IGF2BP2调控食管癌细胞增殖和凋亡相关基因可变剪接的机制研究*
2022-09-09冯静芳马兰英李凡周
冯静芳,唐 勇,马兰英,李凡周,韩 梅
(新疆医科大学附属肿瘤医院消化内科,乌鲁木齐 830011)
食管癌恶性程度高、预后差,是全球恶性肿瘤相关死亡的第6位主要原因。尽管食管癌的诊治水平较前有所进步,但其5年生存率仍停留在较低水平[1]。胰岛素样生长因子2 mRNA结合蛋白2(IGF2BP2)也称为IMP2/VICKZ2。广泛表达于人体多种组织器官,参与胰腺生长、发育并有刺激胰岛素分泌的作用[2]。有研究表明,在食管腺癌或鳞癌患者中,IGF2BP2 表达与短生存期相关,IGF2BP2 可能是Barrett食管和食管腺癌的一个有用的预后标记物[3]。IGF2BP2 rs1470579 A>C多态性与食管胃交界腺癌风险降低相关[4]。然而,IGF2BP2 在食管癌发生、发展中的作用机制目前尚罕见文献报道。
本研究对人食管癌细胞系ECA109中IGF2BP2 进行过表达,并进行转录组测序,通过数据分析在全基因组范围内寻找IGF2BP2 调控的差异表达基因(DEG)和可变剪接基因,探讨了IGF2BP2在食管癌中的作用机制,旨在为食管癌的诊治提供新的思路与临床依据。
1 材料与方法
1.1研究对象
在人食管癌细胞系ECA109中对IGF2BP2 进行过表达,并进行转录组测序。
1.2方法
1.2.1细胞学实验
1.2.1.1克隆和质粒构建
使用 CE Design V1.04设计Hot Fusion 的引物对。每个引物都包含基因特异性序列片段和 pIRES-hrGFP-1a 载体的17~30 bp序列。正向引物:agc ccg ggc gga tcc gaa ttc ATG ATG AAC AAG CTT TAC ATC G;反向引物:gtc atc cttg tag tcc tcg ag CTT GCT GCG CTG TGA GGC。pIRES-hrGFP-1a载体经EcoRⅠ和XhoⅠ(NEB)37 ℃消化2~3 h。用Trizol从HeLa细胞中分离总 RNA。通过寡聚dT引物将纯化的RNA转录为 cDNA,然后通过聚合酶链反应(PCR)扩增合成插入片段,通过化学转化将质粒引入大肠杆菌菌株中。
1.2.1.2细胞培养、转染与分组
ECA-109 细胞(中国普赛尔生命科技有限公司)在含有 10% 胎牛血清、100 μg/mL 链霉素、100 U/mL 的RPMI 1640 中于 37 ℃、5% CO2中培养。使用脂质体2000(英杰,美国加利福尼亚州卡尔斯巴德) 进行ECA-109细胞的质粒转染。48 h后收获转染细胞用于实时荧光定量PCR分析。实验组为IGF2BP2过表达组,对照组为空载体组。
1.2.1.3基因过表达的评估
将3-磷酸甘油醛脱氢酶用作评估IGF2BP2过表达效果的对照基因。cDNA合成通过标准程序完成,实时荧光定量qPCR在Bio-Rad S1000上用Bestar SYBR Green 实时荧光定量PCR预混液(DBI生物科学公司,上海)进行。然后使用2-ΔΔCT法将每个转录本的浓度标准化为磷酸甘油醛脱氢酶mRNA水平。通过使用 GraphPad Prism软件(加利福尼亚州圣地亚哥)与配对t检验进行比较。
1.2.2分子实验
1.2.2.1RNA的提取和测序
总RNA用RQ1脱氧核糖核酸酶处理以去除 DNA。对于每个样品,1 μg总RNA用于通过Illumina®平台的 KAPA Stranded mRNA-Seq试剂盒 (#KK8541) 制备 RNA-Seq文库。
1.2.2.2RNA-Seq原始数据的清理和比对
使用FASTX-工具包(0.0.13版)从原始测序读数中修剪接头和低质量碱基。唯一映射的读数用于基因读数计数和FPKM计算(每百万个映射片段的每千碱基转录物的片段)。
1.2.2.3DEG分析
使用R Bioconductor package edgeR筛选DEG。
1.2.2.4可变剪接分析
样品之间的可变剪接事件和受调控的可变剪接事件使用ABLas进行定义和量化。ABLas 检测10种类型的 ASE 是基于剪接点读数,包括外显子跳跃(ES)、可变的 5′剪接位点(A5SS)、可变的3′剪接位点(A3SS)等。为了评估 RBP 调节的ASE,进行了t检验以评估可变剪接(AS)事件比率变化的显著性。
1.2.2.5DEG和AS事件的实时荧光定量PCR验证
从 RNA-Seq文库制备中剩余的总RNA用于实时荧光定量PCR分析。使用M-MLV逆转录酶将 RNA 逆转录为cDNA。使用SYBR Green PCR试剂盒(Yeasen),StepOne实时荧光定量PCR系统进行实时PCR。所有基因的 RNA 表达水平都针对 GAPDH 的表达水平进行标准化。同时进行实时荧光定量PCR 检测以进行ASE验证。
1.2.2.6功能富集度分析
使用KOBAS2.0服务器识别基因本体(GO)和京都基因与基因组百科全书(KEGG)通路。超几何测试和本杰米尼-霍赫伯格罗斯福控制程序用于定义每个术语的丰富程度。
1.3统计学处理
2 结 果
2.1GF2BP2对基因表达水平的调控作用
IGF2BP2过表达后实验组IGF2BP2表达显著高于对照组,差异有统计学意义(P<0.001),见图1A;RNA-Seq检测也表明实验组IGF2BP2表达(FPKM)显著高于对照组,差异有统计学意义(P<0.001),见图1B。即实时荧光定量PCR和RNA-Seq两种方法都表明过表达效果显著。聚类分析发现过表达和对照组可以明显分开,见图1C。同时其所调控的DEG较少(共 158 个),其中上调基因81个,下调基因77个(FC ≥ 2 or ≤ 0.5,P<0.01),见图1D~E。对 IGF2BP2 过表达后显著上调、下调的基因进行GO分析均未发现所富集的信号通路。即在ECA109细胞中IGF2BP2通过转录调控下游靶基因的方式不明显。

A:实时荧光定量PCR定量IGF2BP2表达;B:通过RNA-Seq数据量化的IGF2BP2表达;C:通过主成分分析进行聚类分析;D:IGF2BP2调控基因的鉴定;E:对照和IGF2BP2 过表达样品中 DEG 的分层聚类;FPKM 值是 log2 转换的,然后每个基因以中值为中心;a: P<0.001;在火山图中,上调基因用红色标记,下调基因用蓝色标记。
2.2IGF2BP2调节数百个基因的选择性剪接
IGF2BP2调控的可变剪接事件的分类,见表1,图2A。对各样本中发生差异可变剪接的基因(RASG)和DEG进行整合分析,得到表达水平和可变剪接水平都发生显著差异的基因共4个,见图2B。IGF2BP2过表达可变剪接水平发生显著变化的基因,在GO分析中富集到糖基磷脂酰肌醇(GPI)锚定生物合成过程、蛋白质稳定性调节、翻译后的蛋白质修饰、DNA 修复、有丝分裂、基因表达、凋亡过程的正向调控、蛋白c端脂肪化作用、核转录mRNA聚(A)尾缩短、细胞增殖等信号通路,其中与食管癌明显相关的是细胞增殖和细胞凋亡过程,见图2C。IGF2BP2 过表达后可变剪接水平发生显著变化的基因,在 KEGG 分析中富集到代谢途径、赖氨酸退化、嘧啶代谢、类固醇生物合成、GPI锚的生物合成、叶酸生物合成、细胞周期、过氧化物酶体增殖物激活受体(PPAR)信号通路、烟酸和烟酰胺代谢、甘油磷脂新陈代谢等信号通路,见图2D。
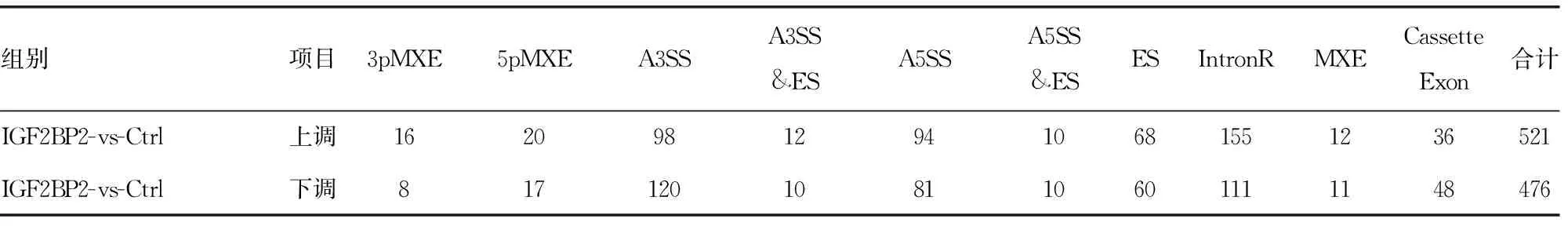
表1 差异可变剪接事件统计

A:IGF2BP2调控的可变剪接事件的分类;B:IGF2BP2调控的DEG和RASG之间的重叠分析;C:IGF2BP2的前10个富集GO生物过程调节交替剪接基因;D:IGF2BP2 的前10条富集KEGG通路调节交替剪接基因。
2.3在细胞增殖和凋亡中具有丰富意义的选择性剪接基因
IGF2BP2过表达后21个在细胞增殖方面可变剪接的基因,包括ADAMTSL4、TRIO、PLEKHG2、RBCK1、非受体酪氨酸激酶(ABL1)、FAM162A、ITSN1、ARHGAP4、tnfrsf12a等。其中IGF2BP2调控ADMTSL4基因的可变剪接,IGV-sashimi图显示A3SS事件,见图3A。IGF2BP2调控基因TRIO的可变剪接,见图3B;IGV-显示ES事件的生鱼片图。IGF2BP2调节基因ABL1、TNFRSF12A、FAM162A、ITSN1、PLEKHG2、RBCK1和的可变剪接,见图3C。

A:IGF2BP2 调控 ADMTSL4 基因的可变剪接,IGV-sashimi plot图显示了一个A3SS事件;B:IGF2BP2 调节基因 TRIO 的可变剪接,IGV-sashimiplot 显示ES 事件;C:IGF2BP2 调节基因 ABL1、TNFRSF12A、FAM162A、ITSN1、PLEKHG2、RBCK1、TNFRSF12A 的可变剪接。
寻找在细胞凋亡中具有丰富意义的可变剪接基因发现23个在IGF2BP2过表达后调控细胞凋亡的可变剪接基因,包括细胞周期蛋白依赖性激酶11B(CDK11B)、制瘤素M的细胞因子受体(OSMR)、MRE11A、母体胚胎亮氨酸拉链激酶(MELK)、FTSJ2等。其中IGF2BP2调控基因CDK11B的可变剪接,IGV-生鱼片图显示A5SS和ES事件,见图4A。IGF2BP2调控基因MRE11A的可变剪接,IGV-sashimi plot图显示A5SS事件,见图4B。 IGF2BP2调控基因FTSJ2、MELK、OSMR的可变剪接,见图4C。

A:IGF2BP2调控基因CDK11B的可变剪接,IGV-sashimi plot图显示A5SS和 ES 事件;B:IGF2BP2调节基因MRE11A的可变剪接,IGV-sashimi plot图显示A5SS事件;C:IGF2BP2调节基因FTSJ2、MELK、OSMR 的可变剪接;在A和B中,每个可变剪接事件的读数分布绘制在左侧面板中,每个基因的转录本如下所示:示意图描绘了ASE、AS1(紫线)和 AS2(绿线)的结构;组成型外显子序列用黑框表示,内含子序列用水平线表示(右图,顶部),而替代外显子用红色框表示,内含子用紫色框表示;ASE的RNA-Seq定量和实时荧光定量PCR验证显示在右侧面板的底部;a:P<0.05。
3 讨 论
本研究通过RNA-Seq获得了人类食管癌细胞系ECA109中IGF2BP2调控的转录组。比较转录组分析显示,IGF2BP2过表达导致ECA109细胞中158个基因发生显著差异表达,其中81个上调,77个下调。然后,对 IGF2BP2 过表达后显著上调表达的基因进行GO分析发现没有所富集的信号通路,其原因可能是 IGF2BP2过表达上调的基因较少,无法富集到相关的功能通路。对 IGF2BP2 过表达后显著下调的基因进行 GO 分析也没有发现所富集的信号通路,其原因与上调类似,可能是 IGF2BP2 过表达下调的基因较少,无法富集到相关的功能通路。因此,在食管癌中IGF2BP2 通过调控DEG的方式影响食管癌的发生、发展,可能并不显著。另外,IGF2BP2过表达造成997个差异可变剪接事件的发生,在 GO 分析中富集到凋亡过程的正向调控,细胞增殖与食管癌相关。在KEGG分析中同样富集到细胞凋亡及增殖与食管癌相关。提示IGF2BP2通过调控基因的可变剪接,继而参与以上生物过程的发生。这些发现支持了IGF2BP2 作为潜在的食管癌促癌因子可能通过细胞增殖、凋亡相关通路中的基因的可变剪接促进食管癌的发生、发展,进一步扩展了对其作为临床治疗靶标的作用机制的认识。
在IGF2BP2调控的可变剪接事件中,在细胞增殖功能通路中发现ADAMTSL4、TRIO、PLEKHG2、RBCK1、ABL1、FAM162A、ITSN1、TNFRSF12A的可变剪接发生了显著变化。有研究表明,ADAMTSL4作为细胞外基质相关特征的基因,通过其表达和免疫功能分析发现这种 ECM 相关的特征基因可能在肿瘤微环境的变化中发挥着重要作用,可能作为独立的预后因素,并为食管鳞癌(ESCC)患者的化疗反应提供潜在的生物标志物[5]。TRIO基因位于染色体5p扩增区,其基因融合已经在4种不同的肉瘤组织类型中被识别出来,可能意味着与肿瘤进展有关的次生事件有关[6]。ABL1被认为是ESCC的预测和预后因素,可以反映肿瘤微环境中的免疫紊乱并独立区分生存率降低的高风险患者,为ESCC患者提供新的预测和治疗靶点[7]。其还可与上述的另一基因PLEKHG2,一种Rho特异性鸟嘌呤核苷酸交换因子,相互作用通过核因子-κB 信号通路通过细胞内蛋白质积累抑制细胞生长[8]。已有研究表明,TNFRSF12A是一种可行的预后生物标志物,也是胶质瘤的潜在免疫治疗靶点,其表达水平也与乳腺癌、甲状腺癌等患者的预后相关,但目前暂无与食管癌直接的相关研究[9-11]。上述基因均已证实与食管癌,抑或肿瘤的发生等相关,与本研究结果类似,但其具体作用是否与IGF2BP2过表达后所致基因在细胞增殖方面的可变剪接有关尚有待于进一步研究。而RBCK1、FAM162A、ITSN1基因在食管癌及其他恶性肿瘤中的研究目前尚未见文献报道,但可能通过其可变剪接参与食管癌的发生、发展,尚有待于进一步研究验证。
在IGF2BP2调控的可变剪接事件中,在细胞凋亡功能通路中发现CDK11B、OSMR、MRE11A、MELK、FTSJ2的可变剪接发生了显著变化。有研究表明,减数分裂重组11同源物与ESCC 肿瘤耐药的异常机制相关。抑制 MRE11A 表达与 ESCC 细胞系中的顺铂耐药性相关。MRE11A可以作为新辅助化疗(NAC)反应的预测因子和 ESCC 患者的预后标志物[12]。MELK是 ESCC 患者的潜在治疗靶点,在 ESCC 细胞系和人类样本中观察到 MELK 的高表达,尤其是在转移性肿瘤组织中。MELK过表达促进了ESCC细胞增殖、集落形成、迁移和侵袭,MELK的增强表达大大加速了体内ESCC细胞的肿瘤生长和肺转移[13]。FTSJ2是一种热休克诱导的线粒体蛋白,对癌细胞的侵袭和迁移具有抑制作用[14]。也有文献报道称FTSJ2可以降低了非小细胞癌细胞的侵袭性,可能是非小细胞癌患者治疗反应的预测性生物标志物[15]。OSMR被认为与多种癌症相关。如OSMR被确定为具有较高表达水平、显著预后价值和优良诊断效率的胰腺癌关键基因[16]。另一研究指出OSMR调节脑肿瘤干细胞增殖和胶质母细胞瘤的发生,抑制OSMR可改善胶质母细胞瘤对电离辐射的反应并延长寿命[17]。但目前没有相关文献报道其在食管癌中的作用机制,包括CDK11B,也暂未见其与食管癌相关性的文献报道,然而有文献报道称其在肝细胞癌 (HCC) 的生物学中起着至关重要的作用,CDK11B可促进HCC干细胞的自我更新,敲除CDK11B可以减弱HCC干细胞的自我更新能力及其体内致癌性[18]。虽然前述研究已证明CDK11B、OSMR、MRE11A、MELK、FTSJ2基因在肿瘤等疾病中的作用,其中不乏与食管癌发生、发展相关的基因,通过本研究,进一步证明了其与食管癌的关系,即可能是发生在IGF2BP2过表达后调控细胞凋亡的可变剪接基础之上。
综上所述,本研究通过在人食管癌细胞系ECA109细胞中对IGF2BP2进行过表达发现IGF2BP2过表达造成997个差异可变剪接事件的发生,经过GO和KEGG分析富集到细胞增殖和凋亡功能通路。IGF2BP2可能通过调控细胞增殖和凋亡相关基因可变剪接的方式从而在食管癌中发挥作用。在后期的研究中将继续对IGF2BP2调控的可变剪接事件进行验证,以期为食管癌诊治及分子标记物的研究提供更多的理论依据。
