异色瓢虫和龟纹瓢虫幼虫对番茄潜叶蛾低龄幼虫的捕食功能反应
2022-09-16杨桂群蒋正雄胡昌雄陈国华张晓明
杨桂群,范 苇,张 倩,李 貌,蒋正雄,段 盼,胡昌雄,陈国华,张晓明
(云南农业大学植物保护学院/云南生物资源保护与利用国家重点实验室,昆明 650201)
番茄潜叶蛾Tuta absoluta隶属于鳞翅目Lepidoptera、麦蛾科Gelechiidae,又名南美番茄潜叶蛾、番茄麦蛾。该虫生活史包括卵、幼虫、蛹、成虫四个发育阶段,繁殖能力强,世代重叠严重;主要以幼虫进行为害,可在番茄植株的任一发育阶段和任一地上部位进行为害,幼虫包括四个龄期,主要潜食叶肉为害,当幼虫龄期比较大时,还可以蛀食顶梢、嫩芽、嫩茎和果实,具有转移叶片和转株为害的习性,严重影响作物的品质和产量[1-3]。该虫为多食性害虫,其寄主植物范围广泛,目前为止可为害的植物达9科39种植物,主要嗜食番茄Lycopersicon esculentum、马铃薯Solanum tuberosum、茄子S. melongena、龙葵S. nigrum等茄科植物[4,5]。自20世纪50年代起,番茄潜叶蛾一直作为南美洲番茄上的毁灭性害虫被广泛关注,2006年该虫传入西班牙并迅速传播扩散至地中海沿岸其他国家和欧洲各国,此后以极快的传播速度在欧亚非大陆扩散,截至2017年5月,已在欧洲、南美洲、非洲、中美洲、亚洲等85个国家或地区发生[6,7]。该虫于2017年8月在我国新疆伊犁的番茄地首次被发现,目前仅有报道在我国新疆和云南等局部地区区域性发生为害,但其环境适应性和成灾性极强,具有高度潜在的扩散风险,能在我国大部分番茄产区定殖[2,8]。番茄、马铃薯在我国各省市广泛种植,种植面积和总产量均位居世界前列,且有逐年增长趋势,这为番茄潜叶蛾的扩散和为害提供了良好的生态条件,番茄潜叶蛾一旦传播扩散开来,将造成巨大的危害和经济损失[9,10]。
自该虫入侵我国以来,我国相应的地区相继开展了防治研究工作,目前番茄潜叶蛾的防治仍然以化学杀虫剂为主,但由于幼虫潜叶为害,化学防治效果并不理想[11]。同时,大量不合理的杀虫剂使用反而加剧了该虫抗药性的发展,已报道番茄潜叶蛾对多杀菌素、氯虫苯甲酰胺等在内的多种类型的杀虫剂产生了不同程度的抗性[12,13]。因此,建立科学的防控技术体系,研究和利用天敌昆虫开展生物防治是番茄潜叶蛾治理的重要方法。国外有关番茄潜叶蛾的生物防治研究已有诸多报道,例如,在欧洲天敌烟盲蝽Nesidiocoris tenuis、蝽象Macrolophus pygmaeus通过取食番茄潜叶蛾的卵和幼虫可以有效控制该害虫在番茄上的种群[14];此外还有许多寄生性天敌,如赤眼蜂类天敌昆虫可以通过寄生番茄潜叶蛾的卵、寡节小蜂科和茧蜂科昆虫可以通过寄生番茄潜叶蛾的幼虫来控制番茄潜叶蛾的发生为害[15,16]。因此,充分发挥本地天敌昆虫对番茄潜叶蛾的控制作用,明确可供利用的本地天敌昆虫资源是番茄潜叶蛾防治工作的重点之一。
瓢虫是一类应用前景良好的重要捕食性天敌昆虫,其中异色瓢虫Harmonia axyridis和龟纹瓢虫Propylea japonica是我国番茄潜叶蛾发生或潜在发生区常见的重要捕食性天敌类群,可以取食多种蚜虫、叶蝉及鳞翅目幼虫,在我国广泛分布,在众多生境中均能发挥较好的捕食能力,在害虫的自然控制中起到重要作用[17,18]。龟纹瓢虫和异色瓢虫在生物防治中的研究和应用主要集中于半翅目和部分鳞翅目幼虫,如目前在控制柑橘木虱Diaphorina citr[19]、草地贪夜蛾Spodoptera frugiperda[20]、刺槐蚜Aphis robiniae[21]等害虫方面发挥了重要作用,且已实现一定规模的商品化生产及应用。异色瓢虫和龟纹瓢虫的4龄幼虫由于即将进入蛹期其食量大增,捕食能力最强,且比较容易饲养,是捕食功能评价的重要材料[22-24]。为探明两种瓢虫对番茄潜叶蛾控害能力,寻找到番茄潜叶蛾的有效生物防治材料,本研究采用两种瓢虫的4龄幼虫,在室内条件下评价其对番茄潜叶蛾低龄幼虫(1龄、2龄)的捕食能力,以期为新入侵重大农业害虫番茄潜叶蛾的生物防治提供科学参考。
1 材料与方法
1.1 供试虫源及条件
番茄潜叶蛾:2020年6月采自云南省玉溪市阳瑞农业科技发展有限公司有机农场(24°20′ N,102°35′E)的番茄植株上,在室内置于养虫笼(30 cm×30 cm×30 cm)中,在人工气候箱(LTC-1000,上海三腾仪器)内用番茄植株继代饲养。挑取生长大小一致的同代番茄潜叶蛾低龄幼虫(1龄幼虫和2龄幼虫)作为供试虫源,虫龄划分参考张桂芬等[4]的鉴定方法。
异色瓢虫和龟纹瓢虫:采自云南农业大学后山农场的玉米田中,在室内用番茄潜叶蛾幼虫喂食,在人工气候箱内以番茄潜叶蛾幼虫继代饲养。饲养获得的4龄幼虫作为供试昆虫用于捕食试验。人工气候箱设置条件:温度(25±1)℃、相对湿度(70±5)%、光周期16 L:8 D。
1.2 异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的捕食功能反应
分别对番茄潜叶蛾1龄幼虫、2龄幼虫进行试验,番茄潜叶蛾1龄幼虫密度设置分别为20、40、60、80和100头/皿,2龄幼虫密度设置分别为10、30、50、70和90头/皿,每个密度设置5个重复。试验前挑选行动敏捷、龄期一致的异色瓢虫和龟纹瓢虫4龄幼虫单头置于培养皿(直径3.50 cm,高1.50 cm)内进行饥饿处理24 h。试验当天在培养皿底铺放一层润湿的滤纸,每皿均匀放置10片剪取自新鲜番茄叶片的0.50 cm×0.50 cm叶碟供番茄潜叶蛾取食,然后用细毛刷接入不同密度的番茄潜叶蛾幼虫,然后分别接入饥饿处理后的单头(异色瓢虫/龟纹瓢虫)4龄幼虫,用保鲜膜封口,并用3号昆虫针在保鲜膜上扎若干密集小孔透气,置于人工气候箱中培养。24 h后在解剖镜下统计每皿中剩余虫体完整的番茄潜叶蛾幼虫数,计算异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾幼虫的单日捕食量,并分别计算出两种天敌对猎物的单日捕食率,单日捕食率=单日捕食量/猎物密度[25]。
采用Holling II型捕食功能反应模型对观察值进行拟合,Holling II型圆盘方程为:Na=a′TN/(1+a′ThN),式中Na为番茄潜叶蛾幼虫被捕食数量,N为番茄潜叶蛾幼虫的密度,a′为捕食者对猎物的瞬时攻击率,T是猎物暴露于捕食者的时间(即试验的总时间,T取1 d),Th为捕食者捕食1头猎物所用的时间,a′/Th是捕食者对猎物的捕食能力,1/Th为捕食者对猎物的日最大捕食量(即捕食上限Namax);将公式变形为1/Na=1/a′TN+Th/T,利用直线回归方程求得Th和a′[26,27]。
1.3 异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的搜寻效应
利用拟合Holling II型捕食功能反应模型所得参数,根据猎物密度估算异色瓢虫、龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的搜寻效应S。搜寻效应方程为:S=a′/(1+a′ThN),式中S为搜寻效应,参数同1.2节。
1.4 数据统计与分析
采用SPSS 26.0软件进行数据分析以及Origin 2018拟合直线回归方程,采用单因素Duncan’s新复极差法(P<0.05)比较差异显著性。
2 结果与分析
2.1 异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的捕食功能反应
异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的日捕食量随番茄潜叶蛾数量的增大而增大:当猎物为番茄潜叶蛾1龄幼虫,密度为20头/皿时,异色瓢虫和龟纹瓢虫的日捕食量分别为19.60头和15.80头;猎物密度增至100头/皿,异色瓢虫和龟纹瓢虫的日捕食量增至70.80头和61.60头。当猎物为番茄潜叶蛾2龄幼虫,密度为10头/皿时,异色瓢虫和龟纹瓢虫的日捕食量分别为9.80头和6.40头;猎物密度增至90头/皿,异色瓢虫和龟纹瓢虫的日捕食量增至64.20头和48.40头。当猎物密度N→+∞时,异色瓢虫和龟纹瓢虫对番茄潜叶蛾1、2龄幼虫的理论日最大捕食量Namax分别为147.06头、140.85头和333.33头、149.25头。当猎物密度为10~20头/皿时,两种瓢虫的日捕食量近似等于每皿猎物数量,随着猎物密度的增大,两种瓢虫的日捕食量增幅逐渐减缓(图1、图2)。

图1 异色瓢虫4龄幼虫对番茄潜叶蛾1和2龄幼虫的捕食功能反应曲线Fig. 1 Functional response curves of 4th instar larvae Harmonia axyridis to 1st and 2nd instar larvae Tuta absoluta
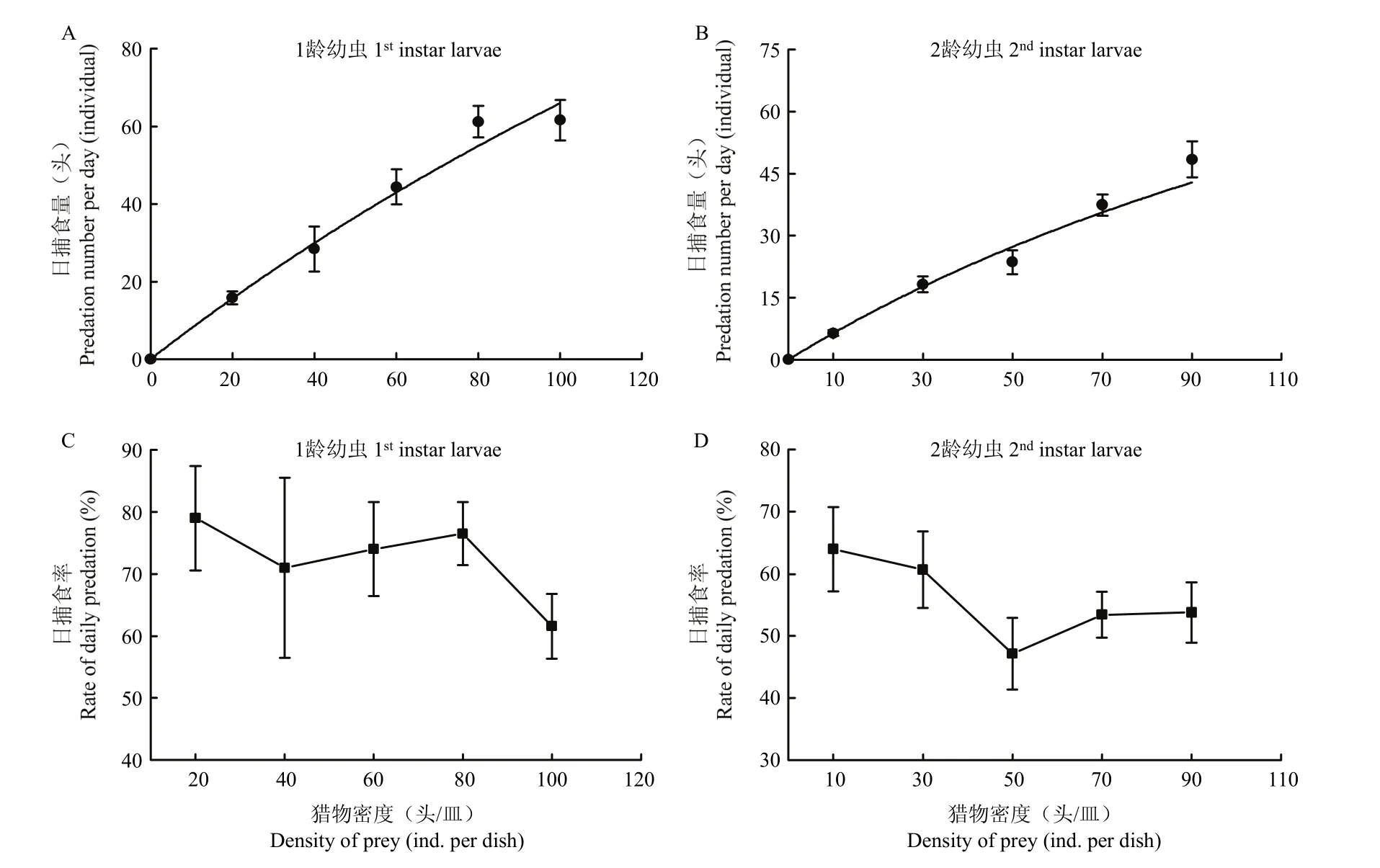
图2 龟纹瓢虫4龄幼虫对番茄潜叶蛾1和2龄幼虫的捕食功能反应曲线Fig. 2 Functional response curves of Propylaea japonica 4th instar larvae to 1st and 2nd instar larvae of Tuta absoluta
异色瓢虫和龟纹瓢虫4龄幼虫的日捕食率随番茄潜叶蛾数量的增大而减小:当猎物为番茄潜叶蛾1龄幼虫,密度为20头/皿,异色瓢虫和龟纹瓢虫的日捕食率为98%和79%;密度增至100头/皿,异色瓢虫和龟纹瓢虫的日捕食率降至71%和62%。当猎物为番茄潜叶蛾2龄幼虫,密度为10头/皿,异色瓢虫和龟纹瓢虫的日捕食率为98%和64%;密度增至90头/皿,异色瓢虫和龟纹瓢虫的日捕食率降至71%和54%。且异色瓢虫对番茄潜叶蛾低龄幼虫的日捕食率显著高于龟纹瓢虫(F2,12=17.55,P<0.05;F2,12=12.17,P<0.05)(图1、图2)。
采用Holling II型捕食功能反应模型拟合得到异色瓢虫和龟纹瓢虫对番茄潜叶蛾低龄幼虫的捕食功能反应及相关参数如表1所示,异色瓢虫对番茄潜叶蛾1、2龄幼虫的瞬时攻击率分别为1.10和1.04;龟纹瓢虫对番茄潜叶蛾1、2龄幼虫的瞬时攻击率分别为0.82和0.67,与龟纹瓢虫相比异色瓢虫对猎物的瞬时攻击率更高;异色瓢虫和龟纹瓢虫处理猎物的时间则相反,处理1龄的时间分别为0.0068 d和0.0030 d,处理2龄的时间分别为0.0071 d和0.0067 d,即与异色瓢虫相比,龟纹瓢虫处理猎物的时间更短;异色瓢虫对番茄潜叶蛾1、2龄幼虫的捕食能力分别为161.67和146.58;龟纹瓢虫对番茄潜叶蛾1、2龄幼虫的捕食能力分别为333.33和149.25,两种瓢虫捕食1龄幼虫的能力更强。

表1 异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的捕食功能方程及反应参数Table 1 Functional response equation and parameters of 4th instar larvae Harmonia axyridis and Propylaea japonica to low instar larvae Tuta absoluta
2.2 异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾低龄幼虫的搜寻效应
两种瓢虫的搜寻效应与番茄潜叶蛾的密度具有负相关性:异色瓢虫对番茄潜叶蛾1、2龄幼虫的搜寻效应与猎物密度的相关性方程分别为Na=1.0196—0.0041N和Na=0.9919—0.0043N,龟纹瓢虫对番茄潜叶蛾1、2龄幼虫的搜寻效应与猎物密度之间的相关性方程为Na=0.8119—0.0015N和Na=0.6536—0.002N,且均满足R2>0.98,即随着番茄潜叶蛾密度的增加,两种瓢虫对番茄潜叶蛾 1、2龄幼虫的搜寻效应均逐渐降低。在同一猎物密度下,两种瓢虫的搜寻效应有明显差异:异色瓢虫对番茄潜叶蛾低龄幼虫的搜寻效应显著高于龟纹瓢虫(F2,12=17.55,P<0.05;F2,12=12.17,P<0.05),异色瓢虫对番茄潜叶蛾1、2龄幼虫的最高搜寻效应高于0.90,而龟纹瓢虫对番茄潜叶蛾的最高搜寻效应低于0.80(图3、图4)。

图3 异色瓢虫4龄幼虫对番茄潜叶蛾1和2龄幼虫的搜寻效应Fig. 3 Searching efficiency rate of Harmonia axyridis 4th instar larvae to 1st and 2nd instar larvae of Tuta absoluta

图4 龟纹瓢虫4龄幼虫对番茄潜叶蛾1和2龄幼虫的搜寻效应Fig. 4 Searching efficiency rate of Propylaea japonica 4th instar larvae to 1st and 2nd instar larvae of Tuta absoluta
3 讨论
Holling II型捕食功能反应模型是反应在固定时间内单头天敌的捕食量随猎物密度改变而发生变化[27]。一般情况下捕食性天敌的捕食数量与猎物密度之间的非线性函数关系大多符合Holling II模型[25]。本研究表明,异色瓢虫和龟纹瓢虫4龄幼虫对番茄潜叶蛾1龄幼虫和2龄幼虫的捕食功能反应均符合Holling II型圆盘方程,这与前人研究中异色瓢虫和龟纹瓢虫对猎物的捕食功能反应一致,如龟纹瓢虫捕食玉米蚜Rhopalosiphum maidis[28],异色瓢虫捕食桃蚜Myzus persicae[29]。龟纹瓢虫和异色瓢虫对番茄潜叶蛾均有较强的捕食能力。研究表明天敌的捕食量和猎物种群数量之间存在相关性,猎物的增多会给天敌增加更多的捕食机会,因此天敌的捕食量会随猎物密度的增大而增加[30,31]。本研究结果显示异色瓢虫和龟纹瓢虫的日捕食量皆随番茄潜叶蛾1、2龄幼虫密度的增加而增加,但随着猎物密度的增加,瓢虫到达饱食状态后,日捕食量增幅逐渐减缓,这与龟纹瓢虫捕食草地贪夜蛾卵和低龄幼虫[20]、玉米蚜[17],异色瓢虫捕食豌豆修尾蚜Megoura japonica[32]、草地贪夜蛾[33]的功能反应一致。异色瓢虫捕食番茄潜叶蛾1、2龄幼虫的最大日捕食量分别为147.06头和140.85头,虽然差异不大,但可以看出对1龄幼虫的捕食量更大,龟纹瓢虫捕食番茄潜叶蛾1、2龄幼虫的最大日捕食量分别为333.33头和149.25头,同样存在对1龄幼虫的捕食量更大的现象,这可能因为低龄幼虫体内所含营养成分较少,也可能与瓢虫对不同虫龄的喜好性不同有关[34]。
Holling[35]认为天敌的搜寻效应依赖于猎物的种群密度,随猎物种群密度的增加而降低。本研究发现,随着猎物密度的增加,异色瓢虫和龟纹瓢虫对番茄潜叶蛾 1、2龄幼虫的搜寻效应均逐渐降低。这与龟纹瓢虫捕食柑橘木虱和异色瓢虫捕食草地贪夜蛾2龄幼虫时对其的搜寻效应类似,这可能是因为在相同的空间内,随着番茄潜叶蛾数量的增加,单位空间内猎物密度增加,缩短了两种瓢虫寻找猎物的时间,降低寻找猎物所需能量,所以搜寻效应下降[19,33,36]。本研究在同一猎物密度下,两种瓢虫的搜寻效应存在差异,异色瓢虫对番茄潜叶蛾的搜寻效应高于龟纹瓢虫对番茄潜叶蛾的搜寻效应,异色瓢虫对番茄潜叶蛾 1、2龄幼虫的最高搜寻效应高于0.90,而龟纹瓢虫对番茄潜叶蛾1、2龄幼虫的最高搜寻效应低于0.80,存在异色瓢虫搜寻效应更高的趋势。搜寻效应是天敌在捕食猎物时对猎物攻击的一种行为效应,其大小与天敌本身存在一定的关系,导致异色瓢虫的搜寻效应比龟纹瓢虫更高的原因可能是异色瓢虫的个体比龟纹瓢虫大,行动敏捷度低于龟纹瓢虫,寻找猎物所需能量大于龟纹瓢虫,所以搜寻效应比龟纹瓢虫高[37]。研究还发现龟纹瓢虫对番茄潜叶蛾2龄幼虫的搜寻效应低于对1龄幼虫的搜寻效应,存在搜寻效应随猎物龄期的增大而降低的现象。孔琳[24]关于四种瓢虫对草地贪夜蛾卵和幼虫捕食的研究同样表明龟纹瓢虫在捕食草地贪夜蛾时,搜寻效应随猎物龄期的增大而降低的现象,导致该现象的原因可能是番茄潜叶蛾2龄幼虫的个体较1龄幼虫的个体大,所以在一定的空间范围内降低了龟纹瓢虫寻找猎物的难度和所需能量,故龟纹瓢虫对番茄潜叶蛾2龄幼虫的搜寻效应低于1龄幼虫。
本研究表明异色瓢虫和龟纹瓢虫4龄幼虫均对番茄潜叶蛾低龄幼虫有较好的捕食能力,均可控制番茄潜叶蛾低龄幼虫的数量,但异色瓢虫的捕食能力强于龟纹瓢虫。前人研究发现,同龟纹瓢虫相比,异色瓢虫定殖能力更强,且具有极快的种群增长速度,可以适应复杂的环境变化,面对陌生生境中的资源结构和温度条件,仍可以保持较高的繁殖能力和生长发育速度,异色瓢虫的生态位宽度远远大于其余捕食性瓢虫,这可能是造成本研究中异色瓢虫捕食能力强于龟纹瓢虫的原因[38,39,40]。本研究结果可为番茄潜叶蛾的初期危害防控提供理论依据,在田间对番茄潜叶蛾进行防治时,可以根据其发生危害情况选择释放异色瓢虫的时期和数量进行早期防治。本研究是在室内特定条件下进行试验,即番茄潜叶蛾不是在潜叶状态下,而是被人为挑取后供两种瓢虫取食,由于潜叶类害虫如水稻潜叶蝇Hydrellia griseola、烟草潜叶蛾Phthorimaea operculella以及本研究中的番茄潜叶蛾等,该类害虫的为害方式主要为潜食叶肉,潜食后藏匿于叶片上下表皮之间,给捕食性天敌昆虫的捕食造成一定障碍,故该类害虫的目前寄生性天敌昆虫的防治研究及应用较多;部分捕食性天敌昆虫主要靠取食卵和较低龄幼虫来控制害虫数量[41,42]。故本研究在一定程度上降低了两种瓢虫取食的难度,与实际应用效果可能存在一定差异。在自然环境下,在判断两种瓢虫的控害能力时,应考虑到时间、空间、寄主植物、害虫在寄主植物上的分布以及瓢虫的饥饿程度等诸多因素[43,44]。因此,两种瓢虫在室内外不同条件下对处于潜叶状态的番茄潜叶蛾捕食能力还有待进一步研究。
