五台山亚高山-高山草甸群落多样性和碳氮磷化学计量特征
2022-09-16张晓龙牛俊杰张殷波史利江郑元润
张晓龙,秦 浩,牛俊杰,张殷波,史利江,郑元润
1. 山西财经大学资源环境学院,山西 太原 030006
2. 山西财经大学统计学院,山西 太原 030006
3. 太原师范学院汾河流域科学发展研究中心,山西 太原 030619
4. 中国科学院植物研究所,北京 100093
山地草甸群落是山地生态系统重要的初级生产者,尤其是在亚高山-高山生态系统中,山地草甸在维持其生物多样性、涵养水源以及提供关键生态系统服务等方面发挥着重要作用[1-3]. 不同于其他山地地带性植被,山地草甸群落分布在山地从低海拔到高海拔的不同植被类型中,其群落组成、结构、多样性以及群落碳氮磷化学计量特征受海拔变化的影响可能存在较大差异[4-7]. 目前关于山地植被的研究多集中于地带性植被的群落生态学特性和多样性特征对海拔梯度的响应方面[8-9],而对山地植被碳氮磷化学计量特征及其对自然环境梯度变化响应规律的研究比较缺乏[10-11],尤其缺乏对连续海拔梯度上山地草甸群落碳氮磷化学计量特征的研究.
五台山地区山地草甸群落垂直分布范围广泛,由于海拔的变化,形成了华北地区最为典型且仅有的亚高山-高山草甸[12-14],这为研究高寒地区山地草甸群落多样性、碳氮磷化学计量特征及其对海拔变化的响应提供了很好的条件. 在连续海拔梯度上,亚高山-高山草甸群落多样性有哪些特点?不同海拔条件下草甸群落对碳氮磷吸收的差异性和趋同性如何?海拔和土壤理化属性变化对亚高山-高山草甸群落多样性及碳氮磷化学计量特征的影响如何?其相对重要性如何?这些问题尚未得到很好的阐释.
为验证和探讨上述问题,该研究以五台山南坡沿海拔梯度自然分布的亚高山-高山草甸植物群落为研究对象,探究连续海拔梯度上亚高山-高山草甸群落多样性及碳氮磷化学计量特征,分析亚高山-高山草甸群落多样性与碳氮磷化学计量特征的关系,定量揭示其与环境因子的关系,这不仅对高寒地区草甸植被资源的利用和保护具有重要意义,也有助于丰富山地草甸生态系统生态化学计量学理论.
1 材料与方法
1.1 样地概况
研究区域包括五台山南坡自然分布的亚高山草甸、高山草甸,地理位置为39°03.08′N~39°04.74′N、113°33.69′E~113°34.23′E,海拔范围为2 201~3 011 m(见表1). 该区域属于典型温带大陆季风气候,气候和植被受地形影响较大,年均温-5~6 ℃,年均降水量500~1 000 mm[8,15]. 亚高山-高山草甸的优势种主要有铁 杆蒿 (Artemisia sacrorum)、 披 针 苔 草 (Carex lanceolata)、早熟禾(Poa annua)、早春苔草(Carex subpediformis)、高山嵩草(Kobresia pygmaea)等(见表1).
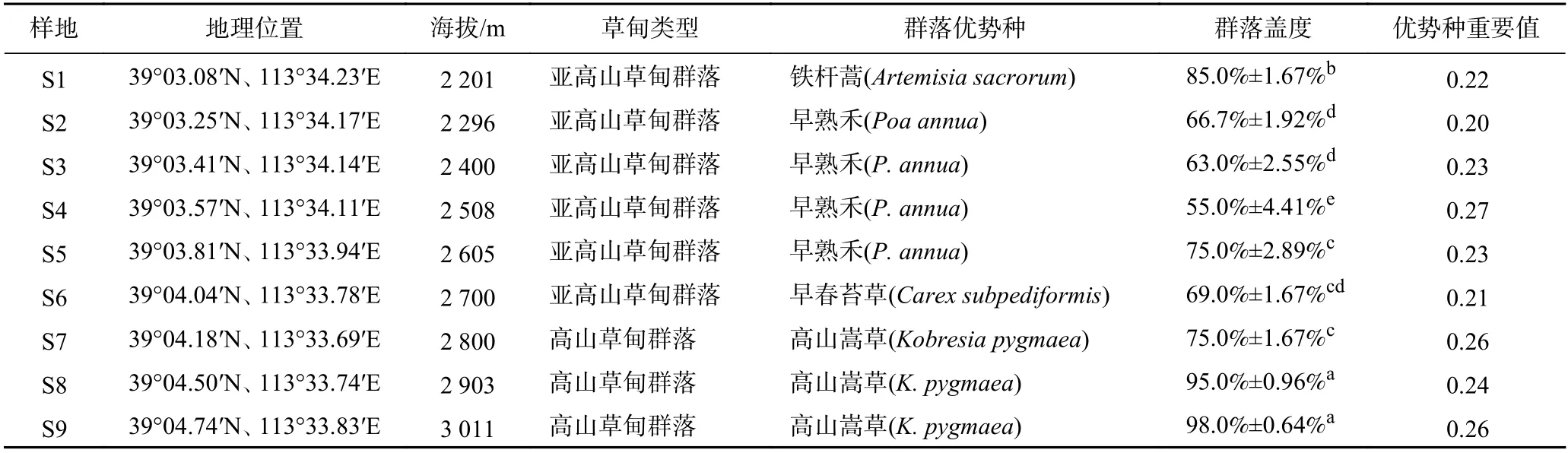
表1 五台山亚高山-高山草甸群落样地特征Table 1 Sampling site characteristics of subalpine-alpine meadow community on Wutai Mountain
1.2 样品采集及测定
于2019 年7-8 月植物生长旺季,沿五台山南坡草地村至北台山顶一线,大致以100 m 海拔为间隔,在海拔2 201~3 011 m 范围内布设9 个调查样地(见图1). 在海拔梯度上,每个样地设3 个重复样方,样方大小为1 m×1 m. 草甸群落优势种由重要值决定,物种重要值最大则为优势种[8],研究区域内群落优势种依次为铁杆蒿(S1)、早熟禾(S2、S3、S4、S5)、早春苔草(S6)、高山嵩草(S7、S8、S9). 在每个草本样方内,调查植物物种种类、株数、高度、盖度等基本群落指标,收集草本样方地上部分用于化学分析. 在与植物群落相应的样地内,分层采集0~10 cm 和10~20 cm土样,每个样地设3 个重复,植物样品与土壤样品分别装入保鲜盒后带回实验室.

图1 研究区样地位置示意Fig.1 Locations of sample plots
土壤含水量和土壤容重分别采用烘干法和环刀法测定[16-17];用于测定TC、TN、TP 含量的植物样品在烘箱内85 ℃下烘干至恒质量,粉碎后过0.149 mm 筛备用;用于测定化学属性的土壤样品自然风干后,粉碎过0.149 mm 筛备用;植物TC 含量、植物TN 含量、SOC(土壤有机碳)含量、土壤TN 含量均采用 C/H/N 元 素 分 析 仪 (Vario EL Ⅲ,Hanau,Germany)测定;植物TP 含量和土壤TP 含量采用电感耦合等离子体发射光谱仪(iCAP 6300,Waltham,USA)测定;土壤pH 和土壤电导率分别采用酸度计法和电导仪法测定[16]. 土壤属性为0~20 cm 平均值,元素含量以质量百分比表示.
1.3 数据分析
在数据满足正态分布和方差齐性的基础上,采用单因素方差分析比较不同海拔条件下群落盖度、群落多样性、群落碳氮磷化学计量特征之间的差异性(P<0.05),多重比较由LSD 检验完成. 对于不满足方差齐性的数据,进行数据转换,数据转换后依然不满足方差齐性,则采用非参数检验[18]. 采用Pearson 相关性分析草甸群落多样性与碳氮磷化学计量特征之间的关系,显著性水平P<0.05[19]. 为研究海拔与土壤因子对亚高山-高山草甸群落多样性及碳氮磷化学计量特征变化的影响,使用Pearson 相关系数分析群落多样性及碳氮磷化学计量特征与海拔、土壤因子之间的关系,通过偏冗余分析(pRDA)确定海拔、土壤因子的单独贡献率以及二者交互作用的贡献率,统计分析在SPSS 18.0 和Canoco 5.0 中完成[19-20].
多样性指数选取植物群落Patrick 指数、Shannon-Wiener 指数、Simpson 指数和Pielou 指数[8,14],公式如下:
Patrick 指数(R):

Shannon-Wiener 指数(H′):
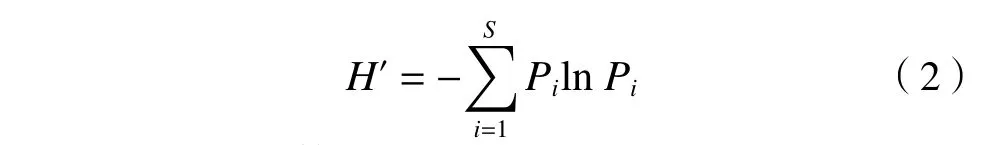
Simpson 指数(D):

Pielou 指数(E):
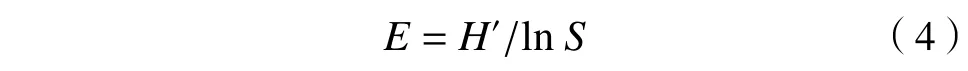
式中,S为物种数,Pi为物种i的重要值.
2 结果与分析
2.1 群落多样性特征随海拔的变化
在海拔梯度(2 201~3 011 m)上,山地草甸类型依次分布有亚高山草甸(2 201~2 800 m)、高山草甸(2 800~3 011 m),植物群落优势种由早熟禾(禾草类)、早春苔草(苔草类)向高寒嵩草类植物过渡(见表1).随着海拔的升高,群落盖度呈先下降后上升的显著变化趋势,群落盖度在海拔2 508 m 处达到最小值,为55.0%;在3 011 m 处达到最大值,为98.0%(见表1).
在海拔梯度上,亚高山-高山草甸群落的Patrick指数(F=9.52,P<0.001)、Shannon-Wiener 指 数(F=10.32,P<0.001)和Simpson 指数(F=2.86,P=0.031)在不同样地间均具有显著性差异,而Pielou 指数(F=0.16,P=0.994)差异不显著(见表2). 随着海拔的升高,Patrick 指 数、Shannon-Wiener 指 数 和Simpson 指 数均呈显著下降趋势,最小值出现在海拔3 011 m 处,分别为7、1.84、0.82(见图2). Pielou 指数随着海拔变化呈平稳趋势,变化不显著(见图2).

表2 不同海拔条件下山地草甸群落多样性和碳氮磷化学计量特征的单因素方差分析Table 2 One-way ANOVA of mountain meadow community diversity and C, N and P stoichiometric traits at different altitudes

图2 不同海拔下亚高山-高山草甸群落的α 多样性指数Fig.2 Subalpine-alpine meadow community α diversity indices among different altitudes
2.2 群落碳氮磷化学计量特征随海拔的变化
亚高山-高山草甸群落TC 含量平均值为461.19 mg/g,TN 含量平均值为23.32 mg/g,TP 含量平均值为1.96 mg/g,群落C∶N、C∶P 和N∶P 平均值分别为19.99、242.17、12.10,其中群落TP 含量变异系数最大(见表2). 在海拔梯度上,群落TC 含量(F=16.27,P<0.001)、TN 含量(F=8.03,P<0.001)、TP 含量(F=3.68,P=0.010)、C∶N(F=14.91,P<0.001)和C∶P(F=4.21,P=0.005)在不同样地间均具有显著性差异,而N∶P(F=1.14,P=0.385)差异不显著(见表2).
随着海拔的升高,亚高山-高山草甸群落TC 含量呈显著上升趋势,而TN、TP 含量随着海拔的升高呈显著下降趋势;C∶N 和C∶P 随着海拔的升高均呈显著上升趋势,而N∶P 变化趋势不显著(见图3).

图3 不同海拔下亚高山-高山草甸群落碳氮磷含量及其计量比Fig.3 Subalpine-alpine meadow community C, N and P contents and stoichiometric ratios among different altitudes
2.3 群落多样性和碳氮磷化学计量特征之间的相关性
在海拔梯度上,亚高山-高山草甸群落的Patrick指数与Shannon-Wiener 指数、Simpson 指数均呈显著正 相 关,而Patrick 指 数、Shannon-Wiener 指 数、Simpson 指数均与Pielou 指数无显著相关性;Patrick指数、Shannon-Wiener 指数均与群落TN 含量、TP 含量呈显著正相关,而Patrick 指数、Shannon-Wiener 指数均与群落TC 含量、C∶N、C∶P 呈显著负相关(见图4). 此外,亚高山-高山草甸群落的Simpson 指数与群落TC 含量、C∶N 均呈显著负相关(见图4).
亚高山-高山草甸群落TC 含量与TN 含量、TP含量呈显著负相关;TN 含量与TP 含量呈显著正相关,而TN 含量与C∶N、C∶P 均呈显著负相关(见图4). 亚高山-高山草甸群落TP 含量与C∶N、C∶P、N∶P 均呈显著负相关,C∶P 与C∶N、N∶P 均呈显著正相关(见图4).
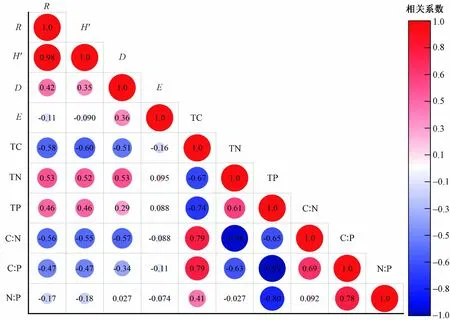
图4 亚高山-高山草甸群落多样性和碳氮磷化学计量特征之间的相关性Fig.4 Correlation analysis between subalpine-alpine meadow community diversity and C, N, P stoichiometric traits
2.4 海拔与土壤因子对群落多样性特征变化的解释贡献率
相关性分析结果表明,亚高山-高山草甸群落的Patrick 指数、Shannon-Wiener 指数均与海拔、土壤含水量、土壤有机碳(SOC)含量呈显著负相关,而与土壤pH 呈显著正相关(见表3). Simpson 指数与海拔、土壤电导率、土壤有机碳(SOC)含量、土壤TN 含量均呈显著负相关(见表3). 偏冗余分析结果显示,海拔与土壤因子共同解释了亚高山-高山草甸群落多样性特征总变化的70.8%,海拔与土壤因子分别独自解释了总变化的45.8%和14.4%,海拔与土壤因子的交互作用解释了总变化的10.6%,其中海拔因子的单独解释率占比最大.
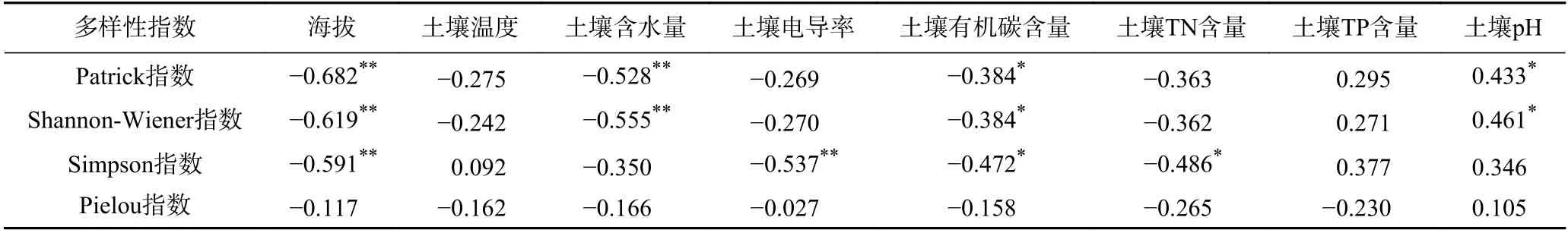
表3 亚高山-高山草甸群落多样性和环境因子之间的相关性Table 3 Correlation analysis between subalpine-alpine meadow community diversity and environmental factors
2.5 海拔与土壤因子对群落碳氮磷化学计量特征变化的解释贡献率
相关性分析结果表明,亚高山-高山草甸群落TC 含量、C∶N、C∶P 均与海拔、土壤含水量、土壤电导率、土壤有机碳(SOC)含量、土壤TN 含量呈显著正相关,而与土壤pH 呈显著负相关(见表4). 亚高山-高山草甸群落TN 含量、TP 含量均与海拔、土壤含水量、土壤电导率、土壤有机碳(SOC)含量、土壤TN含量呈显著负相关,而与土壤pH 呈显著正相关(见表4). 偏冗余分析结果显示,海拔与土壤因子共同解释了亚高山-高山草甸群落碳氮磷化学计量特征总变化的67.8%,海拔与土壤因子分别独自解释了总变化的54.4%和6.7%,海拔与土壤因子的交互作用解释了总变化的6.7%,其中海拔因子的单独解释率占比最大.

表4 亚高山-高山草甸群落碳氮磷化学计量特征和环境因子之间的相关性Table 4 Correlation analysis between subalpine-alpine meadow community C, N, P stoichiometric traits and environmental factors
3 讨论
3.1 群落多样性格局及影响因素分析
在山区,海拔是调控和限制植物群落变化的主要影响因子,植物群落组成、结构和功能均会随着海拔变化引起的水热条件变化而发生改变[21-22]. 在五台山南坡2 201~3 011 m 的海拔范围内,重要值很好地反映了亚高山-高山草甸群落的垂直梯度变化规律,随着海拔的升高,依次分布有铁杆蒿群落(2 201~2 296 m)、早熟禾群落(2 296~2 700 m)、早春苔草群落(2 700~2 800 m)、高山嵩草群落(2 800~3 011 m). 在海拔梯度上,亚高山-高山草甸群落盖度在不同样地间差异显著,大体呈先下降后上升的变化趋势,其中群落盖度在2 296~2 800 m 之间相对较低(见表1),主要是由该海拔段灌木层和草本层之间相互竞争引起的[8,22].在2 200 m 海拔以上,金露梅(Potentilla fruticosa)、鬼箭锦鸡儿(Caragana jubata)灌木物种出现,受上层亚高山灌木层的影响,草本层盖度有所下降,尤其是在2 508 m 处,灌木层盖度达到最大值(80.0%),此海拔段草本层盖度降到最小值(55.0%). 在海拔2 800 m以上,由于水热条件变差,仅分布有耐寒草本物种,灌木物种消失,草甸群落盖度呈上升趋势(见表1),在一定程度上也说明该区域山地植被的生长和分布受海拔变化的制约.
五台山亚高山-高山草甸群落的Patrick 指数、Shannon-Wiener 指数和Simpson 指数均随着海拔的升高呈显著下降趋势,这与河北小五台山[21]、北京东灵山[23]等山区的研究结果相似,这种格局的形成与海拔垂直变化密切相关,即随着海拔的升高,植物群落多样性趋于减少. 与其他指数不同,Pielou 指数在海拔梯度上变化并不显著,这主要是由于亚高山-高山地区寒冷低温的生境对植物物种形成生理限制,使得亚高山-高山草甸群落物种相对稳定,偶见种、稀有种少见,物种分布相对均匀[24]. 偏冗余分析结果表明,海拔和土壤因子较好地解释了该区域亚高山-高山草甸群落多样性随海拔变化而变化的特征,说明海拔变化显著影响着该区域亚高山-高山草甸群落多样性. 在所有解释因子中,海拔与土壤因子共同解释了亚高山-高山草甸群落多样性特征总变化的70.8%,其中海拔较土壤因子的解释率更高,表明该区域海拔变化更易成为山地草甸群落物种多样性的限制因素,且与土壤因子互相作用共同影响着山地草甸群落多样性特征.
3.2 碳氮磷化学计量格局及影响因素分析
在五台山南坡2 201~3 011 m 海拔范围内,亚高山-高山草甸群落TC 含量平均值为461.19 mg/g,略高于中国草地生态系统植物TC 含量平均值(442.4 mg/g)[25],与全球陆地植物叶片TC 含量平均值(461.6 mg/g)[26]相当;群落TN 含量平均值为23.32 mg/g,略低于中国草地生态系统植物TN 含量平均值(26.5 mg/g)[25,27],高于全球陆地植物叶片TN 含量平均值(20.1 mg/g)[26,28];群落TP 含量平均值为1.96 mg/g,略高于中国草地生态系统植物TP 含量平均值(1.91 mg/g)[25,27]和全球陆地植物叶片TP 含量平均值(1.77 mg/g)[26,28],这也可能是导致该区域草甸群落C∶P 和N∶P 相对较低的原因. 与全球尺度和草地生态系统区域尺度的研究结果相比,五台山亚高山-高山草甸群落整体具有相对稳定的TC 含量,以及TN、TP 含量高和N∶P 低的特点. 在海拔梯度上,亚高山-高山草甸群落TP 含量的变异系数高于群落TC 含量和TN含量(见表2),表明该区域草甸群落磷元素稳定性相对较差,且化学计量内稳性比碳、氮元素低. 该区域亚高山-高山草甸群落C、N、P 计量比存在差异性,其中C∶P 变异系数最大,N∶P 变异系数相对较小(见表2),这与长白山、贺兰山等山区的研究结果[29-30]相类似. C∶P 变异系数较大,主要是由于群落TP 含量的较大差异造成的,而群落N∶P 变异系数相对较小,可能是由于氮元素和磷元素之间较强的耦合关系所致[29,31]. 该研究中群落TN 含量与TP 含量呈显著正相关(见图4),在一定程度上说明了该区域氮、磷元素之间较强的耦合关系. 此外,相关分析显示,亚高山-高山草甸群落TP 含量与N∶P 呈显著负相关,而TN 含量与N∶P 没有显著相关性(见图4),表明该区域草甸群落TP 含量的变异对群落N∶P 在海拔梯度上的变化起主导作用.
在海拔梯度上,亚高山-高山草甸群落TC 含量随海拔升高而显著增加(见图3),这主要是由高寒地区植物体内碳元素积累增加引起的[29]. 有研究表明,与低海拔植物相比,海拔较高的植物受到低温、潮湿等多种环境胁迫的作用,主要通过增加体内非结构性碳物质(如淀粉、果聚糖等)的含量用以平衡细胞的渗透压并抵抗冻结,从而适应高海拔生境[29,32];其次,在高海拔地区相对较强的辐射条件下,植物体内碳积累增加[29,33],这些因素导致高海拔草甸群落具有相对较高的碳含量. 随着海拔的升高,亚高山-高山草甸群落TN、TP 含量呈显著降低趋势(见图3),这主要与高寒草甸抵抗低温环境的适应机制关系密切. 随着海拔的升高,低温环境限制土壤微生物的活动,导致土壤中有机质分解速率降低,植物根部可吸收有效养分减少[30,34]. 此外,高海拔地区降水增多导致土壤淋溶作用加剧,养分条件变差[35],这些因素会导致草甸群落TN、TP 含量随海拔升高而降低.
植物群落C、N、P 计量比可在一定程度上反映当地植物碳积累动态以及氮磷养分限制状况[34,36]. 该研究中亚高山-高山草甸群落C∶N、C∶P 随海拔升高均呈显著增加趋势(见图3),这主要是不同海拔条件下草甸群落对不同生境适应的结果. 高海拔草甸群落为了在低温环境下存活,植物倾向于较低的生长速率,代谢活动相对较慢,受高山光照条件影响,碳积累增加[29-30],这样植物体内相对较高碳含量和相对较低的氮、磷含量使得高海拔草甸群落C∶N 和C∶P 相对较高(见图3). 此外,相对较低海拔段的草甸群落生境水热条件相对较好,倾向于较快的生长速率,这可能是导致低海拔草甸群落C∶N 和C∶P 相对较低的主要原因[37-38]. 与C∶N 和C∶P 随海拔梯度的变化趋势不同,群落N∶P 随海拔升高的变化不显著,海拔梯度上N∶P 平均值为12.10(N∶P<14,主要受氮限制),相对较低的N∶P 表明该区域草甸群落在生长旺季更倾向于受氮元素的限制[39-40]. 偏冗余分析结果表明,海拔与土壤因子共同解释了山地草甸群落碳氮磷化学计量特征总变化的67.8%,海拔对草甸群落碳氮磷化学计量特征变化的解释贡献率大于土壤因子以及海拔与土壤因子交互作用的贡献率,进一步表明该区域海拔变化影响着亚高山-高山草甸群落植物生长发育和养分利用状况.
4 结论
a) 在五台山南坡2 201~3 011 m 海拔范围内,重要值能很好地反映出亚高山-高山草甸群落随海拔梯度的变化情况,随着海拔的升高,依次分布有铁杆蒿群落(2 201~2 296 m)、早熟禾群落(2 296~2 700 m)、早 春 苔 草 群落(2 700~2 800 m)、高 山 嵩 草 群 落(2 800~3 011 m). 在 海 拔 梯 度 上,Patrick 指 数、Shannon-Wiener 指数和Simpson 指数在不同样地间存在显著性差异,而群落Pielou 指数变化不显著.Patrick 指 数、Shannon-Wiener 指 数 和Simpson 指 数随着海拔的升高均呈显著下降趋势,在3 011 m 处达到最小值,分别为7、1.84、0.82.
b) 五台山南坡亚高山-高山草甸群落总体上具有相对稳定的TC 含量,以及TN、TP 含量高和N∶P 低的特点. TC 含量、TN 含量、TP 含量、C∶N 和C∶P 在不同海拔样地间存在显著性差异,体现出不同海拔条件下草甸群落对海拔生境的不同适应策略. 随着海拔的升高,TC 含量、C∶N 和C∶P 随着海拔升高均呈显著上升趋势,TN、TP 含量均呈显著下降趋势,而N∶P 变化不显著. 群落水平N∶P(平均值为12.10)的分析结果表明,该区域草甸群落在生长旺季可能面临磷元素过量而氮元素相对不足的情况.
c) 海拔变化对亚高山-高山草甸群落多样性和碳氮磷化学计量特征影响显著,群落多样性和碳氮磷化学计量特征与土壤理化属性存在一定相关性. 偏冗余分析(pRDA)结果表明,海拔和土壤因子较好地解释了亚高山-高山草甸群落多样性和碳氮磷化学计量的变化特征,解释率分别为70.8%、67.8%,其中海拔因子的解释率最高,较土壤因子更为重要,说明海拔变化是限制该区域亚高山-高山草甸群落植物生长的关键因素.
