LPS对肉鸡不同组织lncRNA表达的影响
2022-09-14陈燕涛祖良鹏景梦娜张艳红
陈燕涛,赵 坤,祖良鹏,景梦娜,张艳红,郭 锋
(河南科技学院 动物科技学院,河南 新乡 453003)
随着现代基因学技术的不断发展,人们发现在哺乳动物基因组RNA中90%以上的RNA为非编码RNA (non-coding RNA),按照其长度不同可分为短链非编码RNA(short non-coding RNA,sncRNA)和长链非编码RNA(long non-coding RNA,lncRNA)[1]。lncRNA的长度大于200个核苷酸,其很长一段时间内被视作“转录噪音”。但近些年,越来越多的研究表明,lncRNA能从表观遗传学修饰、转录、前体mRNA可变剪切、mRNA稳定性及翻译等多层面对基因表达进行调控[2-3],在胚胎发育、肿瘤发生及免疫反应等生物学过程中发挥着重要作用。
先天性免疫反应是机体抵抗病原微生物感染的第一道防线。近年来研究发现,lncRNA在调节先天性免疫反应中发挥着重要作用[4]。如Du等[5]研究发现,lncRNA Mirt2能显著抑制脂多糖(LPS)诱导的小鼠外周血巨噬细胞、气管上皮细胞以及肝细胞中肿瘤坏死因子(TNF-α)、IL-1β、IL-6以及IL-12等促炎细胞因子的表达;通过尾静脉注射慢病毒,给予小鼠外源性Mirt2,能有效地缓解LPS诱导的内毒素血症。小鼠髓源性巨噬细胞中表达的lincRNA-Cox2能对多种炎症刺激物如细菌LPS和脂蛋白(Pam3CSK4)、李斯特菌以及仙台病毒等做出反应。LincRNA-Cox2能与RNA结合蛋白互作,抑制抑制趋化因子5 (chemokine 5,Ccl5)、信号转导子和转录激活子1(signal transducer and activator of transcription 1,Stat1)及Toll样受体7 (Toll-like receptor 7,Tlr7) 等免疫相关基因的表达[6]。Rapicavoli等[7]在小鼠胚胎成纤维细胞上的研究也发现,炎症刺激物TNF-α和IL-1β能诱导lncRNA Lethe的表达,Lethe通过与p65蛋白互作,抑制p65与炎症相关基因IL-6、IL-8启动子的结合,进而抑制IL-6及IL-8的表达。另外,lncRNA也可与其他调节RNA如miRNA相互作用调控炎症相关基因的表达。如lncRNA SNHG5能竞争性抑制miR-132与靶基因的结合,进而调控人气道上皮细胞中促炎细胞因子IL-1β、IL-6及TNF-a等基因的表达[8]。Zhou等[9]发现,MALAT1可通过miR-590促进STAT3介导的血管内皮细胞中促炎细胞因子IL-6、IL-8及TNF-α等的表达,促进炎症反应。
应用测序技术,在鸡肌肉、脂肪细胞、卵泡及肝脏中鉴定出大量lncRNA[10-13]。有关禽类的研究提示,lncRNA在调节脂肪代谢[14-15]、肌内脂肪细胞分化[16]、成肌细胞增殖分化[17]等生物过程中发挥着调控作用。另外,病毒感染也会引起病鸡lncRNA表达的差异。如禽白血病病毒可引起鸡胚成纤维细胞36个lncRNA差异表达,且其表达与Stat1、Tlr3、IRF1等免疫相关基因的表达水平具有相关性[18]。Han等[19]研究也发现,马立克氏病病毒感染鸡CD4+T细胞能够促进lncRNA GALMD3的表达,应用shRNA下调GALMD3的表达后,CD4+细胞中与线粒体结构以及细胞周期相关的大量基因表达受到影响。另外,禽传染性支气管炎病毒及传染性法氏囊病病毒感染都会导致病鸡相关组织和细胞中lncRNA表达谱的变化,且差异表达lncRNA与细胞内的先天性免疫反应及细胞凋亡等生物学过程相关[20-21]。上述结果提示,lncRNA在鸡先天性免疫反应中可能发挥了重要调控功能。
我国是肉鸡养殖大国,在养殖过程中,饲养环境中的细菌如大肠杆菌、沙门氏菌等,常引起肉鸡机体发生系统性炎症反应。LPS是革兰氏阴性菌细胞壁的成分,可以通过模式识别受体TLR4诱导机体产生炎症反应,而其对肉鸡各组织中lncRNA表达的影响尚不清楚。本研究以肉鸡为对象,探讨LPS诱导的肉鸡系统性炎症反应中lncRNA在肌肉、肝脏、脾脏中的表达情况,并分析其与促炎细胞因子IL-1β和IL-6基因表达的相关性,旨在为后续研究特定lncRNA在肉鸡炎症反应中的功能提供一定理论依据。
1 材料与方法
1.1 材 料
新生Ross肉鸡,购自河南鹤壁大用牧业有限公司,自由饮水、采食,按照标准饲喂条件饲养于同一环境。大肠杆菌026:B6源LPS (L2654),购自Sigma-Aldrich公司;总RNA提取试剂盒(TRIzol Reagent,15596026),购自Invitrogen;反转录试剂盒(R211-01),购自Vazyme;SYBR Green Master Mix,购自TaKaRa公司。
1.2 LPS诱导炎症模型的建立
取饲喂至21日龄的Ross肉鸡10只,称质量,随机平均分成LPS组和对照组(CK)。LPS组腹腔注射LPS,剂量为0.5 mg/kg(使用生理盐水配制LPS,质量浓度为0.2 mg/mL,依照体质量计算每只鸡的注射体积);对照组腹腔注射相应体积的生理盐水。注射后2 h采集腿肌、肝脏和脾脏,备检。
1.3 RNA提取及反转录
分别取肌肉、肝脏和脾脏,于液氮中研磨,按照TRIzol试剂盒说明书提取总RNA,用DNase Ⅰ处理消除基因组DNA污染;应用变性胶分析,确保总RNA无降解;使用紫外分光光度计测定RNA浓度。取2 μg处理后的总RNA,用反转录试剂盒进行反转录,所得cDNA于-20 ℃保存,备用。
1.4 肉鸡各组织中IL-1β和IL-6基因mRNA表达的检测
选择β-actin作为内参基因,采用实时荧光定量PCR (Real-time PCR,RT-qPCR) 检测IL-1β和IL-6的mRNA表达变化。根据GenBank上鸡的相关cDNA序列,使用Primer Premier 5.0软件设计IL-1β、IL-63和β-actin的引物(表1),引物由上海SangonBiotech公司合成。

表1 供试引物序列Table 1 Nucleotide sequences of specific primers
反应体系总体积为10 μL:10倍稀释的样品cDNA 2 μL、SYBR Green Real-time PCR Master Mix 5 μL、无菌双蒸水2 μL、目的基因上下游引物(浓度为1 μmol/L)各0.5 μL。PCR反应条件为:95 ℃预变性5 min;95 ℃15 s,60 ℃ 30 s,72 ℃ 30 s,循环36次。采用2-ΔΔCt法分析结果。以CK组为基准,确定LPS组IL-1β、IL-6基因的相对表达量。
1.5 lncRNA表达的检测
依据已报道的数据[10-11],选择肉鸡14个高表达lncRNA为备检RNA,以β-actin为内参基因,应用RT-qPCR分别检测肌肉、肝脏、脾脏中14种lncRNA的表达变化。根据GenBank上鸡的相关cDNA序列,使用Primer Premier 5.0软件设计14个lncRNA的引物(表1),引物由上海SangonBiotech公司合成。RT-qPCR反应条件及数据分析方法如1.4所述。
1.6 统计学分析
所有统计分析用SPSS 17.0 for Windows软件进行。所有数据均以“平均值±标准误”的形式表示,差异显著性检验使用独立样本t检验,相关性分析采用双变量Pearson相关性分析。P<0.05表示有显著差异,P<0.01表示有极显著差异。
2 结果与分析
2.1 LPS对肉鸡IL-1β和IL-6基因mRNA表达的影响
如图1所示,LPS组肉鸡肌肉、肝脏及脾脏中IL-1β和IL-6基因mRNA的相对表达量均极显著高于CK组(P<0.01)。
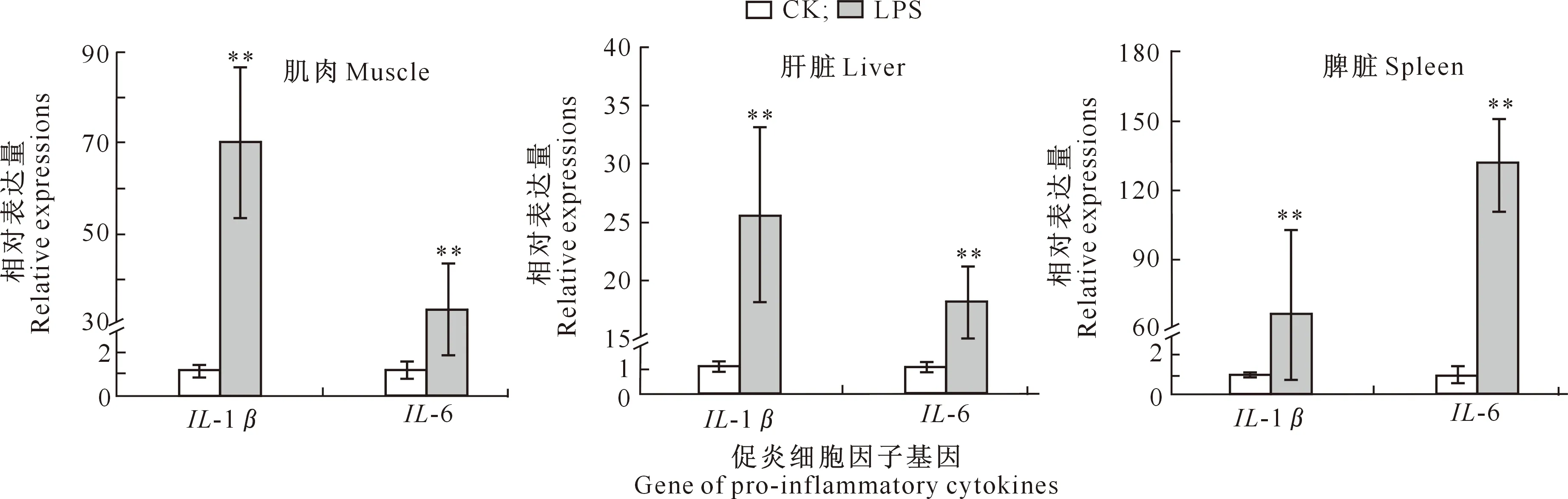
图柱上标*和**分别表示LPS组与CK组之间有显著(P<0.05)和极显著(P<0.01)差异。图2,3,4同 * and ** indicate significant (P<0.05) and extremely significant difference (P<0.01) between LPS group and CK group, respectively.The same forFigures 2,3 and 4
2.2 LPS对肉鸡不同组织中lncRNA表达的影响
2.2.1 肌 肉 检测了肌肉中14种lncRNA的表达,其中lnc0239、TCO619和TCO924未检测到;另外11种lncRNA检测结果(图2)显示,与CK组相比,LPS组lnc0035和TCO873的相对表达量极显著升高(P<0.01),lnc0181的相对表达量显著升高(P<0.05),lnc0119的相对表达量极显著下降(P<0.01),lnc0151的相对表达量显著下降(P<0.05),其他lncRNA的相对表达量无显著变化。
2.2.2 肝 脏 检测了肝脏中14种lncRNA的表达,其中lnc0239未检测到;另外13种lncRNA检测结果(图3)显示,与CK组相比,LPS组lnc0035、lnc0181、TCO354和pouBW1的相对表达量显著升高(P<0.05),lnc0202和TCO873的相对表达量极显著升高(P<0.01),lnc0151的相对表达量显著下降(P<0.05),lnc0119、TCO501和lncDHCR24的相对表达量极显著下降(P<0.01),其他lncRNA的相对表达量无显著变化。
2.2.3 脾 脏 检测了脾脏中14种lncRNA的表达,其中TCO924未检测到;另外13种lncRNA检测结果(图4)显示,与CK组相比,LPS组lnc0181、lnc0202、TCO619和TCO873的相对表达量显著升高(P<0.05),lnc0035和lncDHCR24的相对表达量显著降低(P<0.05),lnc0119和lnc0128的相对表达量极显著降低(P<0.01),其他lncRNA的相对表达量无显著变化。
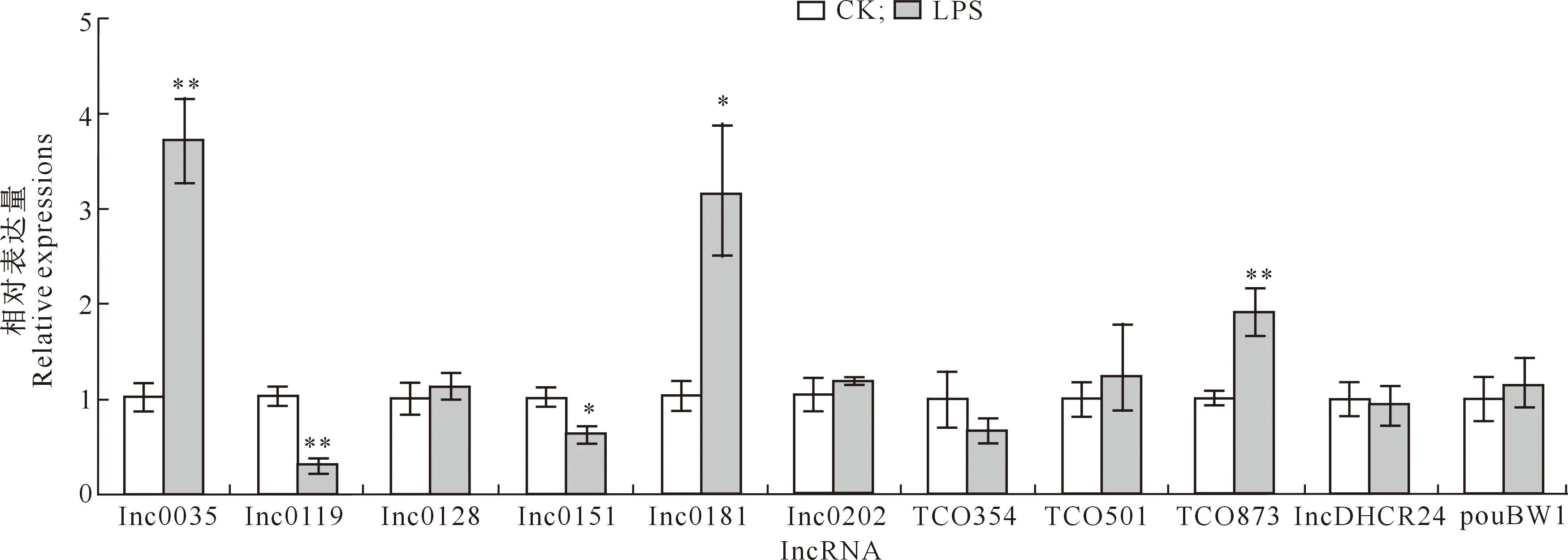
图2 LPS对肉鸡肌肉组织中lncRNA表达的影响(n=5)Fig.2 Effect of LPS on expression of lncRNAs in broiler muscle tissue (n=5)
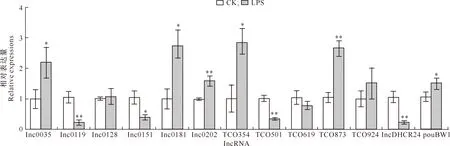
图3 LPS对肉鸡肝脏组织中lncRNA表达的影响(n=5)Fig.3 Effect of LPS on expression of lncRNAs in broiler liver tissue (n=5)
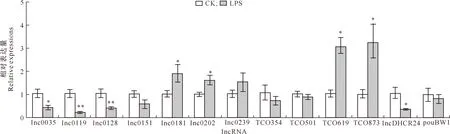
图4 LPS对肉鸡脾脏组织中lncRNA表达的影响(n=5)Fig.4 Effect of LPS on expression of lncRNAs in broiler spleen tissue (n=5)
2.3 lncRNA表达与IL-1β以及IL-6基因表达的相关性
2.2节的结果显示,与CK组相比,LPS组lnc0035、lnc0119、lnc0181、TCO873在肌肉、肝脏、脾脏中的表达水平均出现显著差异,故本试验进一步分析了其表达量与IL-1β、IL-6 mRNA表达量的相关性。结果(表2)显示,在肌肉中,除lnc0035与IL-6 mRNA表达量无显著相关性外,其他3个lncRNA均与IL-1β和IL-6的mRNA表达量显著相关,其中lnc0119与2个促炎细胞因子的表达均呈显著负相关,其他lncRNA与2个促炎细胞因子基因mRNA表达量均显著或极显著正相关。在肝脏中,lnc0035与TCO873与IL-1β和IL-6的mRNA表达量均呈显著正相关,其他lncRNA与2个促炎细胞因子基因mRNA表达量均无显著相关性。在脾脏中,lnc0035、lnc0119与IL-6 mRNA表达量分别呈显著和极显著负相关,lnc0181与IL-6 mRNA表达量呈显著正相关,TCO873与IL-1β和IL-6 mRNA表达量均呈极显著正相关。
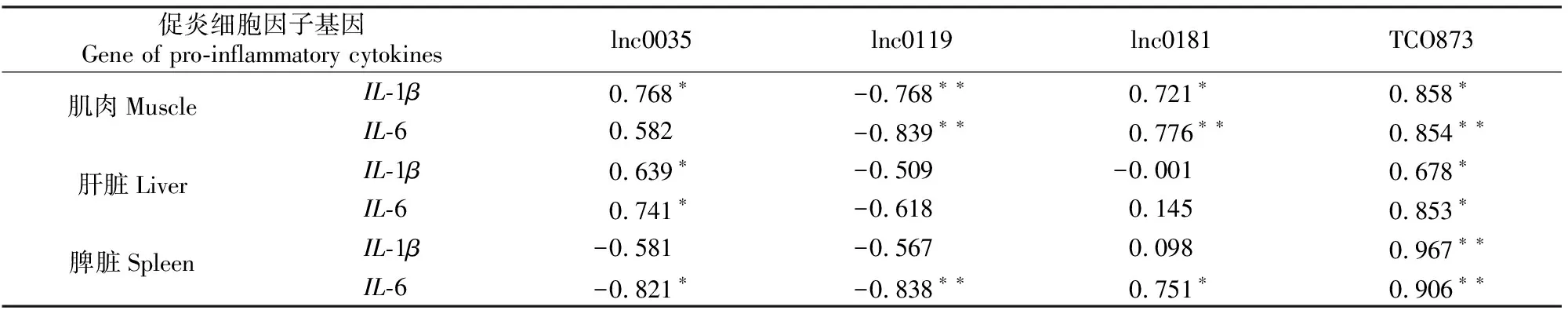
表2 肉鸡肌肉、肝脏以及脾脏中lncRNA与IL-1β、IL-6基因mRNA相对表达量的相关性Table 2 Correlation between relative expression of lncRNA with IL-1β and IL-6 mRNA in muscle,liver,and spleen of broilers
3 讨 论
LPS是革兰氏阴性菌外膜特有的成分,被广泛用于激活先天免疫,诱导机体产生系统性炎症反应。LPS可通过TLR4途径诱导细胞中促炎细胞因子大量表达,其中以IL-1β和IL-6最为典型。本试验结果显示,LPS处理2 h后肉鸡肌肉、肝脏及脾脏组织中IL-1β和IL-6基因的mRNA表达水平均极显著上调,表明LPS诱导急性系统性炎症反应的成功。促炎细胞因子IL-1β、IL-6主要是由激活的巨噬细胞以及一些非免疫细胞如成纤维细胞、内皮细胞等在受到损伤或病原入侵时大量表达[22]。作为局部炎症反应的主要刺激物,这些细胞因子能够招募其他免疫细胞到达炎症部位,对机体抵抗感染具有保护作用。但是促炎细胞因子的过度表达则可能导致组织的损伤、延迟受损组织的恢复,因此其表达须受到严格调控。
lncRNA虽不能编码蛋白,但大量的研究已证实其可作为调节性RNA (regulatory RNA)在基因转录及转录后水平发挥调控作用,且lncRNA可通过RNA-RNA、RNA-蛋白以及RNA-DNA互作等多种方式发挥作用,可正向或反向调控靶基因的表达。大量的研究表明,通过以上机制lncRNA在机体先天性免疫反应中发挥着重要作用。如小鼠lincRNA-Cox2能够促进脂蛋白(Pam3CSK4)诱导的Ccl5、IRF7、Stat3等基因表达,同时抑制IL-10、IL-1β、IL-12等基因的表达[6]。应用高通量测序技术发现,禽白血病病毒、传染性法氏囊病病毒、禽传染性支气管炎病毒以及马立克氏病病毒都能够引起鸡相应组织细胞如T细胞、成纤维细胞、巨噬细胞以及法氏囊组织中大量lncRNA表达的变化[18-21]。本研究利用LPS诱导急性炎症反应,检测肉鸡多个组织中lncRNA的表达情况,结果显示LPS组试验鸡的lnc0035、lnc0119、lnc0181和TCO873 4种lncRNA在3种组织中的表达量与CK组存在显著或极显著差异,其中lnc0181和TCO873在3个组织中表达量均显著或极显著升高;lnc0119的表达量均极显著降低;而lnc0035在肌肉和肝脏中的表达量分别极显著和显著升高,在脾脏中的表达量显著降低。另外,LPS组试验鸡的lnc0151、lnc0202、lncDHCR24表达量在2种组织中与CK组存在显著或极显著差异。这样高比例的差异表达lncRNA的出现提示,lncRNA在鸡炎症反应中可能发挥了重要调控作用。另外通过比较同一lncRNA在不同组织中的表达量可以发现,在LPS诱导的炎症反应中,lncRNA的表达具有组织差异性,提示其在不同组织中的功能可能有差异。
聚焦lncRNA在免疫反应中的调控功能的相关研究证实,lncRNA能够通过多种机制调节细胞因子的表达。如小鼠巨噬细胞中,lncRNA THRIL能够与异源核糖核蛋白L(heterogenous nuclear ribonucleoprotein L,hnRNPL)结合形成RNA-蛋白复合物,结合到TNFα启动子上促进其转录[23]。人肝癌细胞系中,lncRNA DILC能够直接结合到IL-6启动子上促进其表达[24];而在人成纤维样滑膜细胞以及软骨细胞中,DILC能够抑制IL-6的表达[25-26]。同样,lncREN MALAT1也被证实在不同的细胞中对IL-6的调控作用不同,如MALAT1在小鼠小胶质细胞和内皮细胞中能够促进IL-6的表达,而在小鼠软骨细胞中能抑制IL-6的表达[27]。本研究的相关性分析结果显示,尽管LPS组试验鸡4种lncRNA在肌肉、肝脏及脾脏中的表达量与CK组存在显著或极显著差异,但只有TCO873与IL-1β和IL-6基因mRNA的表达量在3个组织中均呈显著或极显著正相关,而其他lncRNA与两种促炎细胞因子表达的相关性都表现出组织特异性。这些结果提示,lncRNA的功能在不同的细胞及组织中具有特异性,这些lncRNA在不同组织细胞中是否直接或间接调节促炎细胞因子的表达,还需要进一步应用多种分子生物学手段来探究。
