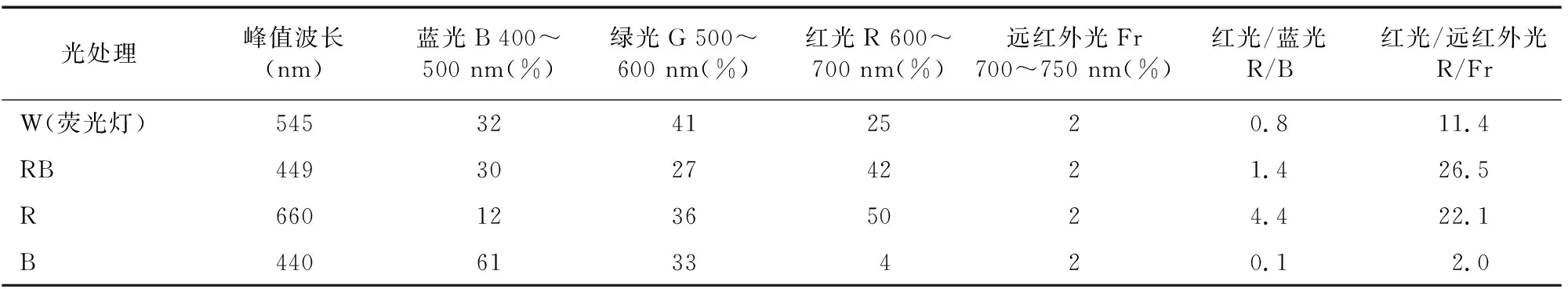
表1 试验用不同光质的光谱特性
4种光源W、RB、R、B的主波长分别为545、449、660、440 nm,它们的红蓝光质比分别为0.8、1.4、4.4和0.1。W光源与RB光源相比较,有较高的绿光比例(41%),峰值波长位于黄绿光区域。根据前人研究,红光(610~720 nm)和蓝光(400~520 nm)是植物敏感光源,与光合作用效率相关性最显著,红光下植物生长旺盛,蓝光次之。茶树为喜阴植物,本试验用R、B、RB和W分别为600~700、400~500、500~600 nm,处于合理的区间内,可满足茶树对不同光质反应研究的需要。
2.2 光质对茶树叶片叶绿素荧光参数的影响
如图2所示,与处理前相比,Y(Ⅱ)在处理1周后,出现下降趋势,但差异不显著;处理2周后,白光和红蓝光处理下与处理前相比Y(Ⅱ)下降显著,蓝光处理与1周后相比显著升高,红光处理下不变;处理3周后除红蓝光外其他光照下均恢复到处理前水平。光质之间比较,处理1周后,红光处理显著低于白光,与其他2种光之间无显著差异。处理2周后,蓝光处理Y(Ⅱ)显著高于红光和红蓝光处理。处理3周后,红蓝光处理明显低于其他光处理。PSⅡ反应中心部分关闭时,捕光色素分子实际原初光能捕获效率即为Y(Ⅱ)[Y(Ⅱ)=(-)/],可快速地确定某种环境条件下 PSⅡ反应中心的作用效率。所以,红光具有提高PSⅡ反应中心作用效率的作用,增强捕光色素分子的光能利用率。
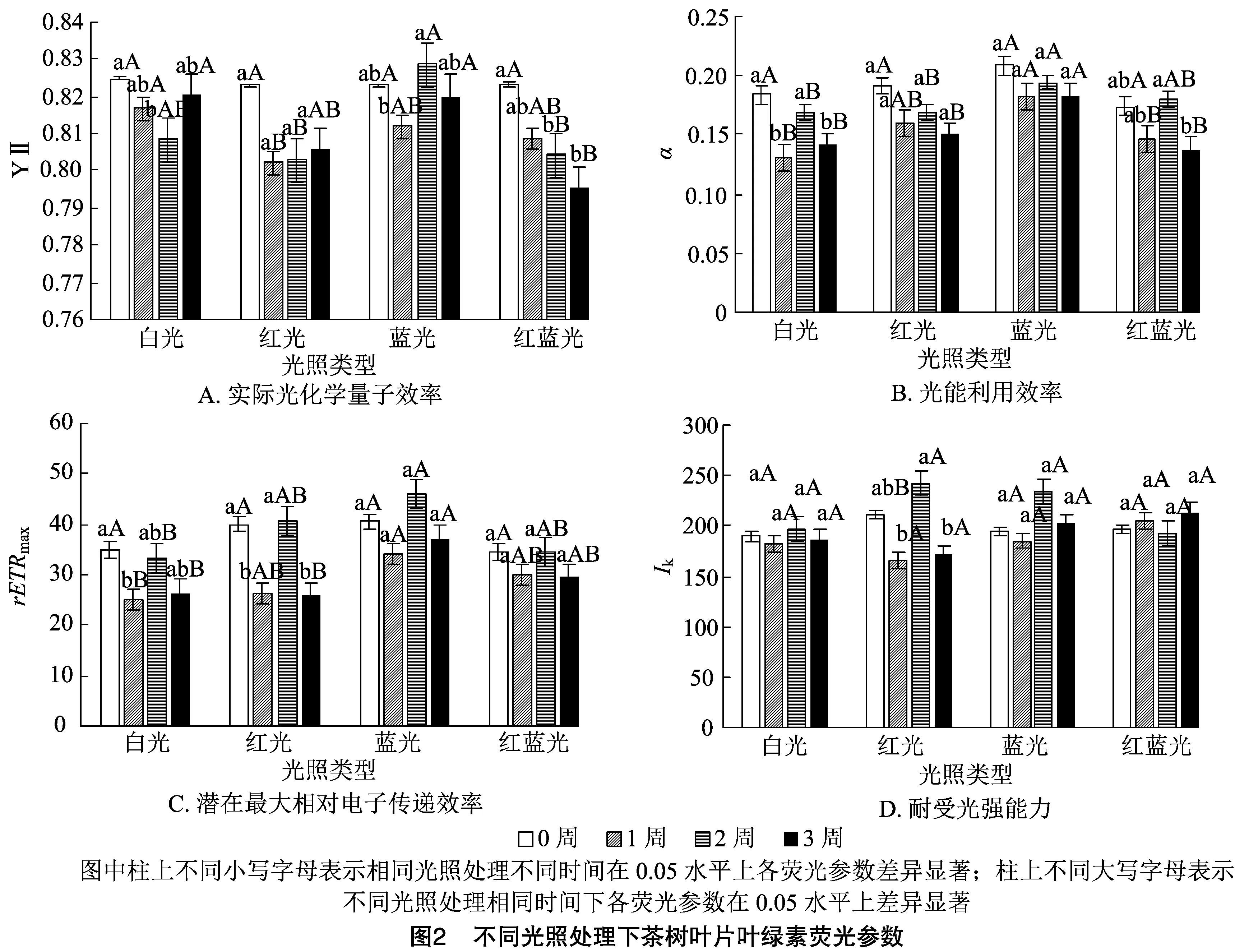
在处理1周后,均表现下降的趋势,但只有在白光处理下与处理前存在显著差异,且蓝光处理下显著高于白光和红蓝光处理(<0.05)。处理2周后,值均开始回升,但仍然只有白光处理下值与处理1周结果存在显著差异;蓝光处理显著高于白光与红光处理。处理3周后,白光和红蓝光处理下值又出现下降,显著低于处理2周后的结果;不同光照之间表现为蓝光处理下显著高于其他光照处理。这说明红光和蓝光照射下,较稳定,且蓝光处理效果较红光好。曲线初始斜率反映光化学反应的启动速率,是反映光合器官对光能利用效率的参数。红光和蓝光处理有稳定茶树光化学反应启动速率的作用。
反映叶片光合作用的潜力。在不同光照处理1周后,白光和红光处理下显著下降,且红光处理下降幅度最大,显著低于其他光照处理。处理2周后,均有上升趋势,但只有红光处理下显著高于处理1周的结果,且红光、蓝光、红蓝光处理下明显高于白光处理。处理3周后,均有下降趋势,但仍然只有红光处理显著低于处理2周的结果;不同光质间比较表现为蓝光处理显著高于白光和红光处理。以上结果表明,红光会降低茶树叶的光合潜力,而B对叶片光合潜力没有影响。
是初始斜率线和水平线的交点的横坐标值,反映样品对强光的耐受能力。相对稳定,只有红光处理2周后比处理1周显著升高,然后到3周又显著降低,说明光质对的影响不大。
2.3 光质对茶树叶片呈味氨基酸含量的影响
18种氨基酸含量在光照处理21 d后进行测定,结果如表2所示。18种氨基酸中只有Pro含量不受光质的影响,酪氨酸(Tyr)、氨基丁酸(-Butyr)、Gly、Phe、Arg在红光处理下含量最高。白光处理下-Butyr、Asp、Ser、Ala、Met、Phe、Arg、His含量均最低,表明红光、蓝光、红蓝混合光处理对大多数氨基酸都有较大的影响,有提高的功效。与白光相比,红光可显著提高-Butyr、Ala、Met、Ser、Phe、Tyr、Lys和Arg的含量,降低Asp、The、Glu和Leu的含量。蓝光可提高-Butyr、Asp、The、Glu、Phe含量,降低Gly、Val含量;红蓝混合光可显著提高Asp、The、Glu、Ala、Met、Thr、Iso、Val、Leu、Lys、Arg和His含量。
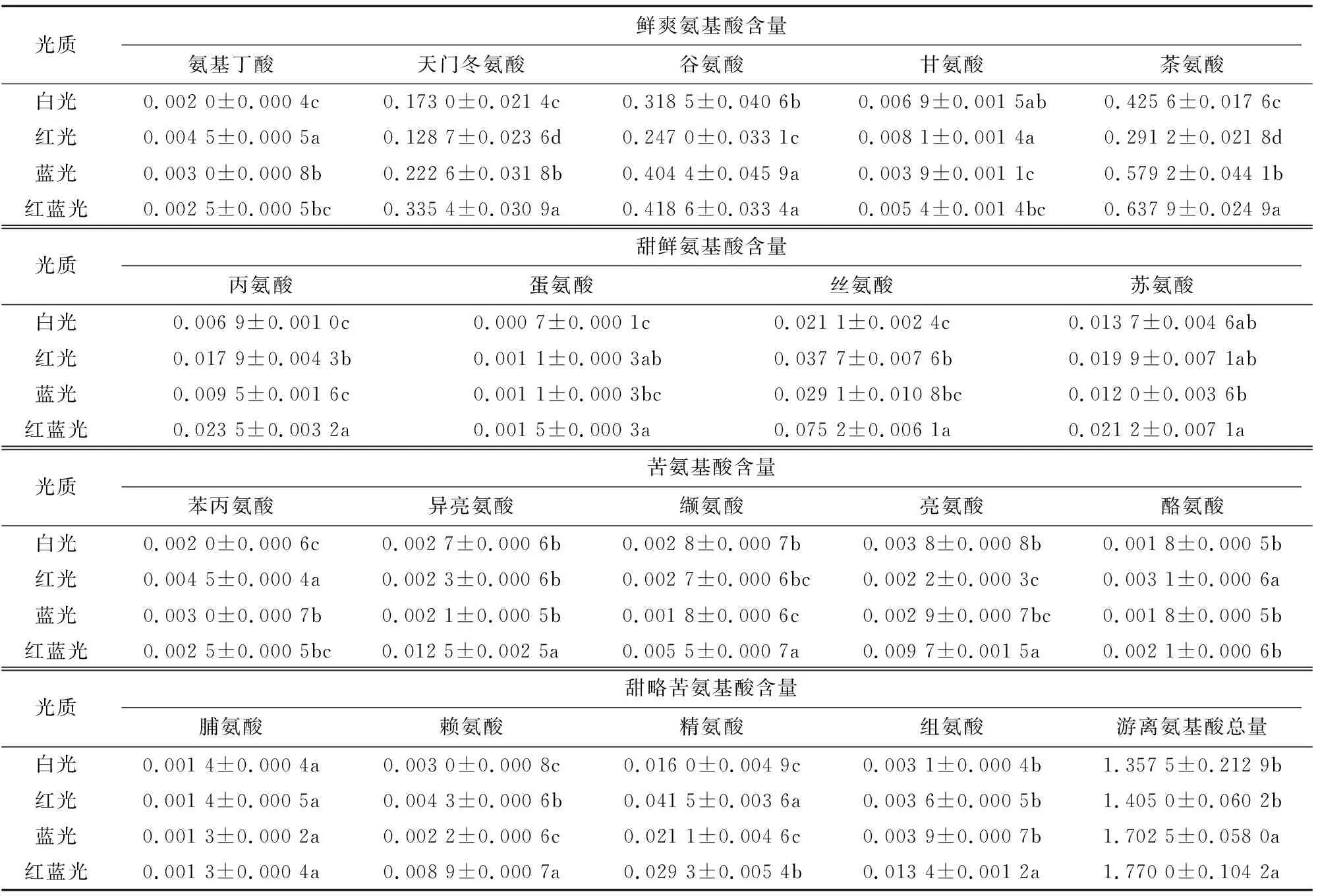
表2 不同光质处理21 d后对福鼎大白茶叶片18种游离氨基酸组分含量的影响 %
不同颜色光照处理下茶叶氨基酸含量之间相比较,红光处理下-Butyr、Gly、Phe、Tyr和Arg含量比蓝光和红蓝混合光处理高,Lys和Ala含量比蓝光高而比红蓝混合光处理低。蓝光处理下Asp、Glu和The含量比红光处理高,Asp、The、Ala、Met、Ser、Thr、Iso、Val、Leu、Lys、Arg和His含量比红蓝混合光处理低。红蓝混合光处理下,Asp、Glu、The、Ala、Met、Ser、Thr、Iso、Val、Leu、Lys和His含量均是最高。
以上结果表明,增加蓝光比例,有助于主要氨基酸(图3,Asp、Glu、The占游离氨基酸比例高于70%,因此定义这3种氨基酸为主要氨基酸)的积累,而红光则主要有助于带苦味的氨基酸的积累。所以,可通过增加蓝光的比例,提高茶树甜爽氨基酸含量,从而提高茶叶的鲜爽滋味。蓝光和红蓝混合光处理下氨基酸总量的提高主要是通过提高天门冬氨基酸、Glu和The这3种主要氨基酸的含量。

2.4 茶树氨基酸含量与叶绿素荧光参数的相关性分析
表3表明,Ala、Lys、Ser、His含量与Y(Ⅱ)呈显著或极显著负相关,Val含量与呈显著负相关,Glu与呈显著正相关,Asp、Glu、Pro、Met、Thr、Val、Leu含量与呈显著或极显著正相关。叶绿素荧光参数可利用便携式仪器原位、快速测定,根据氨基酸含量与叶绿素荧光参数的相关性可间接判断氨基酸含量的高低,对茶树高氨基酸育种具有重要的参考价值。
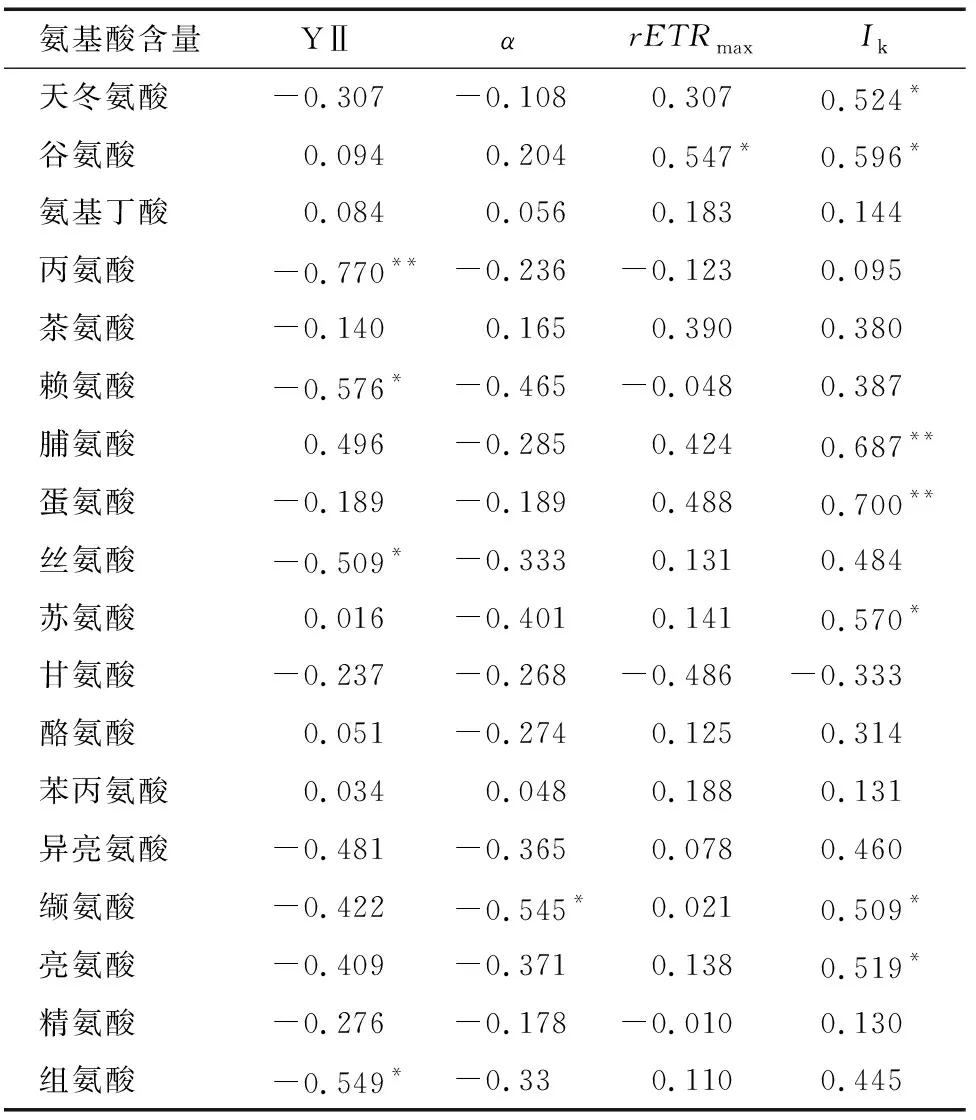
表3 氨基酸含量与叶绿素荧光参数的相关性
3 讨论与结论
绿色植物含多种光受体,如光敏色素、隐花色素、向光素等。不同的光受体对不同色光的敏感不同,如光敏色素对红光敏感,隐花色素对蓝光敏感。因此不同色光对植物生长发育的影响不同。蓝光对脱黄化、开花、昼夜节律、基因表达、向光性、叶绿体运动和气孔运动等都有一定的调节作用,并且,蓝光能增加茶树氨基酸总量。本研究中,蓝光和红蓝混合光处理均增加了氨基酸总量,且红蓝混合光效果更好。并且,蓝光和红蓝混合光主要是通过增加天冬氨酸、谷氨酸和茶氨酸等主要氨基酸的含量而增加氨基酸总量,而对部分苦味氨基酸含量无影响或有降低的作用。所以在茶树栽培过程中,适当增加蓝光的比例,有助于提高茶叶的品质。
红光通过抑制植物节间伸长而抑制植物茎的生长,因此,红光处理有利于控制茶树的株高。红光处理对植物内含物也有较大影响,处理叶片的混合光中升高红光的比例可降低叶片总氮含量,包括叶绿素和蛋白质含量,增加包括还原性糖含量的总碳含量。因此,提高红光的比例,可改善茶叶的茶汤口感,且在施氮水平较高的条件下,还可调节碳氮代谢平衡。水稻试验表明,红光处理5 d的幼苗各种氨基酸含量都远远低于蓝光和白光处理的幼苗。在本研究中,红光处理下,氨基酸总量比蓝光和红蓝光混合光处理的低,与其他植物的研究结果较一致,而本研究中氨基酸总量降低的原因主要是天冬氨酸、谷氨酸和茶氨酸的含量显著降低造成的。
光质对茶树生理的影响主要通过调控光合作用来实现,叶绿素荧光参数则是茶树光合作用过程变化的反映。叶绿素荧光参数中 Y(Ⅱ)反映叶片吸收的光能中用于光合电子传递的能量占比高低,高 Y(Ⅱ)往往代表着高光合效率,包括高效的光子吸收和电子传递,用于暗反应中碳同化的能量,PSⅡ功能降低,Y(Ⅱ)也随之下降。Ala、Lys、Ser和His与Y(Ⅱ)呈显著或极显著负相关。试验中蓝光照射2周后Y(Ⅱ)有上升的趋势,红光照射使 Y(Ⅱ) 下降,所以红光处理Ala和Lys含量升高和蓝光处理这2种氨基酸含量降低的原因可能是外源光质影响Y(Ⅱ)的结果。
是反映植物对光强度耐受程度的参数,本试验中红光处理降低了茶树对强光耐受的能力。相关分析显示,天冬氨酸、谷氨酸含量与呈显著正相关。研究表明,红光可能通过降低而影响了天冬氨酸和谷氨酸的积累,蓝光和红蓝混合光则相反。反映植物捕光能力,是叶片光合作用最大能力的表现。在红光处理下明显降低,与氨基酸含量的相关性研究显示,与Glu含量呈显著正相关,可推测,红光通过降低而降低Glu的含量。反映光能利用效率,蓝光处理下值明显高于其他光照处理,而Val含量与呈显著负相关,可见,蓝光可能通过使升高而降低Val含量。
