长江中下游升金湖湿地越冬鸭属(Anas)鸟类群落结构和行为特征*
2022-09-05范少军周立志
范少军,周立志,于 超
(1:安徽大学资源与环境工程学院, 合肥 230601) (2:湿地生态保护与修复安徽省重点实验室, 合肥 230601) (3:安徽升金湖湿地生态学国家长期科研基地,东至 247230) (4:黄山学院生命与环境科学学院, 黄山 245041)
越冬期是迁徙水鸟生活史的重要阶段[1-2],水鸟在越冬期依赖各类湿地栖息地,常成群聚集形成水鸟群落[3-6]. 群落结构和行为特征是水鸟越冬生态学研究的核心. 水鸟的群落结构,包括种类组成、数量、密度和多样性等常在不同湿地之间呈现差异[4,7-11]. 行为时间分配是水鸟行为特征的直接体现,水鸟通过权衡栖息地的质量来调整行为时间分配[12-16],其中觅食和警戒行为的调整是水鸟对环境变化最直观的响应,觅食行为决定了水鸟的生存和种群适合度,警戒行为则是水鸟对人类活动威胁的直接响应[17-18]. 不同湿地的面积、水深、水生植被特征和人类干扰等环境因素影响湿地中水鸟的群落结构和行为特征[4-7]. 面积大小直接决定湿地对水鸟的容纳量,水深决定湿地的可利用性,而人类活动带来的干扰则是长江中下游地区各类湿地的重要特征. 水生植被是湿地的重要组成部分,能够维持湿地生态系统的稳定,为各种水鸟提供食物资源和隐蔽环境,水鸟往往利用水生植被丰富的湿地进行觅食或躲避风险[19-24].
升金湖是长江中下游地区具有代表性的浅水通江湖泊,是该地区主要的越冬水鸟聚集区之一. 升金湖湿地类型多样,包括浅水、泥滩、草滩和芦苇湿地等. 经过多年的湖泊生境人为改造和近些年的湿地生态修复,升金湖形成了稻田、养殖塘、退耕还湿和水生植被恢复湿地等多种人为改造的湿地类型,这些湿地在冬季可为大量水鸟提供栖息地[23,25]. 鸭属鸟类分布广、数量多,高度依赖各类湿地栖息,对人类活动和环境变化极其敏感,是湿地生态系统的重要指示类群. 升金湖是鸭属鸟类重要的越冬地,越冬群体数量最高超过10000只,但近年来呈大幅下降趋势[23,26]. 人类活动的增强导致升金湖湿地景观破碎化、斑块化,越冬水鸟生境质量变差,食物资源匮乏和隐蔽环境减少,迫使水鸟选择稻田、养殖塘等湿地类型作为觅食或躲避风险的栖息地[19-20,24]. 在破碎化和斑块化的湖泊湿地中,研究水鸟如何调整其群落结构和行为特征以适应各种类型的湿地栖息地,对于理解水鸟越冬生活史特征和各类湿地的管理具有重要意义.
本研究选择自然芦苇湿地和芡实塘、退耕还湿及水生植被恢复湿地4种湿地类型为研究样地,研究升金湖越冬鸭属鸟类的群落结构,并对优势种的行为特征进行研究,分析影响其栖息地利用的环境因素,研究旨在为加强鸭属鸟类的保护和其越冬栖息地的管理提供理论依据.
1 研究区域和方法
1.1 研究区域概况
升金湖国家级自然保护区(30°15′~30°30′N,116°55′~117°15′E)位于长江下游地区的长江南岸(图1),通过黄湓闸与长江连通,湖区位于中国东部季风气候区,7-9月份为丰水期,11月-次年5月为枯水期. 秋冬季节水位的降低使湖区形成大面积的湿地,为冬候鸟提供了食物资源和隐蔽条件,保护区内存在多种湿地类型为冬候鸟提供适宜的栖息地[5,16,25]. 升金湖自1995年以来,长期进行了大范围的水产养殖活动,大量投放草食性的鱼苗、蟹苗,导致湖区沉水植物资源严重退化. 近年来,升金湖保护区陆续实施了圩口水生植被恢复、退耕还湿及生态渔业示范项目[19].
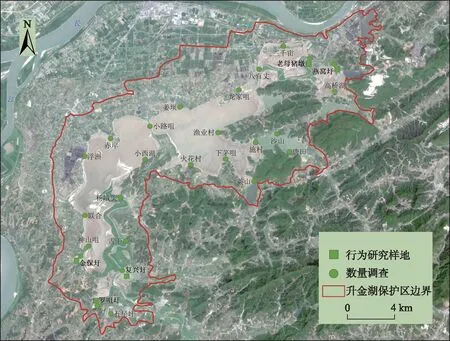
图1 研究地点Fig.1 The study sites
1.2 数据收集
2017年11月-2018年3月,在升金湖选择26个调查样点,对升金湖越冬鸭属鸟类的种类和数量分布情况进行调查,每个月的月初对各样点集中进行一轮调查,每轮调查持续3 d. 样点调查采用直接计数法,调查时避开恶劣天气,调查时间段为7:00-10:59和15:00-17:59,各样点调查时间10~15 min,借助SWAROVSKI ((20~60)×80)单筒望远镜记录湖泊一侧半径约500 m范围内的鸟类数据,从左到右扫描鸭类群落,以避免重复计数. 选择复兴圩、金保圩、燕窝圩、罗咀圩和老母猪墩5个样地,对各样地中越冬鸭属鸟类的群落结构、行为特征及样地的生境特征进行调查(图1).
采用群体瞬时扫描取样法观察鸭属鸟类群落中优势种的行为特征,从左到右选择不少于30只的群体进行扫描,记录每个个体的行为类型. 取样时间为8:00-17:59,每小时扫描的间隔时间为15 min,每次不少于5 min[14-16,24]. 将行为分为6类:觅食(寻找、捕捉及吞咽食物的过程)、休息(睡觉及睡姿的移动)、理羽(理羽、展翅、嬉水和挠头)、运动(游动、飞行和走动)、警戒(直立颈部密切关注周围环境的变化)和社会(追逐、嬉戏和攻击等)[6-9,12-15,26]. 将越冬时期分为前期(2017年10月底到12月底)、中期(2018年1月初到2月底)和后期(2018年3月初到4月中旬). 共收集了265 h的行为数据,扫描了1160次和58427只次.
数据收集期间实时记录气温. 结合Google Earth测量样地的面积和周长,目测记录水域和裸滩占样地面积的比例. 借助船筏等工具,根据样地面积,采用等距离设置法设置采样点,每个采样点设置2~4个样方,沉水植物样方面积为0.5 m×0.5 m,浮水和挺水植物样方面积为1 m×1 m. 记录各样方水生植物的种类和盖度,进而计算整个样地的水生植物数据,每个越冬期收集一次各样地的水生植物数据[27-28]. 11月初在各样地中放置多个测深杆以估测水深,将水深分为<10、10~50、50~100和>100 cm 4个等级. 将人为活动干扰情况分为3个等级,样点或样地边界距居民区和县乡道的垂直距离<200 m,数据收集期间(即样点计数或行为扫描期间)样点周边或样地内船只数量超过3只,样点或样地周边行人和车辆超过10次,均为强度干扰;样点或样地边界距居民区和县乡道垂直距离>500 m,数据收集期间样点或样地及周边无船只、行人和车辆,均为弱度干扰;介于二者之间为中度干扰[4,18].
1.3 数据分析
以研究样地每公顷的鸭属鸟类数量表示密度. 以Shannon-Wiener指数(H′)表示生物多样性,采用Berger-Parker优势度指数确定鸭属鸟类群落中的优势种、常见种、稀有种和罕见种[10-11,29].
(1)
Pi=Ni/N
(2)
式中,S为研究对象种数,Pi为物种i的个体数与所有物种个体数的比值.Pi也代表物种的优势度指数,N为总物种数量,Ni为物种i的个体数量,定义当Pi≥10%时,物种i为优势种;1%≤Pi<10%时,物种i为常见种;0.1%≤Pi<1%时,物种i为稀有种;Pi<0.1%时,物种i为罕见种.
以每个种类每次扫描的结果作为一个独立样本,统计各类行为的时间比例,即某一行为的个体数占扫描中总个体数的比例,进而计算各类行为在研究时间段(8:00-8:59、9:00-9:59、…、17:00-17:59)的行为时间分配,进一步计算其在各湿地类型和越冬时期的行为时间分配,以此代表行为特征.
采用IBM SPSS 22.0对数据进行统计分析. 通过Kolmogorov-Smirnov检验数据的正态性. 若符合正态性则采用独立样本t检验和单因素方差分析(One-way ANOVA)检验不同群落结构指标、各行为时间分配在各湿地类型或越冬时期间的差异. 不符合正态性的数据采用非参数检验Mann-Whitney U检验(双独立样本)和Kruskal-Wallis H检验(多独立样本)进行分析.
采用广义线性混合模型分析影响各湿地类型中鸭属鸟类群落结构和行为特征的关键环境因素,群落结构指标包括种数、数量、密度和多样性指数,行为特征指标包括优势种的觅食和警戒行为时间分配. 选择湿地面积、水域面积比例、裸滩面积比例、周长、湿地类型、水生植物盖度、浮水植物盖度、挺水植物盖度、沉水植物盖度、气温、水深、干扰度、月份和越冬时期14个环境因素为自变量. 对各自变量与因变量、各自变量间进行Pearson相关性分析,去除同时与多个自变量的相关系数大于0.6且与因变量的相关系数小于0.6的自变量[4,7,30],最后保留了湿地面积、水域面积比例、湿地类型、挺水植物盖度、沉水植物盖度、温度、水深和干扰度8个变量. 利用基于小样本的赤池信息准则(AICc)计算各模型的值以指导模型选择,用ΔAICc(候选模型与最优模型的差异)和权重wi对所有候选模型进行排序,文中列出ΔAICc小于4的候选模型,基于最优模型参数均值计算参数估计和每个解释变量的相对重要性[4,31].
文中数据以平均值±标准差表示. 设显著性水平α=0.05.
2 结果
2.1 湿地生境特征
选择5个圩口作为研究样地,代表4种湿地类型,金保圩和燕窝圩是水生植被恢复湿地;复兴圩是退耕还湿型湿地;罗咀圩是芡实塘湿地;老母猪墩是自然湿地中的芦苇湿地. 面积最大的样地是为复兴圩(272 hm2),燕窝圩面积最小(仅19 hm2). 罗咀圩、复兴圩的水域面积比例最高,均为90.00%. 平均水深最深的是罗咀圩和老母猪墩,平均水深均超过了100 cm. 2个水生植被恢复湿地的平均水深较低,均介于10~50 cm之间. 水生植物盖度最高的样地分别为金保圩(86.67%)、燕窝圩(36.43%),罗咀圩和复兴圩的水生植被盖度较低,分别为12.14%、13.57%(表1).

表1 研究样地生境特征
2.2 群落结构和动态
2017年11月-2018年3月,研究区域共调查到越冬鸭属鸟类10种(附表Ⅰ). 其中斑嘴鸭、绿翅鸭和绿头鸭的累计调查数量超过5000只,是升金湖越冬鸭属鸟类群落中的优势种. 琵嘴鸭、赤颈鸭、罗纹鸭、针尾鸭的累计调查数量介于200~1800只,属常见种. 赤膀鸭的累计调查数量为46只,属稀有种. 白眉鸭、花脸鸭的累计调查数量小于30只,属罕见种. 各样地中,芦苇湿地中分布有最高种类(10种)和数量(5085只)的越冬鸭属鸟类(附表Ⅰ). 芡实塘湿地、退耕还湿湿地和水生植被恢复湿地分别分布有6、4和3种越冬鸭属鸟类. 在各样地中,斑嘴鸭均为升金湖越冬鸭属鸟类群落中的优势种,绿翅鸭在水生植被恢复湿地、芡实塘和芦苇湿地中为优势种,绿头鸭在各样地中属常见种.
不同湿地类型间,越冬鸭属鸟类的种数(χ2=19.90,P<0.001)、数量(χ2=9.38,P=0.03)和密度(χ2=12.15,P=0.01)均有显著性差异,多样性指数在各湿地类型间差异不显著(χ2=0.70,P=0.87). 自然芦苇湿地中鸭属鸟类群落的种数、数量、密度和多样性指数最高,芡实塘湿地和退耕还湿湿地次之,除密度外,水生植被恢复湿地中其他群落结构指数均最低(表2). 在整个越冬时期,芦苇湿地中鸭属鸟类的种数(7.00±2.55)、数量(1250.00±127.40)、密度((17.93±4.46)只/hm2)、多样性指数(0.96±0.29)均最高. 越冬前期,水生植被恢复湿地中鸭属鸟类的各群落结构指标最低;越冬中期和后期,芡实塘湿地中鸭属鸟类群落结构指标最低(图2).
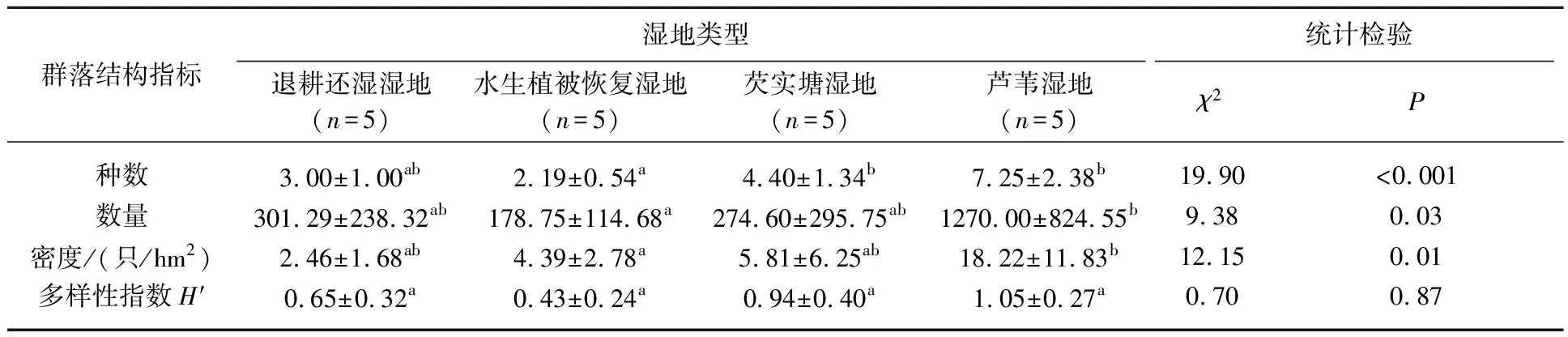
表2 不同湿地类型中越冬鸭属鸟类群落结构特征*
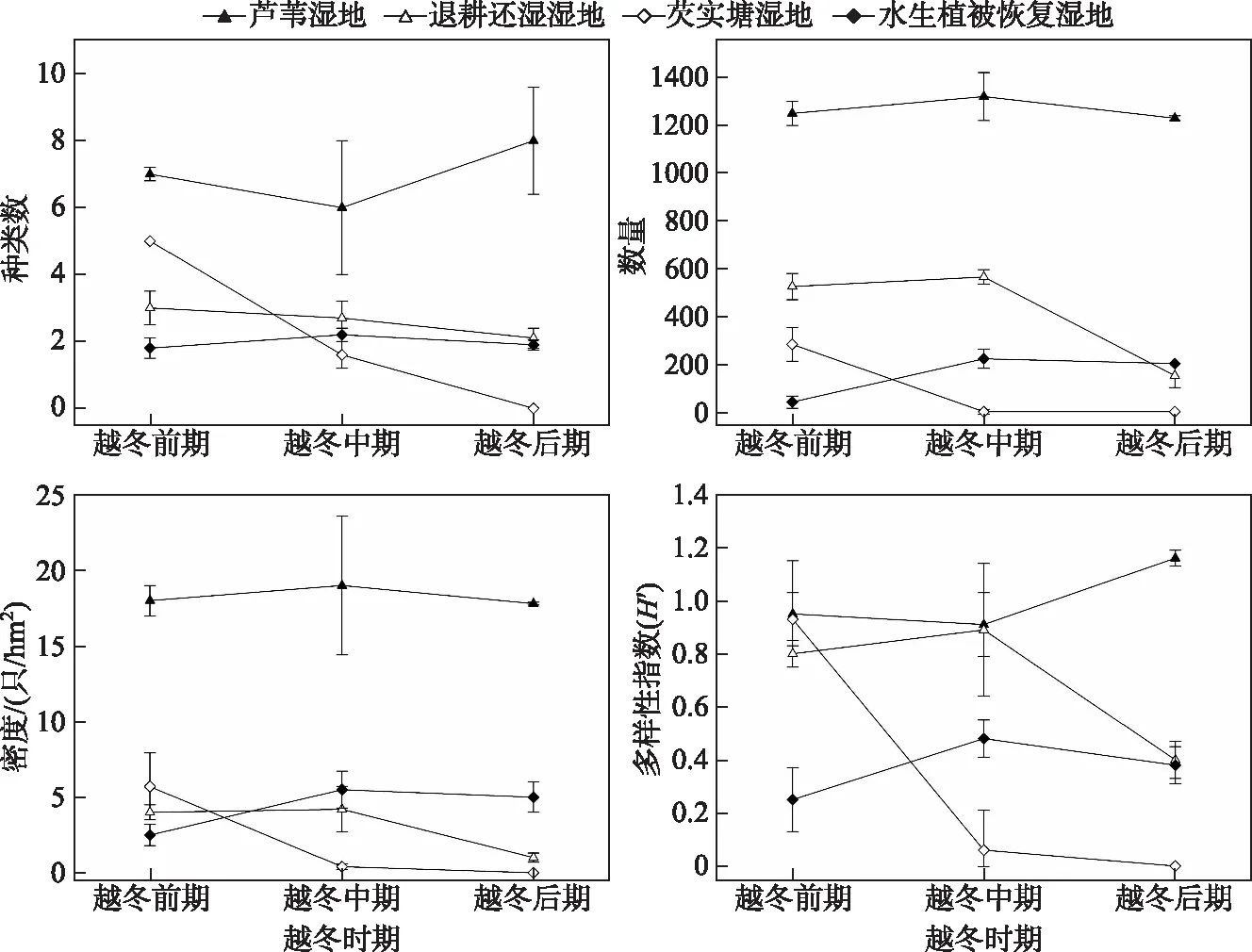
图2 鸭属鸟类群落结构在不同越冬时期的变化Fig.2 Changes of community structure of dabbling ducks in different wintering periods
2.3 优势种行为特征
对升金湖越冬鸭属鸟类群落中两种优势种-斑嘴鸭和绿翅鸭的行为特征进行研究,优势种绿头鸭在多个样地中的数量不足30只,因此未研究其行为特征.
2.3.1 斑嘴鸭的行为特征 不同湿地类型中,越冬斑嘴鸭的觅食(χ2=30.03,P<0.001)、休息(χ2=12.84,P=0.01)和警戒(χ2=32.98,P<0.001)行为时间分配具有显著性差异,运动(χ2=2.33,P=0.51)、理羽(χ2=4.89,P=0.18)和社会(χ2=1.29,P=0.73)行为时间分配差异不显著(表3). 越冬斑嘴鸭的觅食行为时间分配在芡实塘湿地中最高(28.64%±19.81%),在退耕还湿湿地中次之(20.00%±7.40%),在芦苇湿地中最低(13.37%±12.71%). 休息行为时间分配在芦苇湿地中最高(59.29%±21.48%),芡实塘中最低(42.45%±26.01%). 警戒行为时间分配在退耕还湿湿地中最高(5.02%±5.65%),且显著高于其它类型的湿地.

表3 不同湿地类型中越冬斑嘴鸭行为时间分配*
2.3.2 绿翅鸭的行为特征 不同湿地类型中,越冬绿翅鸭的觅食(χ2=42.23,P<0.001)、休息(χ2=21.42,P<0.001)、运动(χ2=58.75,P<0.001)、理羽(χ2=23.94,P<0.001)、警戒(χ2=10.38,P=0.02)和社会(χ2=8.84,P=0.03)行为时间分配均有显著性差异(表4). 觅食行为时间分配在芡实塘湿地中最高(18.33%±12.88%),芦苇湿地中最低(4.29%±4.06%). 休息行为时间分配在水生植被恢复湿地中最高(55.24%±26.68%),芦苇湿地中最低(34.73%±20.16%). 运动行为时间分配在芦苇湿地中最高(52.01%±17.74%),且显著高于其它湿地类型. 绿翅鸭的警戒和社会行为时间比例均小于1.00%,其中警戒行为时间分配在水生植被恢复湿地中最高(0.50%±1.58%),芦苇湿地中最低(0.36%±0.84%).
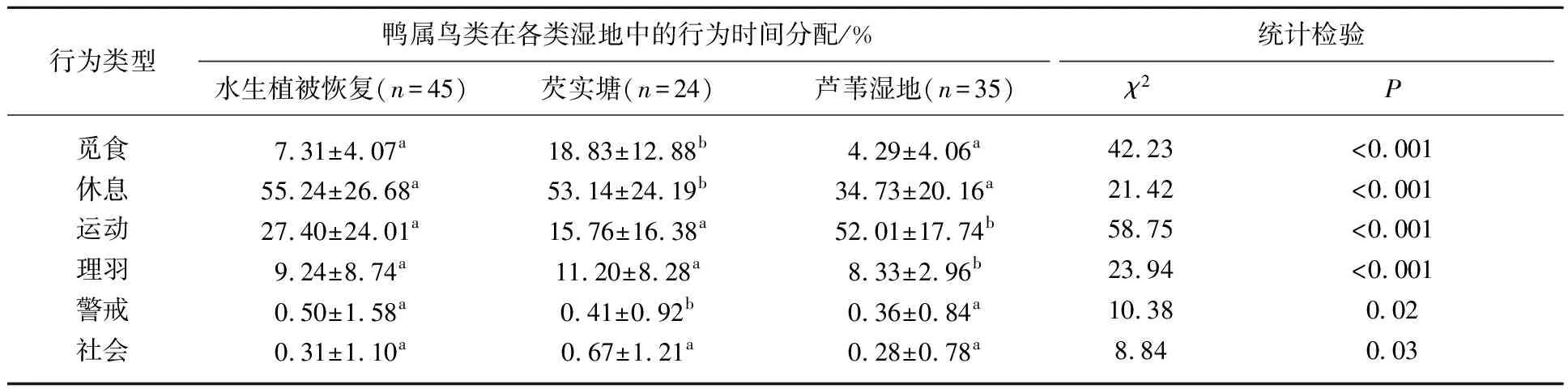
表4 不同湿地类型中越冬绿翅鸭行为时间分配*
2.4 影响鸭属鸟类群落结构的环境因素
利用广义线性混合模型分析影响各湿地类型中越冬鸭属鸟类种数、数量、密度和多样性指数的关键环境因素. 影响种数的最优模型包括湿地面积、干扰度、水域面积和湿地类型4个变量,种数与湿地的面积、水域面积比例和湿地类型呈正相关,与干扰度则呈负相关;影响数量的最优模型包括干扰度1个变量,呈负相关;影响密度的最优模型包括挺水植物盖度1个变量,呈正相关;影响多样性指数的最优模型包括干扰度和水域面积比例2个变量,与干扰度呈负相关,与水域面积比例呈正相关(表5、6).

表5 广义线性混合模型评估越冬鸭属鸟类群落结构*
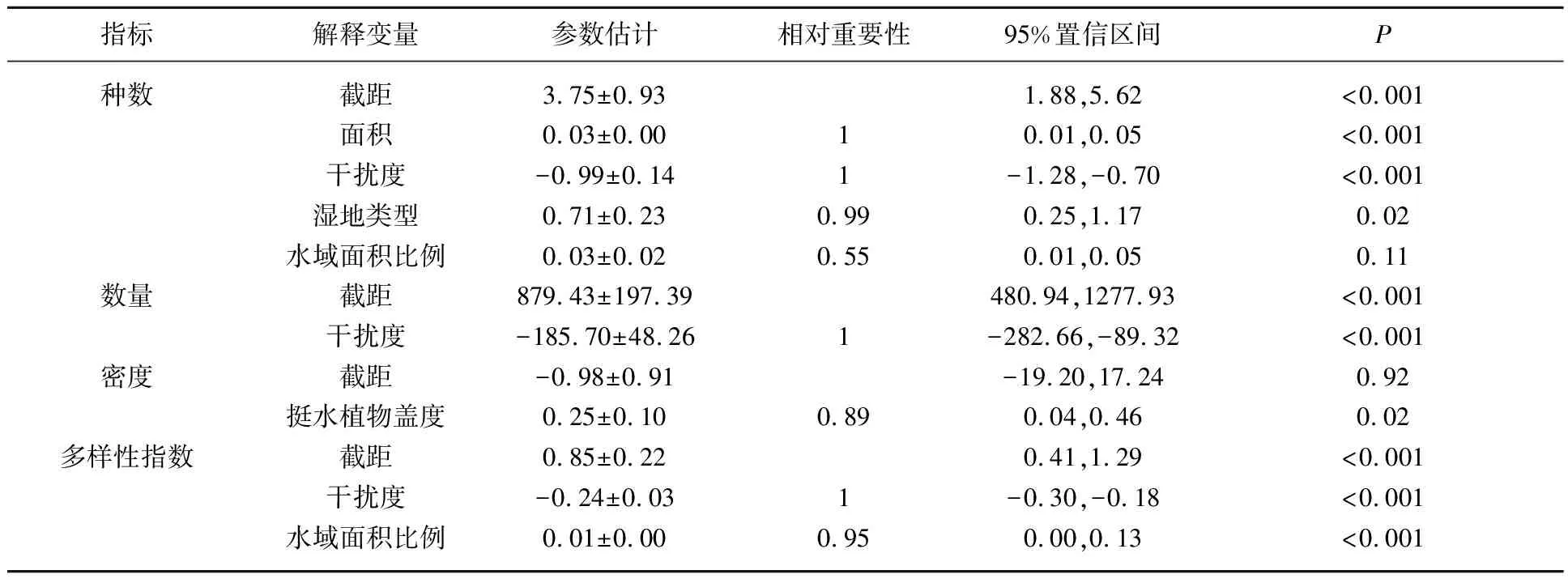
表6 评估越冬鸭属鸟类群落结构最优模型的平均参数估计和相对重要性*
2.5 影响优势种行为特征的环境因素
利用广义线性混合模型分析影响各湿地类型中斑嘴鸭和绿翅鸭觅食行为、警戒行为特征的关键环境因素. 影响斑嘴鸭觅食行为时间分配的最优模型包括气温、水深和干扰度3个变量,与气温呈正相关,与水深和干扰度呈负相关;影响斑嘴鸭警戒行为时间分配的最优模型包括干扰度和湿地类型2个变量,与干扰度呈正相关. 影响绿翅鸭觅食行为时间分配的最优模型包括气温、水深和沉水植物盖度3个变量,其中与气温和沉水植物盖度呈正相关系,与水深呈负相关;影响绿翅鸭警戒行为时间分配的最优模型包括干扰度和挺水植物盖度2个变量,与干扰度呈正相关,与挺水植物盖度呈负相关(表7、8).
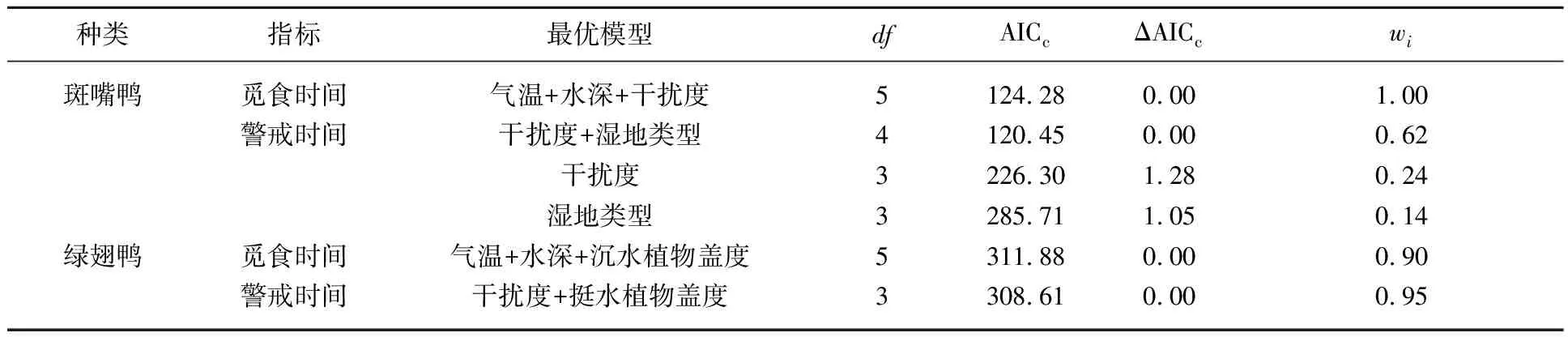
表7 广义线性混合模型评估越冬鸭属鸟类行为特征*

表8 评估越冬鸭属鸟类行为特征最优模型的平均参数估计和相对重要性*
3 讨论
3.1 越冬鸭属鸟类的群落结构
长江中下游湖泊是东亚-澳大利西亚候鸟迁徙路线上主要的候鸟越冬地和迁徙停歇地,升金湖是长江中下游湖泊湿地的重要组成部分[11,23],鸭科水鸟占升金湖所有越冬水鸟数量的77%,是升金湖越冬水鸟中的优势群落,鸭属在鸭科水鸟中种类最多[23]. 中国分布有12种鸭属鸟类[32],升金湖分布达10种. 鸭属鸟类均为食性泛化种,以水生无脊椎动物、水生植物种子、水生植物和陆生草本植物的嫩枝芽为食[5,16],在升金湖湖区沉水植物资源严重退化的情况下,临近湖区的各类养殖塘和水生植被恢复的湿地已经成为越冬水鸟特别是鸭属鸟类在升金湖的替代性栖息地.
不同湿地类型的景观结构和生境特征不同,在越冬水鸟的栖息地选择中发挥了不同的作用. 鸭类偏好选择隐蔽性强、水域开阔、微生境复杂的湿地栖息,如芦苇湿地[15,21-22,33-35]. 芦苇湿地作为自然湿地,仍是水鸟的重要栖息地. 5个样地中,鸭属鸟类的种数、数量、密度和多样性指数在芦苇湿地最高,老母猪墩距离公路和居民区较远(>1 km),人为活动弱且芦苇(Phragmitescommunis)和菰(Zizanialatifolia)等挺水植物盖度高,是鸭类适宜的隐蔽栖息地. 水生植被恢复湿地种植有盖度较高的水生植被,但鸭属鸟类群落的种数、数量、密度和多样性指数较低,这可能是由于金保圩、燕窝圩2个样地距离居民区较近(<100 m),人类活动频繁且面积较小. 芡实塘和退耕还湿湿地也吸引了种类和数量较多的鸭类在此觅食,这可能是因为两种湿地种植了芡实(Euryaleferox)、苦草(Vallisnerianatans)等水生植物,而苦草的块茎和农户收割后遗留的芡实,能为鸭属鸟类提供丰富的食物来源[19,36]. 在升金湖分布的10种越冬鸭属鸟类中,斑嘴鸭在本研究选择的5个样地中均为优势种,绿翅鸭则是芦苇和水生植被恢复湿地中的优势种. 与许多研究的结果相似,越冬斑嘴鸭对各类生境均有较强的适应性,绿翅鸭则偏好选择隐蔽性强的湿地栖息[6,8,12-13,15].
越冬前期,芡实塘和退耕还湿湿地还未开展芡实收割作业,多个样地的水深介于10~50 cm之间,还处于适宜鸭类栖息的水平,同时越冬前期是迁徙候鸟到达越冬地的高峰期[23]. 越冬中期,芡实塘中的水被全部排干,复兴圩和罗咀圩的芡实收割作业陆续开展,多个湿地均已不适宜鸭类栖息. 到了越冬后期,随着繁殖期的到来,迁徙水鸟陆续回迁繁殖地. 因此可以看出,在本研究所选择的样地中,升金湖越冬鸭类群落的种数、数量、密度、多样性指数等群落结构指标呈越冬前期-中期-后期不断降低的趋势.
3.2 越冬鸭属鸟类在不同湿地中的行为特征
鸟类花费在各种行为上的时间和能量都会影响到其生存状态,鸟类的行为时间分配是对环境变化的一种适应. 本研究中越冬鸭属鸟类的觅食时间在芡实塘湿地中最高(斑嘴鸭为28.64%±19.81%,绿翅鸭为18.83%±12.88%),与崇明东滩水产养殖塘中越冬绿头鸭的觅食时间(21.97%)及法国西海岸自然保护区越冬绿头鸭觅食时间(5%~20%)相似[7-8,26]. 芡实塘中丰富的食物资源使鸭类能够花费较少的时间觅食就能获得充足的食物,其余更多的时间则用于休息和游动,以这种行为策略来节省能量支出,达到自身最大的适合度. 斑嘴鸭的觅食时间在水生植被恢复湿地中最低(13.02%±10.30%),绿翅鸭觅食时间在芦苇湿地中最低(4.29%±4.06%),与田纳西州自然保护区水生植物栽种实验区的绿头鸭、绿翅鸭和琵嘴鸭的觅食时间相似(<20%)[15],这可能是因为挺水植物恢复湿地燕窝圩和芦苇湿地的挺水植物盖度高,隐蔽条件好,鸭类主要行为是休息和游动以躲避风险,而金保圩作为沉水植被恢复湿地,苦草块茎等水鸟的食物资源可能在秋季时就已消耗殆尽. 斑嘴鸭警戒时间在退耕还湿湿地中最高(5.02%±5.65%),且显著高于其它类型湿地,该结果与乌鲁木齐白湖湿地中的白眼潜鸭(A.nyroca)(5.10%)、鄱阳湖藕塘中越冬小天鹅(C.cygnus)(<5%)的警戒时间相似[37-38]. 本研究中,退耕还湿样地中虽然食物资源丰富,但频繁的行人、车辆和芡实收割作业使得鸭类不得不在觅食的同时增强警戒行为的强度.
3.3 越冬鸭属鸟类栖息地选择的影响因素
水鸟对湿地的栖息地利用和行为适应受湿地生境质量的直接影响[2-7,17-22,33-35]. 广义线性混合模型评估结果表明湿地面积是影响越冬鸭类群落在不同湿地中种数的关键因素. 面积大小直接影响水鸟对湿地的利用,面积越大的湿地往往水域开阔、微生境复杂,能容纳更多种类和数量的水鸟[11, 19]. 结果还发现鸭属鸟类的种数和多样性指数与水域面积比例呈正相关. 水域面积对水鸟特别是游禽的栖息地利用有很大影响,水域面积越大,游禽可利用的栖息地面积越大[19-22,24]. 影响鸭属鸟类群落的种数、数量和多样性指数的最优模型均包括干扰度且为负相关,显然,湿地周边频繁的人为活动对水鸟造成了不利影响. 湿地中的挺水植物能为水鸟提供隐蔽条件[6,22,24],本研究结果表明挺水植物盖度越高,鸭属鸟类群落的密度越高.
影响斑嘴鸭和绿翅鸭在湿地中觅食行为的关键环境因素是水深、气温、干扰度和沉水植物的盖度,影响警戒行为的关键环境因素是干扰度和挺水植物的盖度. 随着水深的增加,鸭类的觅食时间趋于减少. 有大量研究已经表明水深是影响觅食行为的主要因子,水深直接决定了水鸟对湿地的可利用性[4-7,19-22,33-35]. 本研究发现气温是影响鸭属鸟类觅食时间的关键因素之一. 不同越冬时期气温不同,越冬前期,水鸟刚刚抵达越冬地,需要花费较多的时间觅食以补充能量;越冬后期,水鸟会增加觅食以应对即将到来的北迁;而在越冬中期,升金湖气温常低于0℃,水鸟多采取静息的方式维持体能[12,14,26]. 在人口密度较高的长江中下游地区,人类活动带来的干扰是各类湿地最主要的特征之一,人为干扰对水鸟的行为有显著影响[4,18]. 本研究的样地周边农渔业活动和行人、车辆频繁,研究结果表明随着干扰度的增强,斑嘴鸭和绿翅鸭的觅食时间呈降低趋势,警戒行为则增强. 研究发现绿翅鸭的觅食时间随湿地中沉水植物盖度的增高而增加,这可能是因为在冬季,几个恢复类型的湿地中仍有苦草块茎等沉水植物成为鸭类的食物来源. 挺水植物能为水鸟提供隐蔽条件[6,22,24],本研究即发现绿翅鸭在挺水植物盖度高的湿地降低了警戒行为的强度.
4 结论和建议
本文以长江中下游重要湿地升金湖为研究区域,对鸭属鸟类这一主要的水鸟类群开展了越冬生态学研究,从而为该区域的越冬水鸟保护提供理论基础. 研究发现除自然湿地外,多种恢复类型的湿地成为了越冬鸭属鸟类的重要栖息地,鸭属鸟类偏好选择面积大、水深介于10~50 cm、水生植物盖度高和人类活动少的湿地栖息,种植沉水植物的芡实塘和退耕还湿湿地成为鸭属鸟类重要的觅食地,挺水植物丰富的芦苇和水生植被恢复湿地成为鸭属鸟类躲避风险的主要场所. 结果提示,在保护自然湿地的同时,恢复多种类型的湿地栖息地对于越冬水鸟的保护具有重要意义.
建议在自然湖泊的浅水区和湖滨带辅以人工手段种植沉水和挺水等水生植物,快速恢复水生植被,为冬候鸟提供丰富的食物来源和适宜的隐蔽条件;芡实收割作业时可适当保留,为冬候鸟提供一定的食物来源;合理调控湿地水深,湿地的水文管理方面应尽量避免短时间内的排干,水深保持在10~50 cm左右能容纳更多种类和数量的鸭属鸟类;冬季,通过管理减少各湿地周边的人为活动可提高水鸟群落的多样性.
5 附录
附表Ⅰ见电子版(DOI:10.18307/2022.0514).
