柚木野螟成虫复眼外部形态和超微结构观察
2022-09-17苑鹏宇胡可炎钱沉鱼王胜坤温秀军
苑鹏宇, 胡可炎, 钱沉鱼, 王胜坤, 温秀军, 王 偲, 马 涛,*
(1. 华南农业大学林学与风景园林学院, 广州 510642; 2. 中国林业科学研究院热带林业研究所, 广州 510520)
昆虫复眼(compound eye)是探测和识别外部环境的主要视觉器官,其形态学和内部结构特征决定了视觉功能特性,在昆虫觅食、求偶、搜寻产卵场所、归巢和避敌等行为中发挥着重要作用,进而影响其识别形状和颜色以及检测周围环境中的特定对象的能力(Rutowskietal., 2002; Warrantetal., 2010)。研究复眼的形态和结构特征为探讨昆虫对明暗环境的适应调控奠定组织学和生理学基础,也为理解其光视觉行为与相关生理反应提供重要依据。

柚木野螟Eutectonamachaeralis属鳞翅目草螟科,是危害珍贵树种柚木Tectonagrandis的主要食叶害虫(吴士雄等, 1979; Lanetal., 2020)。幼虫取食叶肉留下叶脉,呈疏网状,严重时林相如火烧;成虫白天隐藏在林内地被物或杂草中,夜间集中树冠处活动。该虫发生量大、世代周期短、为害期长,并且虫口高峰期正值柚木生长季节,严重影响植物光合作用,致使柚木生长减慢,对阔叶树生态系统产生严重威胁。目前相关研究多侧重于柚木野螟生物学特性和生物化学方面(张蒙等, 2014; 蓝来娇等, 2018, 2019, 2020),关于其复眼结构特征、视觉行为及其光适应机制等尚未研究报道。
本研究利用光学和电子显微技术观察柚木野螟复眼的形态、组织结构和超微结构,旨在阐明其复眼形态和结构特征,为更好地了解柚木野螟复杂的视觉行为和感光机制的关系奠定基础,以期为发展柚木野螟靶标精准、环境友好的绿色防控技术提供理论依据。
1 材料与方法
1.1 供试虫源
柚木野螟成虫和幼虫均采于广东省惠州市龙源兴茂生态园(22.50°N, 114.30°E),人工气候箱中饲养,饲养温度为26±1℃,相对湿度为75%~80%,光周期为16L∶8D。
1.2 扫描电子显微镜样品制备及观察
解剖观察前,取柚木野螟雌雄成虫各10头,冷冻麻醉处理10 min。断头后,置于2%多聚甲醛和2.5%戊二醛混合固定液(10%多聚甲醛∶25%戊二醛∶0.2 mol/L磷酸缓冲溶液∶蒸馏水=2∶1∶5∶2, v/v)(pH 7.2)中4℃固定24 h。0.1 mol/L磷酸缓冲液(pH 7.2)清洗样品3次,每次10 min。30%, 50%, 70%, 80%和90%乙醇梯度脱水各15 min,再用100%乙醇脱水3次,每次15 min。脱水后,用乙酸异戊酯置换2次,各30 min。之后,经过CO2临界点干燥,用双面导电胶将样品固定到样品台上,放入自动精细离子溅射仪(JFC-1600)内喷金2次,每次1 min。置于扫描电子显微镜(JEOL, JSM-6510LV)下观察并拍照。
1.3 透射电子显微镜样品制备及观察
取柚木野螟雌雄成虫各10头,断头后,在光学显微镜下剖获完整复眼。将复眼放入2%多聚甲醛+4%戊二醇混合液中,经醋酸双氧铀块染色后4℃下避光过夜。固定后的复眼样品经缓冲液漂洗4次,每次20 min;再用锇酸固定液避光固定4 h,固定毕用双氧水漂洗多次。30%, 50%, 70%, 85%和95%乙醇溶液逐级脱水,每次15 min,再用100%的乙醇脱水2次,每次20 min。随后,用3个不同体积比例的丙酮和树脂混合物(丙酮∶树脂=3∶1, 1∶1和1∶3, v/v)依次渗透,再用纯树脂渗透两次,每次12 h。渗透好的样品用纯树脂包埋,将包埋好的样品置于30℃烘箱中聚合12 h,再在60℃下聚合48 h。
聚合完成后,在Leica EM UC7冷冻超薄切片机上,将样品块分别用玻璃刀、钻石刀进行半薄切片(约2 μm)和超薄切片(约70 nm)。半薄切片置于带水滴的载玻片上,烘干后用1%甲苯胺蓝染液染色,用蒸馏水冲洗掉染料,在连接CCD成像系统的Olympus BX-51型光学显微镜下观察并拍照。超薄切片依次用2%醋酸双氧铀染色8 min和4%柠檬酸铅染色10 min,在FEI/Talos L 120C透射电子显微镜下观察并拍照。
1.4 数据分析
上述所拍摄照片均使用Adobe Photoshop CC 2018添加比例尺和标注,应用ImageJ 1.8.0软件测量,每个指标至少观测5个样品。数据采用SPSS 16.0软件进行统计,其中利用独立样本t检验对扫描电子显微镜图中柚木野螟雌、雄成虫的小眼周长、直径等测量数据进行分析。
2 结果
2.1 柚木野螟成虫复眼外部形态
柚木野螟成虫复眼着生于头部触角基部,属对称性复眼。从虫体头部的背面观可以看出,复眼位于头部两侧,左右复眼外部形态基本一致,呈椭球形,有光泽,长轴约823.69 μm,短轴约386.15 μm(图1: A, B)。小眼面微外凸,排列整齐紧密有规律,各小眼间平均间隔0.18 μm,复眼中心小眼呈正六边形,边缘偶有五边形(图1: C, D),外表面密布着角膜乳突(图1: E)。柚木野螟雌、雄成虫复眼总面积相差不大,但在小眼数量、大小及边长等方面上表现出差异,雌虫小眼面积略小于雄虫,雌、雄成虫小眼数量分别为2 300~2 755个和1 950~2 316个(表1),小眼间偶尔可见感觉毛,长约8.50~9.75 μm(图1: F)。

表1 柚木野螟雌雄成虫复眼外部形态结构的参数比较

图1 扫描电子显微镜下柚木野螟成虫视觉器官外部形态
2.2 柚木野螟成虫复眼组织结构
柚木野螟成虫单个小眼平均长度为193.40±1.85 μm,从外到内为角膜、晶锥、视杆、初级色素细胞(primary pigment cell)、次级色素细胞(secondary pigment cell)和基膜(图2: A)。综合纵切和不同层深的横切可见,柚木野螟成虫复眼中心区晶锥多层紧密排布,晶锥下方无细胞结构和色素颗粒,呈空腔状;边缘区角膜下方的晶锥呈子弹状,与视网膜细胞之间存在明显的透明区,长约29.92±4.75 μm,主要为次级色素细胞,属重叠像眼(图2: B, C)。
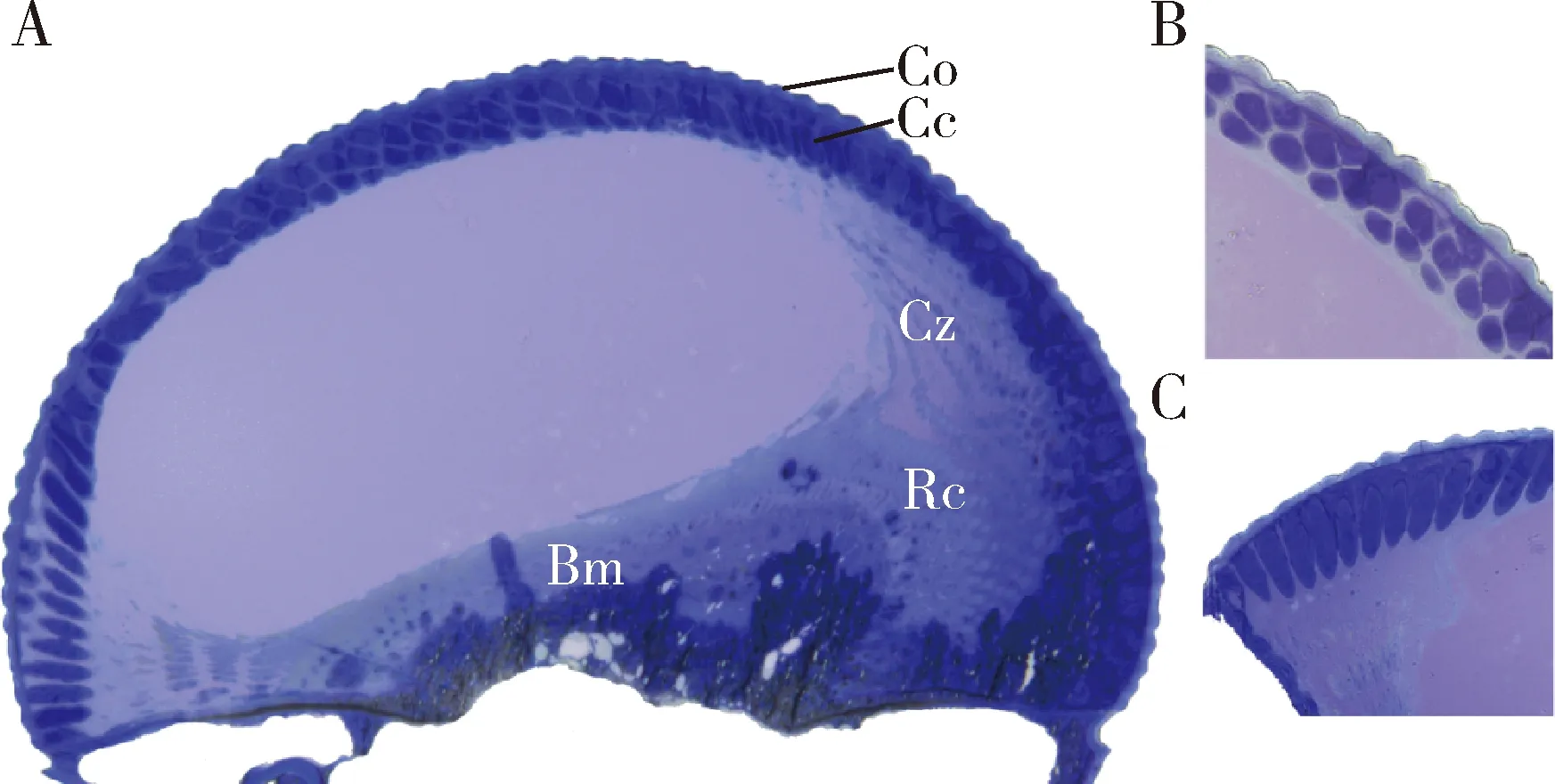
图2 柚木野螟雌成虫复眼组织结构光学显微镜观察
2.3 柚木野螟成虫复眼组织超微结构
柚木野螟雌、雄成虫小眼的形态和结构均无明显差异。因此,为方便描述,下文统一选择雌蛾的小眼从外到内的结构进行描述。
小眼的屈光器由角膜和晶锥构成,角膜位于最外层,宽18.34±1.21 μm,外曲面半径为15.45±0.73 μm,最厚处约为9.03±0.33 μm,主要保护复眼免受机械损伤。角膜外密集分布微小凸起,即角膜乳突(corneal nipples),以减少光线反射并增加感受能力,直径约180±10 nm,高约270±30 nm。角膜纵切面为透明层状结构,呈凸透镜状,约有25个纤维层,内外层排列紧密,中间排列较疏松,各层染色深浅不一,表明其质地不均匀(图3: A)。
角膜下方的晶锥被一对初级色素细胞包围,纵切面呈上宽下窄的子弹状,长约28.54±1.4 μm,最宽处约15.67±1.24 μm,与角膜一同构成屈光器(图3: B)。晶锥横切面近卵圆形,由4个等分的晶锥细胞(crystalline cone cell)及其细胞分泌物组成,染色强度由内到外逐步变亮(图3: C)。晶锥细胞细胞核位于晶锥最远端(图3: D),细胞质从核区沿中央分泌区外围向下延伸(图3: B),并且分布大量平行于晶锥轴的微管。
一对初级色素细胞与角膜相接,一直围绕晶锥(图3: A),细胞核卵圆形,位于晶锥的中部水平(图4: C),细胞质中分布许多直径为0.77±0.09 μm的球状色素颗粒,近端有许多电子致密粒子(图3: A; 图4: D),远端含有线粒体和光面内质网,以分泌形成新的角膜生成物(图3: D)。
6个次级色素细胞占据相邻小眼之间的空间,从上至下依次围绕初级色素细胞、视网膜细胞直至基膜,也被称为膜间色素细胞(图3: A; 图4: C)。 次级色素细胞的细胞核分布于晶锥的远端(图4: A),细胞质多呈中性或弱电子密度,形成一个“透明区”,宽度为25.74~38.60 μm,含大量囊泡,将小眼的屈光器和感光层分开,分布许多直径为0.55±0.05 μm的球状色素颗粒(图4: B),明显小于初级色素细胞中色素颗粒大小(表2),并且电子密度分布不一。
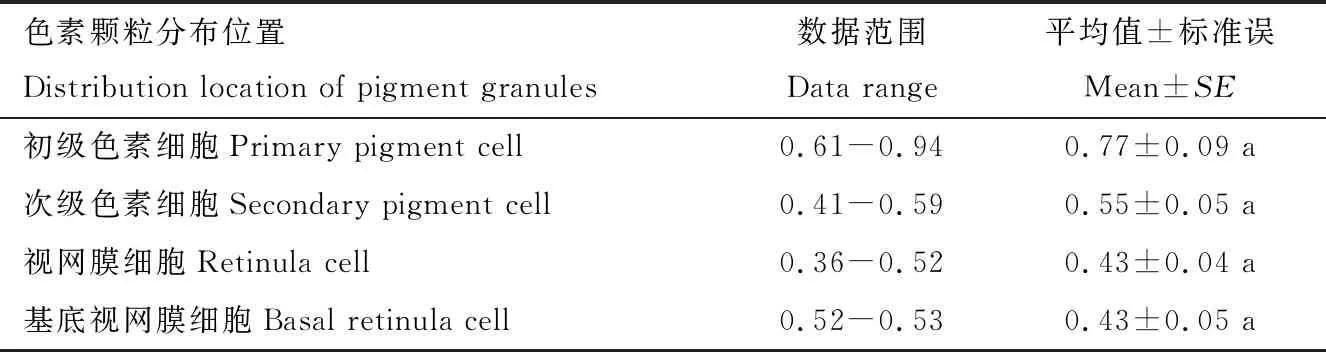
表2 柚木野螟成虫复眼内色素颗粒直径 (μm)

图3 柚木野螟雌成虫小眼远心端透射电子显微镜观察

图4 柚木野螟雌成虫小眼远心端透射电子显微镜观察
透明区将视杆层与屈光器分隔,偶尔视杆远端延伸至透明区。晶锥下方,沿小眼纵轴11个视网膜细胞内侧细胞膜特化形成细丝状微绒毛,这些微绒毛整齐紧凑地排列在一起组成视小杆,呈放射状排列,组合呈融合型视杆(图5: A)。另一个视网膜细胞可见于近基膜端,故称为基底视网膜细胞。中央融合的视杆,全长约125.98±5.18 μm,从晶锥的下方延伸至基底,远端视杆的视网膜细胞数目变化为±1,视杆通过锥形微气管绒毡层彼此隔离,与次级色素细胞不同,相邻小眼之间不共用微气管,因此小眼间可见双层微气管(图5: B)。透明区消失于视杆远端和近端交界处。次级色素细胞填充相邻视网膜细胞之间的空间,但在视杆近端,该细胞逐渐变细,接近基底时末端膨大(图5: B, D)。视网膜细胞色素颗粒多集中于基膜近端,直径约0.43±0.05 μm的色素颗粒,且该区域围绕视网膜细胞的微气管明显变大,并呈现出更加复杂嵴结构(图5: C)。基底视网膜细胞被其余11个视网膜细胞的轴突完全包裹,细胞核几乎充满整个细胞质空间,并且细胞质中分布线粒体和高尔基体等细胞器(图5: D)。

图5 柚木野螟雌成虫小眼视网膜细胞近心端横切透射电子显微镜观察
小眼最底部为栅栏状的基底膜,厚度约1.03±0.27 μm,是一种高度特化的规则细胞外基质变体,排列在上皮细胞的底部上,将眼部感光层与神经层分隔(Fischeretal., 2012)。基底膜上方,基底视网膜细胞变成以拥有线粒体和内质网为特征的轴突,同其他视网膜细胞和次级色素细胞的轴突一起穿过基底膜上小孔进入视叶(图6: A, B)。一些膜下色素细胞参与基底膜的形成,可看到其细胞核和色素颗粒(图6: C)。基膜上的小孔位于每个小眼的基底中央,并和基膜下方气管细胞产生的膜间间隙,是视网膜细胞轴突向内神经投射的开口(图6: D)。
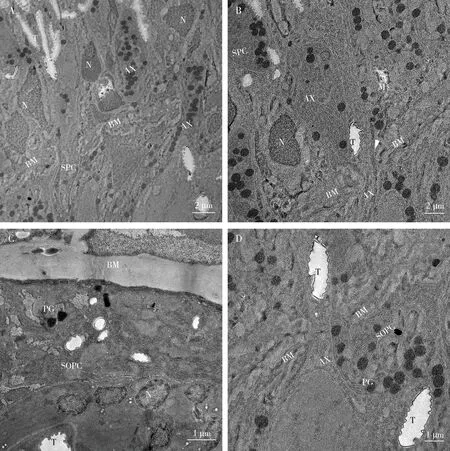
图6 柚木野螟雌成虫小眼基膜透射电子显微镜观察
3 讨论
柚木野螟复眼呈椭球形,属对称性复眼,占据头部大部分空间,视野较大而小眼间角较小,对目标物体的敏锐度高。其雌、雄成虫复眼在小眼数量和大小上表现出差异: 雌性成虫复眼有2 300~2 755个小眼,雄性成虫有1 950~2 316个小眼,并且雌虫小眼面积略小于雄虫的。昆虫视觉能力受复眼大小、空间位置及小眼数量等诸多因素的影响(Land, 1997; 张胜杰等, 2015),一般情况下复眼中小眼数量越多,面积越大,成像越清晰(文超等, 2019)。由此推测,柚木野螟雌性成虫的复眼分辨能力稍强于雄性,更利于定位寄主或产卵场所等。这种复眼性二态的现象也表现在其他昆虫中,像二点委夜蛾Athetislepigone和Operophterabrumata雄蛾小眼数均多于雌蛾,雄蛾有更发达的视力寻找雌蛾交配(Meyer-Rochowetal., 2008; 杨心月, 2015);鞘翅目(Coleoptera)大场雌光萤Rhagophthalmusohbai雌、雄成虫小眼数量差异极大,雄虫小眼个数约3 000个,而雌性小眼个数仅35个(Lauetal., 2007);膜翅目(Hymenoptera)红火蚁Solenopsisinvicta复眼的形状、小眼数目和形状、表面被物随性别和品级不同均有较大差异,生殖蚁显示出更为发达的视觉能力(范凡, 2008)。

表3 柚木野螟成虫复眼内部形态结构
柚木野螟复眼的小眼角膜表面被有无数微小的角膜乳突,Bernhard等(1970)根据角膜乳突高度进行了分类:Ⅰ类(高度低于50 nm),Ⅱ类(高度50-200 nm)、Ⅲ类(高度约250 nm)。角膜乳突通常出现在夜行型昆虫复眼表面,在其复眼光学系统中发挥防反射作用,类别编号(Ⅰ-Ⅲ)越高,抗反射功能越有效(Stavengaetal., 2006)。其中Ⅱ类和Ⅲ类,有效减少眼睛表面的光线反射,为夜行型昆虫提供伪装,并且在空气和角膜材料之间提供折射率的分级转换,显著增加进入角膜晶锥的光量效率(Meyer-Rochowetal., 1979)。柚木野螟复眼的角膜乳头高度为255.95±35.05 nm,属于Ⅲ类,具有多波段抗反射的作用。此外柚木野螟小眼间偶见感觉毛,但是否利于其感知风速和进行飞翔调控还需进一步研究。


图7 柚木野螟成虫小眼结构示意图
在许多夜行性昆虫的重叠像眼中,主要通过瞳孔反应来适应明暗环境的变化(Warrantetal., 1996),主要表现为初级和次级色素细胞内的色素颗粒在光适应条件下向近侧迁移以减少视网膜处的光通量,在暗适应条件下向远侧迁移以增加光通量,如双翅目(Diptera)黑带食蚜蝇Episyrphusbalteatus、橘小实蝇Bactroceradorsalis(张胜杰等, 2015; 刘欢等, 2017)和脉翅目(Neuroptera)大草蛉Chrysopapallens(张海强等, 2007)等。通常重叠像眼小眼数量越多、焦距越短、瞳孔和视杆的直径越大,其光学灵敏度越高(杨小凡等, 2021)。电生理学研究表明,色素颗粒的纵向迁移可使重叠像眼灵敏度提升3个量级单位,极暗环境下,重叠像眼感光细胞的光传导具有级联反应,最大可提高6个量级单位(Bernhardetal., 1960a, 1960b; Warrantetal., 1996)。同时,夜蛾类复眼不仅通过色素颗粒移动调节和适应明暗条件,而且其明暗适应的状态可随光暗环境强度变化而转化。近来对草地贪夜蛾Spodopterafrugiperda的研究结果,证明了昆虫复眼明暗适应状态转化与趋光性的关系(蒋月丽等, 2021)。本研究中,柚木野螟试虫是在光适应下解剖,可观察到色素颗粒均匀分布于透明区、视网膜细胞和色素细胞中(图4: B),但柚木野螟是否也是通过色素颗粒的纵向移动来适应不同明暗环境尚待深入研究,后期可进一步探索柚木野螟趋光行为的发生和调控机制。
