瑶山亚种树鼩IFN-β和IFN-γ原核表达及其多克隆抗体制备
2022-09-05曹颖颖李宝莹运晨霞唐海波
曹颖颖,李宝莹,王 婷,梁 亮,运晨霞,唐海波,冷 静
(广西中医药大学/广西高发传染病中西医结合转化医学重点实验室/广西特色实验动物病证模型重点实验室,广西南宁 530200)
0 引言
【研究意义】树鼩()是在进化关系上除灵长类动物外与人类最接近的物种(Novacek,1992;Waddell et al.,2001),在生理机能、生化代谢和基因组等方面均与人类相似,且已证实树鼩能被多种人类疾病相关的病毒感染,在一些特殊人类疾病动物模型上具有独特优势(Glebe et al.,2003;Yan et al.,2012)。但作为一种新兴的实验动物,树鼩的基础生物学数据及专有的商品化检测试剂匮乏,极大限制了其推广应用。因此,制作树鼩源性抗体等检测试剂对促进树鼩实验动物模型的应用及研究具有重要意义。【前人研究进展】自Lindblad-Toch等(2011)利用美国Board研究所公布的不完整树鼩基因组数据重构29个哺乳动物系统发育关系发现树鼩是最接近灵长类的物种以来,滇西亚种树鼩在基因组学、转录组学、蛋白组学及解剖学等方面已得到一定研究(Li et al.,2012;Fan et al.,2013),且证实在构建肿瘤模型(Park et al.,2000)、乙肝病毒感染模型(Park et al.,2000)、抑郁症模型(Fuchs,2005;王静等,2011)等方面具有广阔的应用前景。在树鼩专有生物制剂研究方面,干细胞生长因子、白细胞介素6、载脂蛋白AV等细胞因子及蛋白已有表达纯化的研究报道(王艳等,2013;龙琼等,2019)。干扰素(Interferon,IFN)是研究最早的一类细胞因子,具有种属特异性(Samarajiwa et al.,2009;吴丹等,2013),其家族成员中的IFN-α、IFN-β和IFN-γ已广泛应用于人类病毒性疾病与癌症治疗。IFN属于多效性细胞因子,与相应受体结合并激发细胞信号级联反应,通过激活多种干扰素刺激基因(Interferon-stimulated gene,ISG)的转录与表达,而发挥抗病毒、抗肿瘤及免疫调节等作用(Osterlund et al,2005;罗婵等,2015)。针对树鼩IFN的研究主要有:李明利等(2012)以树鼩全基因组数据为基础,通过基因预测和蛋白建模等方法对树鼩IFN家族的基本构成进行了系统分析;丁晨等(2018)通过构建原核重组表达载体克隆表达出滇西亚种树鼩IFN-γ,且证实复性纯化后具有良好的免疫原性。【本研究切入点】滇西亚种和瑶山亚种树鼩是主要分布在我国云南和广西的2个树鼩亚种,相对于滇西亚种树鼩,瑶山亚种树鼩的基础生物学研究匮乏,其免疫系统研究尚缺少商品化的树鼩特异性抗体等生物制剂,且不同亚种树鼩间的遗传差异和分化程度尚无详细数据。【拟解决的关键问题】通过RT-PCR扩增瑶山亚种树鼩的和基因,构建原核表达载体进行诱导表达,经免疫小鼠获得多克隆抗体并鉴定其反应性,以期为进一步研究树鼩病毒感染动物模型打下基础。
1 材料与方法
1.1 试验材料
瑶山亚种树鼩购自广西中医药大学人工驯化养殖实验中心,麻醉后静脉采血用于外周血淋巴细胞分离;昆明小鼠[SCXK(湘)2019-0004]购自湖南斯莱克景达实验动物有限公司。动物试验经广西中医药大学实验动物福利伦理审查委员会审查通过(DW20201206-123),严格遵循3R原则。原核载体pET28a(+)由广西中医药大学刘鹏副教授惠赠,pcDNA3.1(+)由广西高发传染病中西医结合转化医学重点实验室保存提供;大肠杆菌BL21(DE3)和DH5α感受态细胞购自北京全式金生物技术有限公司;淋巴细胞分离液购自沃卡威(北京)生物技术有限公司;脂质体2000及限制性内切酶H I和I购自赛默飞世尔科技公司;分子克隆相关试剂盒购自天根生化科技(北京)有限公司;弗氏完全佐剂和弗氏不完全佐剂购自上海碧云天生物技术有限公司;碱性磷酸酶标记马抗小鼠IgG抗体购自北京中衫金桥生物技术有限公司;FITC标记山羊抗小鼠IgG抗体购自美国Sigma公司。
1.2 试验方法
1.2.1 瑶山树鼩和基因克隆及真核/原核表达质粒构建 根据GenBank已公布的滇西亚种树鼩和基因序列设计引物,委托生工生物工程(上海)股份有限公司合成。采用TRIzol法提取树鼩外周血淋巴细胞总RNA,并反转录合成cDNA,以引物IFN-β-1(5'-ATGTTCTACAGAAGC ATCCT-3')和IFN-β-2(5'-TCAGTCTCGGAGGTAG TCTA-3')、IFN-γ-1(5'-ACCAGGGCTACCGATTTG A-3')和IFN-γ-2(5'-TTATCTGGAAGCTCTCCGAC C-3')扩增包含ORF的瑶山亚种树鼩和基因片段,然后克隆至pMD18-T载体上,挑取单克隆进行菌液PCR鉴定,经测序验证后再以亚克隆引物pcDNA3.1-IFN-β-U(5'-CGGATCCGCCACCATGT TCTACAGAAGCATCC-3',划线部分为H I酶切位点)和pcDNA3.1-IFN-β-L(5'-CCTCGAGTCAGT CTCGGAGGTAGTCTAT-3',划线部分为I酶切位点)、pcDNA3.1-IFN-γ-U(5'-CGGATCCGCCACC ATGAAGTATACAAGTTATA-3',划线部分为H I酶切位点)和pcDNA3.1-IFN-γ-L(5'-CCTCGAGTT ATCTGGAAGCTCTCCGAC-3',划线部分为I酶切位点)进行PCR扩增并回收目的条带,采用限制性内切酶H I和I在37 ℃下酶切目的条带和质粒pcDNA3.1(+),酶切产物经胶回收试剂盒回收后用T4 DNA连接酶连接,构建真核表达载体pcDNA3.1-(图1-A)和pcDNA3.1-(图1-B);经酶切及测序鉴定正确后,将得到的阳性重组质粒转染仓鼠肾细胞(BHK-21)进行真核表达。原核表达载体(图1-C和图1-D)的构建同真核表达载体。在基因测序完成后利用DNAStar对IFN-β和IFN-γ氨基酸序列进行比对,在不影响蛋白主要免疫活性及免疫原性的前提下剔除掉疏水氨基酸序列,最终设计引物如下,pET28a-IFN-β1(5'-CGGATCC ATGGACTGCCTCAAGGACCGTAT-3',划线部分为H I酶切位点)和pET28a-IFN-β2(5'-CCTCGAG ACAGCTGCTGTACTCCTTGG-3',划线部分为I酶切位点)、pET28a-IFN-γ1(5'-CGGATCCATGCCA TTTATGAAAGAAGTACA-3',划线部分为H I酶切位点)和pET28a-IFN-γ2(5'-CCTCGAGTCTGGA AGCTCTCCGACCTC-3',划线部分为I酶切位点),构建的原核表达载体pET28a-和pET28a-转化BL21(DE3)感受态细胞进行原核表达(唐海波等,2014;赵祥秀等,2020)。

1.2.2 重组蛋白表达纯化及鉴定 挑取转化了原核表达载体pET28a-和pET28a-的菌株单克隆,接种至含100 μg/mL Kan的LB液体培养基中,37 ℃下摇床(220 r/min)培养至OD约0.6,取出1 mL留样保存,其余加入终浓度为0.5 mmol/L的IPTG,30 ℃下诱导表达6 h,收集菌体,取出1 mL留样保存,剩余菌体进行超声波破碎,破碎后4 ℃下12000 r/min离心15 min分离上清液和沉淀,同样取适量上清液和沉淀留样保存。将上述每步留取的样品进行SDS-PAGE电泳,鉴定融合蛋白的表达效果及表达形式。鉴定后的融合蛋白采用含1、2、3和4 mol/L尿素的洗涤液进行洗涤,SDS-PAGE电泳摸索最佳包涵体洗涤条件,洗涤获得的包涵体以梯度递降尿素浓度透析的方法复性,-80 ℃保存备用(孟先明等,2010)。Western blotting检测鉴定融合蛋白:纯化复性后的融合蛋白经SDS-PAGE电泳后转移至PVDF膜上,以鼠抗His标签抗体为一抗、碱性磷酸酶标记马抗小鼠IgG抗体为二抗,BCIP/NBT碱性磷酸酯酶显色试剂盒显色。
1.2.3 抗体制备及反应性检测 将弗氏佐剂与纯化复性后的融合蛋白(IFN-β和IFN-γ)乳化后免疫昆明小鼠(100 μg/只),免疫周期为第1、7、28、35和45 d,免疫方式为背部皮下多点注射。首次免疫用弗氏完全佐剂与融合蛋白乳化,第2、3、4和5次免疫则用弗氏不完全佐剂与融合蛋白乳化。第5次免疫后1周经眼球采血并分离血清,通过Western blotting和IFA检测制备获得多克隆抗体的反应性。Western blotting检测以不同稀释度的抗IFN-β和IFN-γ多克隆抗体为一抗,未免疫小鼠血清为阴性对照,碱性磷酸酶标记马抗小鼠IgG抗体为二抗,BCIP/NBT显色观察。间接免疫荧光(IFA)检测采用转染24 h后的单层BHK-21细胞,按1∶100分别稀释制备的抗IFN-β、IFN-γ多克隆抗体及未免疫小鼠阴性血清为一抗,以1∶300稀释FITC标记的山羊抗小鼠IgG为二抗,在荧光显微镜下进行观察。
2 结果与分析
2.1 瑶山亚种树鼩IFN-β和IFN-γ基因克隆结果
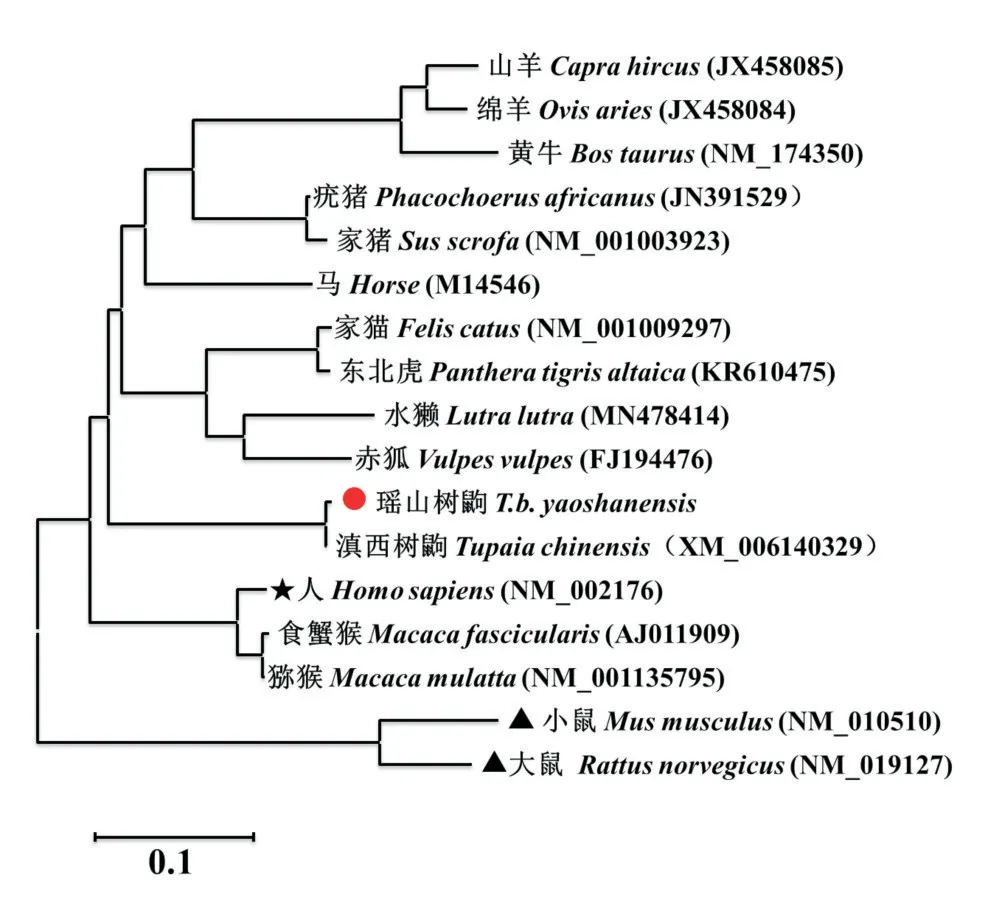
以瑶山亚种树鼩外周血淋巴细胞为材料提取总RNA,Oligo(dT)引物反转录合成的cDNA为模板,分别扩增获得564 bp的基因片段(图2-A)及501 bp的基因片段(图2-B),与预期的目的基因片段长度相符。将和基因克隆至pMD18-T载体上并转化DH5α感受态细胞,取少量转化菌株涂布于LA培养基上,37 ℃过夜培养后挑取单菌落进行菌液PCR鉴定,琼脂糖凝胶电泳结果显示获得对应的目的条带(图2-C和图2-D),表明和基因已成功克隆至pMD18-T载体上。对阳性克隆质粒进行测序分析,发现与GenBank已公布滇西亚种树鼩基因(XM-006140329)、基因(KC905628)的核苷酸序列相似性分别为99.6%和100.0%。基于和基因的核苷酸序列相似性构建系统发育进化树(图3和图4),结果发现与啮齿类大鼠、小鼠的IFN相比,树鼩的IFN-β和IFN-γ更接近人类IFN。
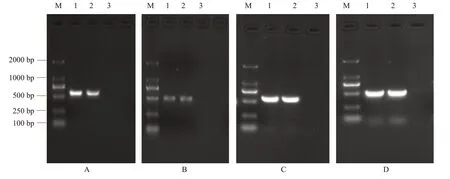

2.2 瑶山亚种树鼩IFN-β和IFN-γ真核/原核表达载体
以添加有酶切位点的亚克隆引物扩增重组质粒pMD18-T-和pMD18-T-,回收PCR扩增产物再次与pMD18-T载体连接,然后转化DH5α感受态细胞,挑取单个菌落进行菌液PCR鉴定,阳性克隆测序分析正确后分别酶切pcDNA3.1(+)和亚克隆阳性质粒,将酶切产物跑琼脂糖凝胶电泳,胶回收带有酶切位点的、基因片段及酶切后的线性pcDNA3.1(+)空载体,用T4 DNA连接酶将目的基因片段连接至真核表达载体pcDNA3.1(+)上,然后转化DH5α感受态细胞,挑取单克隆进行菌液PCR鉴定和酶切鉴定,结果(图5)显示均为阳性质粒。经测序分析,瑶山亚种树鼩和基因在真核表达载体中读码正确,说明真核表达载体pcDNA3.1-和pcDNA3.1-构建成功。原核表达载体构建方式同真核表达载体,分别命名为pET-28a-和pET-28a-。

2.3 融合蛋白IFN-β和IFN-γ表达纯化及鉴定结果
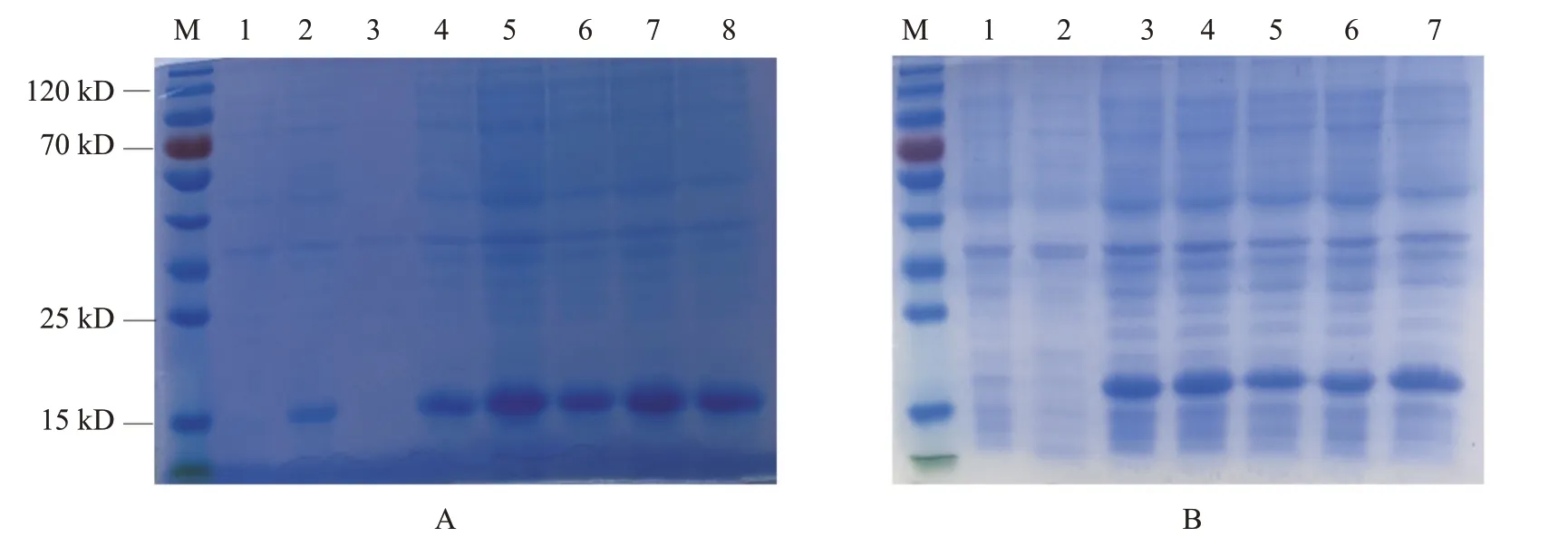
以原核表达载体pET-28a-和pET-28a-转化BL21(DE3)感受态细胞并诱导表达,结果发现以30 ℃下0.5 mmol/L IPTG诱导6 h获得的融合蛋白表达量最高(图6-A和图6-B)。为验证融合蛋白的表达方式,经超声波破碎后进行SDS-PAGE电泳,结果在细胞沉淀中检测到融合蛋白IFN-β和IFN-γ,说明融合蛋白主要以包涵体形式表达(图6-C和图6-D)。以含不同浓度尿素的洗涤液洗涤包涵体进行初步纯化,结果(图7)显示,包涵体经含1、3和4 mol/L尿素的洗涤液洗涤后,仍有较多杂蛋白存在于目的蛋白中;而经含2 mol/L尿素的洗涤液洗涤后可获得纯度相对较高的融合蛋白,故确定2 mol/L尿素为最佳包涵体洗涤浓度。由于诱导表达融合蛋白的pET-28a(+)系统上带有多聚组氨酸(6×His)标签,因此可利用携带His标签的抗体进行Western blotting检测,结果表明,纯化的融合蛋白与预测的目的蛋白基本一致(图8),说明瑶山亚种树鼩和基因在原核表达载体中正确表达。
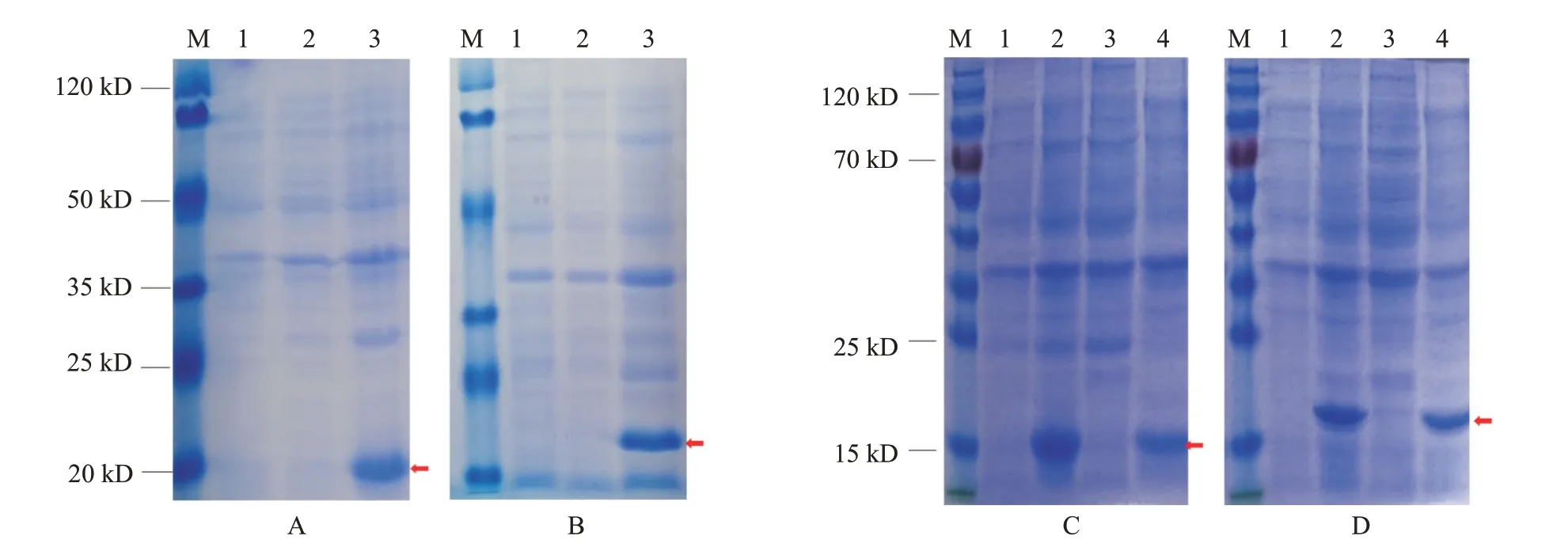
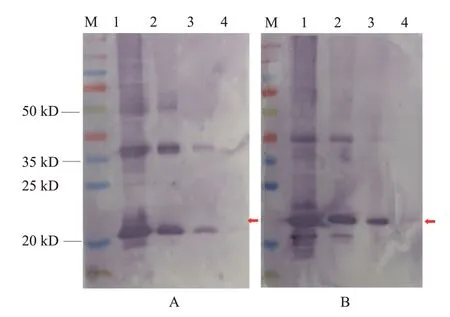
2.4 多克隆抗体Western blotting检测结果
以复性后的融合蛋白IFN-β和IFN-γ与佐剂乳化后分别免疫昆明小鼠,第5次免疫后1周采集血液分离血清,所得血清即为多克隆抗体。Western blotting检测结果(图9)显示,制备获得的IFN-β多克隆抗体在稀释度为1∶8000时仍能与0.1 μg的融合蛋白IFN-β结合,IFN-γ多克隆抗体在稀释度为1∶8000时仍能与1.0 μg的融合蛋白IFN-γ结合,均具有良好的反应性。

2.5 多克隆抗体IFA检测结果
通过IFA进一步验证制备获得的多克隆抗体与IFN-β和IFN-γ的结合能力,结果(图10)显示,经真核表达载体pcDNA3.1-和pcDNA3.1-转染的BHK-21细胞分别与制备获得的IFN-β和IFN-γ多克隆抗体共孵育后,均能观察到特异的绿色荧光,而与阴性血清共孵育的转染BHK-21细胞未观察到绿色荧光。

3 讨论
IFN作为机体抵御病毒感染的第一道防线,具有抗病毒、抗肿瘤、免疫调节及抑制细胞增殖等多重作用(Kotenko et al.,2003;Brand et al.,2005;Osterlund et al.,2005;Ank et al,2006;李聪聪等,2021)。树鼩是一种外形与松鼠类似而遗传上最接近灵长类的动物,在病毒感染模型方面相对于啮齿类实验动物有着不可替代的作用。本研究通过RT-PCR扩增获得瑶山亚种树鼩的和基因片段(包含整个ORF),遗传进化分析发现瑶山亚种树鼩的(I型)和(II型)基因与人类、食蟹猴的基因处于更接近的分支上,进一步验证其进化上的优势。此外,树鼩能被多种与人类疾病有关的病毒感染(Xu et al.,2007;Li et al.,2008)。与灵长类实验动物相比,树鼩具有形体小、繁殖快及成本低等特点,在医学研究中的价值已受到越来越多的关注。迄今为止,科学家们对树鼩的解剖学、生殖生理学、中枢神经系统、视觉系统及动物模型等已进行大量研究(Eichmann and Holst,1999;Fuchs and Flügge,2002;Pradidarcheep et al.,2003;Norton et al.,2010)。在病毒学研究方面,树鼩已成为研究肝炎病毒、轮状病毒等病毒感染及致病机制的理想动物模型(Collins et al.,2007;Norton and Siegwart,2013)。近年来,针对滇西亚种树鼩的基础生物学研究较深入,但树鼩作为病毒感染动物模型其免疫系统、免疫应答相关细胞因子的研究相对匮乏,且市面上缺乏树鼩特异性抗体等生物制剂,致使流式细胞术、免疫组织化学等研究难以开展,严重限制了树鼩作为病毒感染动物模型在相关研究领域的应用。
本研究通过构建原核表达载体成功诱导表达获得瑶山亚种树鼩IFN-β和IFN-γ蛋白,且证实获得的融合蛋白主要以包涵体形式表达。原核表达载体pET-28a(+)携带有His标签,为后期的蛋白纯化及检测提供了方便。瑶山亚种树鼩和基因长度分别为564和501 bp,通过预测瑶山树鼩IFN-β和IFN-γ蛋白结构发现:IFN-γ蛋白N端富含疏水性氨基酸,IFN-β蛋白N端和C端均富含疏水性氨基酸,可能与IFN的分泌功能有关。在不影响蛋白主要免疫活性及免疫原性的前提下,本研究在设计引物时截去N端疏水性氨基酸,通过原核表达载体成功诱导表达获得融合蛋白IFN-γ,但未表达出融合蛋白IFN-β。为此,在重新设计引物时同时去除N端和C端疏水性氨基酸,最终成功通过原核表达载体诱导表达获得融合蛋白IFN-β,且免疫原性及活性完好。由于富含疏水性氨基酸肽段在原核载体中不易表达,因此试验设计时在保留免疫活性及免疫原性的基础上应尽可能去除富含疏水性氨基酸肽段的表达。
在融合蛋白纯化方面,本研究尝试在变性条件下采用Ni-NTA进行纯化,但发现融合蛋白IFN-β和IFN-γ均未能被吸附在树脂上,大部分融合蛋白随穿透液流出,可能是融合蛋白以包涵体形式表达时其构象将6×His标签包裹在内部,而His标签在8 mol/L尿素变性条件下仍暴露不够充分(赵慧等,2009)。孟先明等(2010)曾报道,在Ni-NTA纯化不理想时可尝试通过采用含有不同浓度尿素的洗涤液充分洗涤包涵体以获得较高纯度的目的蛋白。为此,本研究将0.5 mmol/L IPTG诱导6 h的重组菌株在液氮反复冻融3次后以超声波裂解菌体,由于大部分菌体自身蛋白为可溶性蛋白,通过离心可去除绝大部分可溶性蛋白,实现包涵体与可溶性蛋白分离,收集获得的包涵体中杂蛋白含量较少。包涵体洗涤液中含有TritonX-100,可将吸附在包涵体表面的一些不溶杂蛋白除去,再以含不同尿素浓度的洗涤液进行洗涤,结果显示,经含l、3和4 mol/L尿素的洗涤液洗涤后SDS-PAGE电泳的杂带还是相对较多,而以含2 mol/L尿素的洗涤液洗涤后可获得纯度相对较高的融合蛋白,且损失较少,故确定包涵体洗涤液中的尿素最佳浓度为2 mol/L。融合蛋白IFN-β和IFN-γ经乳化后免疫小鼠,成功制备获得多克隆抗体,Western blotting和IFA检测结果均显示,制备获得的2个多克隆抗体具有良好的反应性,可为构建树鼩抗感染模型及开展抗病毒作用机制研究等提供技术支持。
4 结论
通过原核表达系统可在大肠杆菌中成功诱导表达获得瑶山亚种树鼩IFN-β和IFN-γ,复性后的融合蛋白具有良好的反应性和免疫原性,免疫小鼠制备获得的多克隆抗体反应性良好且抗体效价高,可作为检测工具用于研究瑶山亚种树鼩病毒感染模型。
猜你喜欢
——和田盘羊
