野菊舌状花形态变异的遗传分析
2022-09-07任江珊顾雪琪牛雅静付海蓝戴思兰
任江珊, 顾雪琪, 牛雅静,2, 蒲 娅, 付海蓝, 黄 河, 戴思兰
(1.北京林业大学园林学院/花卉种质资源创新与分子育种北京市重点实验室/国家花卉工程技术研究中心/城乡生态环境北京实验室,北京 100083; 2.北京市植物园管理处/北京市花卉园艺工程技术研究中心,北京 100093)
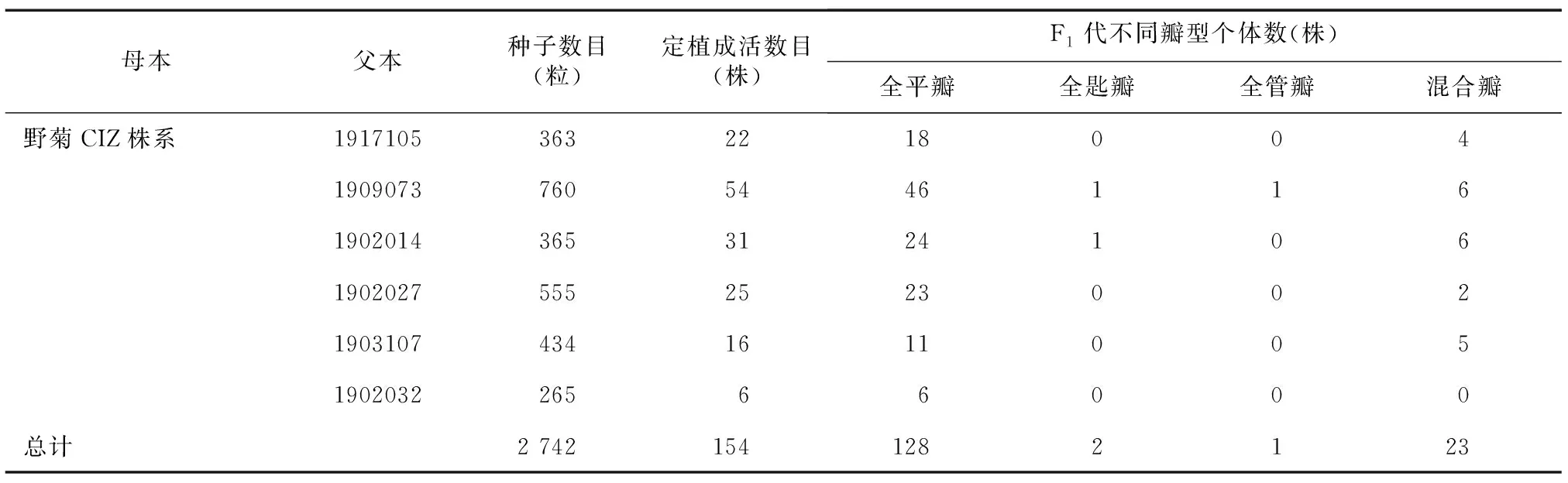
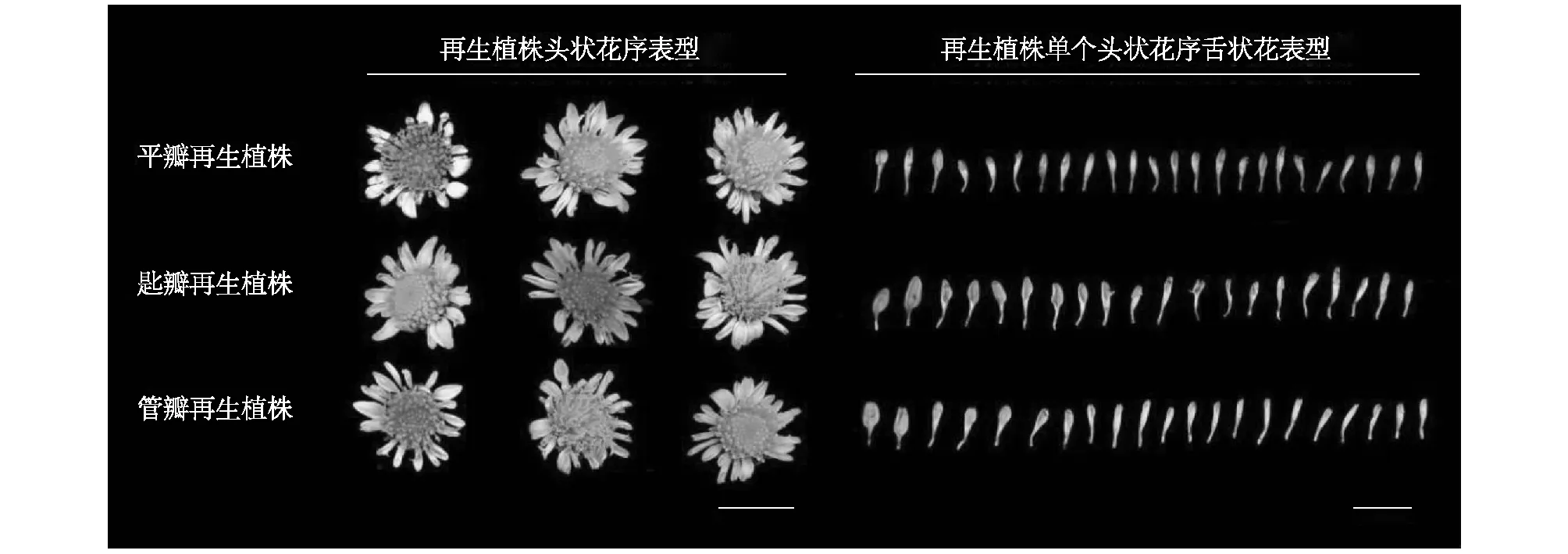
菊花(Chrysanthemum×morifolium)是中国十大传统名花之一[1],其丰富多样的花型与瓣型吸引着全世界园艺学研究者的关注。菊花的花型主要依据头状花序的形态,即中央的管状花和周围的舌状花通过不同的瓣型、数量、姿态排列组合,从而形成不同的花型[2]。瓣型是菊花花型分类的第一级标准[3],根据舌状花形态可分为平瓣、匙瓣、管瓣3种基本瓣型,以及管状花花冠筒伸长并先端开裂成星芒状的桂瓣瓣型,舌状花尖端龙爪状或剪绒状开裂,或着生毛刺等附属物的畸瓣瓣型[1,4]。花冠筒长度在菊花DUS测试中是一个重要指标[5]。Song等[6]将花冠筒长(CTL)与舌状花长(RFL)的比值定义为花冠筒基部合生程度(CTMD),依次划定3种不同舌状花瓣型,0≤CTMD≤0.2为平瓣型,0.2 舌状花形态是影响菊花观赏性的重要因素,研究其遗传规律对于菊花瓣型改良育种十分重要。长久以来,菊花各瓣型的来源及其演变关系存在争议,相关遗传机制尚不明确。陈俊愉[8]认为菊花舌状花的演化,是从平瓣演化出匙瓣,再成为管瓣,同时舌状花数量增加。然而在野生菊和栽培菊中,均发现平瓣、匙瓣、管瓣共生于同一个花序上的现象,由此推测菊花各种瓣型之间不存在相互演化的关系[3,9]。前人通过对不同组合的亲本及其F1代性状进行统计分析,发现菊花瓣型存在平瓣遗传力更强和偏母性遗传等现象[10]。陈云志等[11]的试验结果也表明在分离的后代中,平瓣出现的概率最大,其次为匙瓣,管瓣出现的概率最小;夏伯顺[12]发现瓣型性状表现出明显的偏母性遗传,后代瓣型与母本瓣型相同的比例达80%以上;王希等[13]的试验结果同样表明菊花瓣型遗传具有偏母性遗传特点,但也依亲本的瓣型而定,不同瓣型的遗传力不同。 菊属植物种间生殖隔离不完全,种间杂交较为容易[14],因此现代菊花作为栽培杂种复合体,具有复杂的染色体倍性及起源演化关系,这使得以菊花为材料进行舌状花遗传规律的研究十分困难。野菊(Chrysanthemumindicum)是菊科(Asteraceae)菊属(Chrysanthemum)多年生草本植物,其为菊属四倍体野生植物资源,在中国分布广泛。前人研究结果表明,野菊是现代菊花起源的重要亲本[15-18],参与形成了如今庞大的栽培菊花杂种。课题组前期在野菊分布区域内收集到一株野菊瓣型突变株,不同于舌状花全部为平瓣的野菊野生型,其舌状花自然突变为3种瓣型,平瓣、匙瓣、管瓣并生于一个头状花序,且通过扦插繁殖该变异能够稳定存在,可以作为菊花舌状花形态研究及遗传规律研究的重要模式植物,我们将其命名为CIZ株系(图1)。此外,不同瓣型的舌状花并存于同一头状花序,是一种类似嵌合性状的表现。植物嵌合体是指遗传上不同的2种植物的组织机械地共存于同一个生长点的植物[19]。目前国内外对菊花嵌合体的遗传特性报道较少[20-21],CIZ株系是否为嵌合体,其3种瓣型是在花发育前期还是花器官成熟时期形成的尚不明确。 本研究构建了野菊CIZ株系和野生型株系CIW的F1代杂交群体,通过统计后代舌状花花冠筒基部合生程度以探究舌状花形态变异遗传规律;同时通过野菊CIZ株系与平瓣瓣型小菊杂交探究瓣型突变性状的遗传性及野菊CIZ株系作为菊花瓣型改良育种材料的可行性。另一方面,为了探究野菊CIZ株系瓣型嵌合性状在无性繁殖中的稳定性,分别以不同瓣型舌状花离体培养,观察不同瓣型舌状花外植体再生植株瓣型。野菊瓣型变异规律的研究,对菊花瓣型遗传机制的研究及菊花瓣型改良育种具有重要意义。 通过野外收集和扦插繁殖获得2个野菊株系,野菊CIW株系表现为舌状花全部为平瓣(图1A);野菊CIZ株系舌状花自然突变为3种瓣型(图1B),即表现为平瓣、匙瓣、管瓣并存于一个头状花序。 A:野菊CIW株系;B:野菊CIZ株系。图1 野菊及其瓣型突变株Fig.1 Chrysanthemum indicum and strains of petal type mutation 为探究野菊CIZ株系是否可以作为菊花瓣型改良的育种材料,从课题组资源圃中挑选6个株型紧凑的平瓣园林栽培小菊品种(1902014、1909073、1902027、1902032、1903107、1917105),它们都具有花量大、花径大等优良观赏性状。将6个小菊品种与野菊CIZ株系进行杂交试验,探究瓣型突变性状在有性繁殖中的遗传稳定性,并为丰富盆栽小菊瓣型性状提供创新种质。 1.2.1 杂交群体构建 以野菊CIW株系为母本、野菊CIZ株系为父本进行人工杂交。在母本花蕾初开时去除内轮管状花,同时剪去舌状花花瓣直至柱头可见,用硫酸纸袋套袋,同时收集父本花粉。待母本柱头伸出开叉并分泌黏液时,于晴天的中午对母本进行授粉、套袋,次日重复授粉。冬季采集干燥的人工授粉的花序,脱粒,获得F1代杂交种子,次年播种。观察记录杂交后代瓣型性状分离情况,分析遗传规律。以野菊CIZ株系为母本,具有优良性状的6个平瓣小菊品种为父本,杂交方法同上,得到F1代杂交后代。 1.2.2 F1代舌状花数据采集与分析 对野菊CIW株系与CIZ株系F1代群体各个单株进行数据采集,每个单株随机选取3个花序,测量花序上每个舌状花的花冠筒长和舌状花长,并根据花冠筒长/舌状花长计算CTMD。统计杂交F1代CTMD测量结果的最小值(Min)、最大值(Max)、极差(R)、平均值(x)、标准差(S)等,以0.1倍于极差的组距分级,将CTMD值划分为10组,统计各级的频数,绘制次数分布频次图,并利用非参数检验中的单样本K-S法检验CTMD这一性状是否符合正态分布。 统计野菊CIZ株系与6个平瓣小菊品种杂交后代F1代种子数目、定植成活数目等,并对杂交F1代进行瓣型统计。 1.2.3 杂种优势分析及显著性检验 以中亲优势和中亲优势率表示杂种优势,将杂交F1代CTMD的平均值(Fm)与中亲值(Mid-parents value,MPV)之差定义为中亲优势(Hm):Hm=Fm-MPV,将 (Fm-MPV)/MPV×100%定义为中亲优势率(RHm),RHm>0为正向优势,RHm<0为负向优势。其中,中亲值为双亲某一性状的平均值。 超亲优势(Hb)为杂交F1代CTMD平均值(Fm)与高值亲本表型值(HP)之差,Hb=Fm-HP,超亲优势率(RHb)=(Fm-HP)/HP×100%,RHb>0说明F1代表型值偏向于高值亲本,RHb<0说明F1代表型值偏向于低值亲本。采用Excel 2016和SPSS 23.0进行数据的统计分析。 1.2.4 不同瓣型舌状花的离体培养及再生植株性状观察 植物组织培养试验在北京林业大学园林学院组培室完成。将野菊CIZ株系3种瓣型的舌状花消毒处理后,接种在以MS为基础培养基、含有2 mg/L细胞分裂素(6-BA)与2 mg/L萘乙酸(NAA)的花瓣再生培养基上,待再生芽长至1.5 cm左右时,将其转移到以1/2 MS为基础培养基、含有0.2 mg/L NAA的生根培养基上;组培苗生根40 d时,选择根系发达且生长健壮的组培苗,拿出并洗净根部的残余培养基,将其移栽至提前高温灭菌的基质(营养土∶蛭石∶珍珠岩=1∶1∶1,体积比)中,放至人工气候室内培养,移栽后在长日照条件(16 h光照/8 h黑暗)下培养70 d后,转入短日照条件(12 h光照/12 h黑暗)下诱导花芽分化培养,植株开花后对其瓣型性状进行观察和统计,针对不同瓣型再生植株分别随机选取3个花序,测量花序上每个舌状花的花冠筒长和舌状花长,并根据花冠筒长/舌状花长计算CTMD值。 2.1.1 杂交群体F1代瓣型性状表型分布 以野菊CIW株系为母本,野菊CIZ株系为父本,构建F1代杂交群体,共得到F1代种子1 044粒,出苗243株,出苗率23.28%,成苗227株,成苗率93.42%。 开花后统计227株杂交后代瓣型(图2)。统计结果显示,在杂交群体中,舌状花全表现为平瓣的有166株,约占杂交后代群体的73.13%;舌状花表现为混合瓣的有59株,约占杂交后代群体的25.99%;杂交后代中,出现了2株全管瓣,约占杂交后代群体的0.88%。 图A为部分子代表型图片;图B为典型平瓣子代、匙瓣子代、管瓣子代。图2 野菊CIW和CIZ及其部分杂交后代的花部性状Fig.2 Floral traits of Chrysanthemum indicum CIW, CIZ and part of their hybrid offsprings 2.1.2 F1代瓣型性状的描述性统计及杂种优势表现 野菊杂交F1代CTMD表型特征值显示,后代CTMD最大值为0.95,最小值为0,极差为0.95,平均值为0.06,标准差为0.13。由野菊杂交F1代CTMD频率分布图(图3)可知,F1代群体大部分为平瓣,由正态性检验结果可知,野菊杂交F1代CTMD不符合正态性分布,而是表现为偏向于低值亲本的偏态分布。说明后代舌状花表型出现较大分离,为菊花舌状花性状进一步的遗传分析提供了较好的遗传差异基础,但也可能是由于平瓣子代所占比例过大,平均值接近0。以野菊CIW平瓣株系为母本,CIZ混合瓣株系为父本,F1代群体中出现2株全管瓣的超亲个体,但由于大部分子代仍为平瓣,所以并未形成明显的超亲优势。 CTMD:花冠筒基部合生程度。图3 野菊CIW株系(♀)×野菊CIZ株系代群体CTMD频率分布直方图Fig.3 Frequency distribution histogram of CTMD derived from the F1 generation of CIW strain (♀) and CIZ strain of Chrysanthemum indicum 以野菊CIZ株系为母本,课题组资源圃6个平瓣小菊品种(1902014、1909073、1902027、1902032、1903107、1917105)为父本进行杂交,统计F1代种子数目、定植成活数目等。经统计,6个杂交组合共获得F1代种子2 742粒,发芽180粒,发芽率6.56%;成苗154株,成苗率85.56%(表1)。不同杂交组合收获的种子数和发芽情况不同,其中CIZ×1902014成苗率最高,其次是CIZ×1909073组合。 表1 杂交后代情况统计 对野菊CIZ株系与平瓣小菊杂交F1代进行瓣型统计。结果显示,杂交后代瓣型全部为平瓣的有128株,占比83.12%;混合瓣有23株,占比14.94%;此外还出现超亲子代3株,分别为全匙瓣2株,全管瓣1株。野菊CIZ株系与平瓣小菊杂交F1代中出现了瓣型变异与性状分离(图4),表明野菊CIZ株系舌状花突变性状具有遗传性,可作为园林小菊瓣型育种的重要亲本。 图为部分子代表型图片,A~J父本分别为1902014、1909073、1917105、1903107、1902014、1902032、1917105、1902027、1917105。图4 野菊CIZ株系与平瓣小菊杂交后代表型Fig.4 Phenotype of hybrid offsprings derived from Chrysanthemum indicum and flat petaled chrysanthemum 为了探究野菊CIZ株系瓣型嵌合性状在无性繁殖中的稳定性,以不同瓣型舌状花为外植体,通过诱导愈伤组织、分化丛生芽、生根培养、壮苗移栽等步骤,并在人工气候室内栽培及用短日照处理诱导开花。平瓣、匙瓣、管瓣类瓣型分别获得4株再生植株,随机选择再生植株的3朵花序,测量花序上每个舌状花的花冠筒长和舌状花长,并计算CTMD。统计不同舌状花再生植株的平均CTMD。 观察再生植株盛花期发现,平瓣、匙瓣、管瓣3种瓣型的再生植株开花后均表现为混合瓣型,保持原来的嵌合性状,未实现性状的分离(图5),不同瓣型再生植株平均CTMD无显著差异。其中匙瓣再生植株平均CTMD相对较小,为0.29,平瓣与管瓣外植体再生植株平均CTMD基本相同,分别为0.35和0.34。 图5 野菊花瓣再生苗瓣型表型Fig.5 Phenotype of regenerated plants from petals of Chrysanthemum indicum with different petal types 菊花是中国十大传统名花之一,经过长期自然变异和人工选育,现代菊花形成了丰富多样的品种类型,而不同菊花品种瓣型的形态多样性更是有目共睹。菊花起源较早且育种历史较为漫长,有证据显示,在栽培菊花的形成过程中,毛华菊和野菊资源扮演了十分重要的角色[15]。前人曾针对毛华菊花朵形态变异进行整理分析,发现毛华菊花朵变异类型与栽培菊花十分相似[22],但同样作为菊花起源的重要亲本,关于野菊的研究则较少。本研究所关注的野菊CIZ株系,瓣型特殊,在同一个花头上同时具有平瓣、匙瓣、管瓣3种瓣型,是菊花瓣型研究的重要材料。因而有助于针对野菊CIZ株系的研究,进一步解开菊花瓣型多样之谜,探究菊花平瓣、匙瓣、管瓣的演化关系,同时也为园林应用菊花瓣型的改良及定向育种提供参考。 在前人对菊花花部器官性状的研究中,发现花径、舌状花长等性状均表现为连续性较好的正态分布趋势,符合多基因控制的数量性状的基本特征[23-24]。然而,本研究中野菊CIW株系与CIZ株系杂交后代菊花花部性状CTMD集中在0.20以下,后代正态性检验结果表现为偏向于低值亲本的偏态分布。这与Song[7]等关于CTMD基本符合正态分布的结论有所不同。Song等[7]所选亲本为栽培群体,而本研究中的野菊为受人工选择影响相对较小的野生种。菊花不同的杂交组合间,花部性状杂种优势的表现程度不同[23-25],同时由于菊花基因组高度杂合且不同瓣型的遗传能力存在差异,使得杂交 F1代性状发生广泛分离,从而导致F1代杂种优势衰退[12,25-27]。另一方面,以混合瓣型野菊CIZ株系为母本,平瓣小菊为父本,杂交后代也表现为瓣型以平瓣为主,这与夏伯顺[12]得出的瓣型性状偏母性遗传的结论不符。而Song 等[7]研究发现,在试验的平瓣(母本)和管瓣(父本)杂交组合中,后代中主要为匙瓣,因而考虑野菊CIZ株系的混合瓣型可能为中间性状,其遗传力不如平瓣。综合2个杂交后代群体,进一步验证了前人提出的在分离的后代中,平瓣出现的概率最大的结论[11],相比匙瓣与管瓣,平瓣遗传力更强。 杂交是形成物种遗传多样性的原因之一[28-30]。本研究中,在以混合瓣型的野菊CIZ株系作为母本,平瓣小菊作为父本的杂交后代中,出现了瓣型性状的分离,杂交后代出现全平瓣、全匙瓣、全管瓣及混合瓣等表型,考虑到野菊还具有抗逆性强等优良性状,这进一步证明了野菊CIZ株系作为瓣型突变体在菊花瓣型育种中的价值。 野菊CIZ株系平瓣、匙瓣、管瓣3种瓣型共存于同一头状花序,是一种类似于嵌合性状的表现。目前针对菊花嵌合体的研究主要集中在花色研究方面,花色嵌合体植株不同颜色花瓣再生植株往往不能保持原本的嵌合性状,出现性状分离[31-33]。而本研究对野菊花瓣再生植株进行观察发现,平瓣、匙瓣、管瓣3种瓣型的再生植株开花后均表现为混合瓣型,保持原来的性状。Thangmanee等[34]对菊花花瓣外植体愈伤组织的组织学研究结果表明,由菊花舌状花诱导的再生芽为单一起源,来自于单个分生组织细胞。Pu等[35]通过对毛华菊舌状花形态学观察分析发现,平瓣型舌状花与管状型舌状花的形态差异出现在花瓣细胞分裂旺盛的花发育早期。结合本研究中组织培养中嵌合性状的保持现象,推测野菊CIZ株系的瓣型变异可能发生于花发育早期。 菊花瓣型是十分重要的观赏性状,而受制于菊花瓣型多样、基因型高度杂合及染色体倍性复杂等因素,对菊花瓣型遗传规律的研究及不同瓣型性状的定向育种显得尤为困难。而野菊作为现代菊花起源的重要亲本之一,对其瓣型遗传规律的研究有利于更好地解释现代菊花的瓣型起源,同时也为菊花的瓣型改良提供参考。1 材料与方法
1.1 试验材料

1.2 试验方法
2 结果与分析
2.1 野菊CIW株系与CIZ株系F1代瓣型性状遗传分析


2.2 野菊CIZ株系与平瓣小菊品种F1代瓣型性状遗传分析

2.3 野菊突变体花瓣再生植株的瓣型表型
3 讨 论
