野鸦椿和圆齿野鸦椿对低温胁迫的生理响应及抗寒性评价
2022-09-05陆思羽涂淑萍姜雪茹
刘 聪,陆思羽,涂淑萍,姜雪茹
(江西农业大学林学院,江西南昌 330045)
0 引言
【研究意义】低温是限制植物地理分布及影响植物生长发育的重要环境因子(Pearce,2001)。野鸦椿[(Thunb.)Dippel]和圆齿野鸦椿(Hayata)均属省沽油科(Staphyleaceae)野鸦椿属()灌木或小乔木,为药赏兼用树种,前者为落叶树种,分布范围广,后者为常绿树种,主要分布在南亚热带。作为观赏树种,圆齿野鸦椿观赏性明显优于野鸦椿,但适宜生长范围小于野鸦椿。圆齿野鸦椿的抗寒性是影响其推广应用的主要因素之一。低温胁迫可引起植物生理代谢紊乱,严重时可导致植株死亡。筛选评价与野鸦椿和圆齿野鸦椿抗寒性相关的生理指标,对培育抗寒品种和扩大生产应用范围具有重要意义。【前人研究进展】植物的抗寒性与抗氧化酶和渗透调节物质关系密切(朱爱民等,2021)。同时,同科同属的不同植物间抗寒性差异也比较明显(于飞等,2014)。黄一波等(2021)在对7种木莲属()植物抗寒性评价及其生理机制的研究中发现,其抗寒性与相对电导率呈显著正相关。谢雨萱等(2021)在研究自然越冬期不同姜花属()植物的抗寒性时发现,抗寒性强的种质叶片其抗氧化物酶(过氧化物酶POD、超氧化物歧化酶SOD)活性和渗透调节物质(可溶性蛋白Pro)含量更高。在有关野鸦椿属抗寒性的研究中,康文娟(2015)研究得出,可溶性糖(SP)和丙二醛(MDA)含量及SOD活性与不同家系圆齿野鸦椿苗木抗寒能力显著相关。也有研究表明,过氧化氢酶(CAT)、POD和SOD等细胞保护酶活性,以及相对电导率(REC)、脯氨酸、MDA、SP和Pro含量等是评价植物抗寒性的重要生理指标(田星等,2018;马晓华等,2021),同时结合主成分分析和隶属函数法对植物抗寒性进行综合评价,能较全面、准确地反映植物间抗寒能力的差异,避免单一指标的片面性(王娜等,2016;李瑞雪等,2017;金龙飞等,2021)。【本研究切入点】目前,对野鸦椿和圆齿野鸦椿抗寒性的研究较少见,尚未见将野鸦椿与圆齿野鸦椿二者进行抗寒性比较的研究。【拟解决的关键问题】比较野鸦椿与圆齿野鸦椿在4 ℃低温条件下处理随时间变化,其叶片Pro、SP、MDA含量,CAT、POD、SOD活性及REC的变化,并利用主成分分析和隶属函数法综合评价二者的抗寒性。筛选出判断野鸦椿与圆齿野鸦椿抗寒性的最佳生理生化指标,并验证二者抗寒性的强弱,为野鸦椿属植物的抗寒性育种提供参考。
1 材料与方法
1.1 试验地概况
试验地设在南昌市青山湖区江西农业大学校内,位于东经115°4948、北纬28°4534,属于亚热带湿润季风气候,温和湿润,降雨充沛,年平均降雨量1600~1700 mm,日照充足。年平均气温17.0~17.7 ℃,气温最高40.9 ℃、最低-15.2 ℃(刘娟等,2018)。
1.2 试验设计
以当年春季播种繁殖的野鸦椿和圆齿野鸦椿盆栽苗为试材。花盆规格为10 cm×10 cm;培养土为园土∶泥炭∶蛭石=3∶1∶1,加少量有机肥。于2019年9月份挑选生长健壮、无病虫危害的野鸦椿和圆齿野鸦椿各120株,置于25±2 ℃的人工气候培养室培养7 d后,剪下长度为20~30 cm的枝梢,先后用自来水和蒸馏水冲洗干净,擦干表面水,剪口处封蜡,然后每10根一扎装入自封袋,置于4 ℃的冰箱中进行低温处理;依次于低温处理的第0、5、10和15 d,每个树种各取出3扎枝梢(3次重复),先取少许叶片测定REC,然后将其余叶片剪下,用锡箔纸包裹后迅速放入液氮中,随后装入自封袋,置于-80 ℃的超低温冰箱中保存,用于生理生化指标的测定。
1.3 测定项目及方法
POD活性采用愈创木酚法测定,CAT活性采用紫外吸收法测定,SOD活性采用氮蓝四唑光化还原法测定(李合生,2000)。MDA含量采用硫代巴比妥酸显色法测定,Pro含量采用考马斯亮蓝法测定,SP含量采用蒽酮比色法测定,REC采用浸泡法测定(孔祥生和易现峰,2008)。
1.4 统计分析
利用Microsoft Excel 2017和Origin 2019b进行数据整理和绘图。利用SPSS 22.0进行多重比较(Duncan’s法)、皮尔逊(Pearson)相关分析及主成分分析。利用模糊数学中的隶属函数法对2个树种的抗寒性进行综合评价。
2 结果与分析
2.1 低温胁迫下野鸦椿和圆齿野鸭椿叶片的SOD、POD和CAT活性变化
2.1.1 叶片SOD活性 由表1可知,野鸦椿与圆齿野鸦椿的叶片SOD活性在低温胁迫下的变化趋势一致,均在4 ℃处理5 d时达最大值,并随着低温胁迫时间的延长,活性逐渐降低;且在相同处理时间下,野鸦椿的叶片SOD活性始终高于圆齿野鸦椿。此外,二者在处理初期(前5 d)叶片SOD活性均上升,随着处理的进行,二者叶片SOD活性均表现出下降趋势,并逐渐与其低温处理0 d时的活性相近,表明此时二者已经对该低温产生适应性。

2.1.2 叶片POD活性 野鸦椿的叶片POD活性整体上高于圆齿野鸦椿,在低温胁迫下,二者POD活性均表现为先升后降。野鸦椿叶片POD活性变化起伏相对较小,最大值出现在低温处理10 d时,之后略有下降;圆齿野鸦椿叶片POD活性变化起伏相对较大,最大值出现在低温处理5 d时,之后大幅下降。
2.1.3 叶片CAT活性 野鸦椿与圆齿野鸦椿叶片CAT活性的变化趋势大致相同,随着处理时间的延长,均表现为先升后降。其中,野鸦椿叶片CAT活性在低温处理5 d时迅速上升,10 d时开始下降;圆齿野鸦椿叶片CAT活性在低温处理5 d时缓慢上升,10 d时开始下降,但下降幅度不显著(>0.05,下同)。野鸦椿CAT活性变化幅度大于圆齿野鸦椿。
综上所述,野鸦椿与圆齿野鸦椿的叶片SOD、POD和CAT活性在低温胁迫下均表现为先升后降的变化趋势。除野鸦椿叶片POD活性在低温处理第10 d达最大值外,其他均在低温处理的第5 d酶活性达最大值。表明2个树种在4 ℃低温处理的初期均产生抵抗低温逆境伤害的反应,此后,随着植物对低温的逐渐适应,酶活性开始逐渐降低。
2.2 低温胁迫下野鸦椿和圆齿野鸭椿叶片SP和Pro含量变化
2.2.1 叶片SP含量 由图1可知,低温处理0 d时,圆齿野鸦椿的叶片SP含量显著高于野鸦椿(<0.05,下同),但在低温处理5 d以后,野鸦椿与圆齿野鸭椿的SP含量差异不显著;且在整个低温处理过程中,二者的SP含量均呈现降—升—降的变化趋势。在处理初期,圆齿野鸦椿叶片SP含量显著下降,而野鸦椿叶片SP含量下降不显著,至低温处理5 d时,圆齿野鸦椿的叶片SP低于野鸦椿。低温处理5~10 d,二者的叶片SP含量均显著升高;低温处理10~15 d时,野鸦椿叶片可SP含量显著降低,而圆齿野鸦椿叶片SP含量降低未达显著水平。

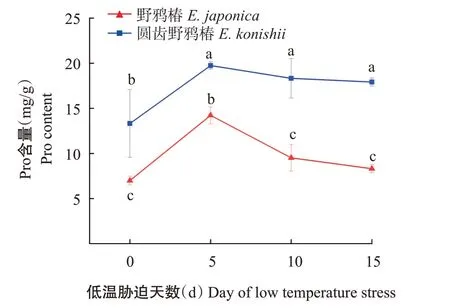
2.2.2 叶片Pro含量 由图2可知,无论是在低温处理前还是处理过程中,圆齿野鸦椿的叶片Pro含量均显著高于野鸦椿。在低温处理初期野鸦椿和圆齿野鸦椿的叶片Pro的含量均显著上升;低温处理5 d以后,野鸦椿叶片Pro含量至低温处理10 d时下降至低温处理0 d的水平并维持,而圆齿野鸦椿Pro含量下降不明显,一直维持在较高水平。
2.3 低温胁迫下野鸦椿和圆齿野鸭椿叶片MDA和REC的变化
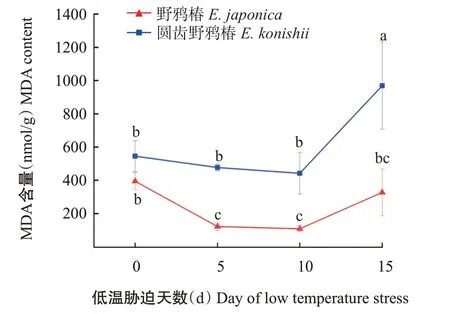
2.3.1 叶片MDA含量 由图3可知,无论是在低温处理前还是在处理过程中,圆齿野鸦椿的叶片MDA含量始终高于野鸦椿。圆齿野鸦椿叶片MDA含量在前10 d的变化幅度不大,10 d之后显著上升;而野鸦椿叶片MDA含量在低温处理初期显著降低,此后变化不大。
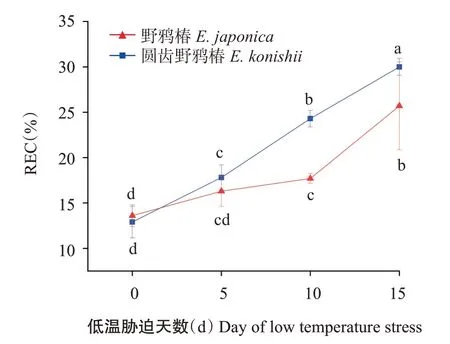
2.3.2 叶片REC 由图4可知,在低温胁迫处理前,2种植物的叶片REC相近,随着低温胁迫天数的增加,2种植物叶片REC均呈稳步上升的趋势,低温处理5 d以后,圆齿野鸦椿的叶片REC上升速度快于野鸦椿,低温胁迫10 d后,圆齿野鸦椿的叶片REC显著高于野鸦椿。
2.4 低温胁迫下野鸦椿和圆齿野鸦椿抗寒性生理指标的相关分析
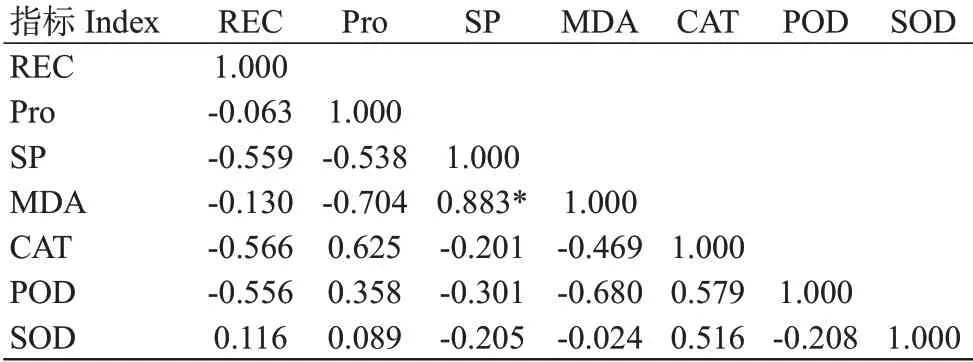
由表2可知,低温胁迫下,MDA含量与SP含量呈显著正相关,其他生理指标间均不显著相关,无法直接通过相关性来判定各生理指标对野鸦椿和圆齿野鸭椿抗寒性的影响。因此要准确筛选出对抗寒性强弱影响较大的生理指标,还需进一步进行主成分分析。
2.5 低温胁迫下野鸦椿与圆齿野鸦椿抗寒性生理指标的主成分分析
选取野鸦椿与圆齿野鸦椿4 ℃低温处理10 d时的7个生理生化指标进行主成分分析,计算其特征值的方差贡献率和累积方差贡献率,并根据主成分的特征值大于1.00的原则提取主成分。由表3可知,前2个综合指标的方差贡献率分别为72.575%和17.235%,累积方差贡献率达89.810%,超过85.000%,基本包含所测指标的全部信息。故取第一主成分(Y)和第二主成分(Y),其余主成分可忽略不计。决定第一主成分的指标主要有POD(0.988)、REC(-0.988)、Pro(-0.936)、MDA(-0.896)、SP(0.864),决定第二主成分的主要是CAT(0.917)。因此,判别野鸦椿与圆齿野鸦椿抗寒性的生理指标及其重要性排序依次为POD=REC>Pro>MDA>SP>CAT。
2.6 野鸦椿和圆齿野鸦椿抗寒性的综合评价
2.6.1 主成分分析法 表3中2个主成分的表达式分别为:


把每个原始指标进行标准化后的数据代入到公式(1)和公式(2)中,可得到野鸦椿与圆齿野鸦椿对应的前2个主成分的值,再分别以2个主成分所对应的贡献率为各自的权重,从而得到基于主成分分析法的野鸦椿与圆齿野鸦椿的抗寒性综合评价模型,即加权值计算公式(陆思羽等,2020)为:

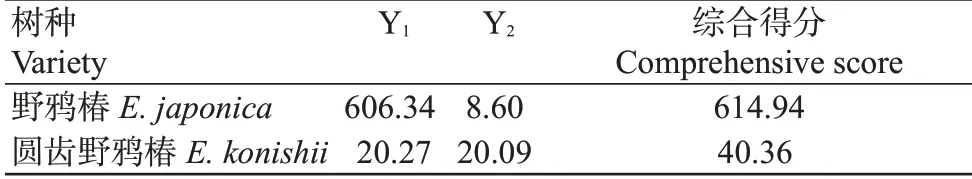
从表4可看出,第一主成分得分较高的为野鸦椿,第二主成分得分较高的为圆齿野鸦椿,而综合主成分得分,野鸦椿(614.94分)远高于圆齿野鸦椿(40.36分),说明野鸦椿抗寒性远强于圆齿野鸦椿。
2.6.2 隶数函数法 利用隶属函数法对主成分分析中提取的6个判别2个树种抗寒性的重要生理指标,即POD和CAT活性、Pro、MDA和SP含量及REC进行综合分析,综合评价野鸦椿与圆齿野鸦椿抗寒性强弱。隶属函数值的计算公式(涂淑萍等,2021)为:

如果某一项指标与抗寒性为负相关,则用隶数函数进行转换,计算公式为:

公式(4)和(5)中,为各处理某一指标的实际测定数值,和分别为该指标实际测定值中最小值和最大值。用模糊数学中的隶属函数方法对2个树种6个指标隶属函数值进行累加,求平均值。隶属函数值越大,抗寒性越强;隶数函数值越小,抗寒性越弱(令凡等,2015)。由表5可知,野鸦椿抗寒性远强于圆齿野鸦椿。

3 讨论
3.1 低温胁迫对野鸦椿和圆齿野鸦椿保护酶活性的影响
POD、CAT和SOD保护酶共同构成植物组织活性氧清除系统,保障植物组织活性氧处于动态平衡,保护膜结构免受低温的破坏,保障植物的抗寒能力(Krasensky-Wrzaczek and Jonak,2012)。本研究中POD、CAT和SOD保护酶呈先升后降的变化趋势,与吴海宁等(2013)的研究结果类似,在低温胁迫初期3种保护酶活性增强对植物体抵御低温寒害起一定的作用(Javadian et al.,2010)。本研究中,除低温处理0 d时,野鸦椿的叶片CAT活性低于圆齿野鸦椿外,其他各时间的胁迫处理野鸦椿的3种叶片酶活性圴高于圆齿野鸦椿。徐小军等(2015)对26份甜瓜()耐冷性相关生理指标的综合研究表明,抗寒性强的品种其保护酶活性亦较强。由此可见,野鸦椿抗性可能较圆齿野鸦椿强。同时,主成分分析表明,判别野鸦椿和圆齿野鸦椿抗寒性的生理指标中,POD的重要性最高,与REC并列,可作为判定野鸦椿和圆齿野鸦椿抗寒性的生理指标;CAT的重要性最低,表明其对野鸦椿和圆齿野鸦椿抗寒性的影响不大,不适合用来作为判定野鸦椿和圆齿野鸦椿抗寒性的生理指标。
3.2 低温胁迫对野鸦椿和圆齿野鸦椿渗透调节物质的影响
在本研究中,圆齿野鸦椿叶片SP含量总体呈先降后升再降的趋势。低温胁迫初期圆齿野鸦椿叶片SP含量的下降,可能是低温胁迫初期,为适应外界环境变化,糖的分解代谢增强所致(王一峰等,2019;刘钰玺等,2020)。在大多数人的研究结果中,如林榕燕等(2021)对秋石斛(spp.)的抗寒性研究及黄丽芳等(2021)对咖啡的抗低温胁迫研究均表明,SP含量随着低温胁迫的进行而升高,且SP含量高的抗寒性强。而在本研究中,5 d后野鸦椿和圆齿野鸭椿叶片SP含量差异并不显著,无法通过SP含量来判断抗寒性强弱。因此,本研究中植物叶片SP含量可能与植物抗寒性关联性不大。主成分分析中,叶片SP含量对判定野鸦椿和圆齿野鸦椿抗寒性的重要性排名也较低。综合研究结果认为,叶片SP含量不适合用作判定植野鸦椿和圆齿野鸦椿抗寒性的生理指标。
本研究中,野鸦椿和圆齿野鸦椿的叶片Pro含量在低温胁迫过程中均表现为先升后降,与谢雨萱等(2021)对自然越冬期不同姜花属植物的抗寒性研究结果类似。低温胁迫初期,Pro含量升高的原因可能是分解了复合蛋白,或增加了具有特殊作用的新蛋白质(王兆,2014)。低温胁迫后期叶片Pro含量降低至胁迫之初的水平,这可能与野鸦椿与圆齿野鸭椿逐渐适应了该低温环境有关。在本研究中,圆齿野鸦椿叶片Pro含量在低温胁迫5 d后有所下降,但下降不明显,基本维持在一个较高的水平,说明该植物在4 ℃的低温条件下产生了逆境胁迫反应。而野鸦椿Pro含量在低温胁迫5 d后出现回落,表明野鸦椿对低温胁迫的适应性强于圆齿野鸦椿。在判别野鸦椿和圆齿野鸦椿抗寒性的生理指标的重要性排序中,叶片Pro仅次于POD和REC,因此适合用作野鸦椿和圆齿野鸦椿抗寒性的判定。
3.3 低温胁迫对野鸦椿和圆齿野鸦椿细胞膜透性的影响
植物的抗寒性越强,在低温条件下受到的损伤越小,MDA含量越低(Gu,1999)。黄丽芳等(2021)研究发现,不同咖啡()叶片中MDA含量的变化幅度与其抗寒性呈负相关。本研究表明,圆齿野鸦椿叶片MDA含量在低温胁迫处理前10 d变化不明显,10 d后显著上升;而野鸦椿叶片MDA含量在低温胁迫初期显著下降,之后其变化幅度较小,且野鸦椿的MDA含量始终低于圆齿野鸦椿。由此可见,野鸦椿的抗寒性比圆齿野鸦椿更强。在主成分分析中,MDA对野鸦椿和圆齿野鸦椿的抗寒性判定贡献率较高,可用作筛选野鸦椿和圆齿野鸦椿抗寒性强弱的生理指标。
本研究表明,野鸦椿和圆齿野鸦椿叶片REC均随低温胁迫时间的延长而升高,与王摇宁等(2020)对樟()的研究及李呈呈等(2021)对彩叶桂(‘Caiye Gui’)的研究结果一致;而郑素兰等(2020)研究发现,姜荷花()叶片REC与抗寒性呈反比,抗寒性较强的种类其REC低于抗寒性较弱的种类。本研究中野鸦椿的REC始终低于圆齿野鸦椿,说明野鸦椿的抗寒性优于圆齿野鸦椿。因此,在判别野鸦椿和圆齿野鸦椿的抗寒性生理指标重要性排序中,REC和POD并列第一,可用于野鸦椿和圆齿野鸦椿的抗寒育种筛选。
植物的抗寒性受多种因素及指标影响。主成分分析是考察多个变量间相关性的一种多元统计方法,可从原始变量中导出少数几个主成分,尽可能多地保留原始变量的信息,且彼此间互不相关作为新的综合指标(范宗民等,2020);运用隶属函数法能较为科学地评价植物的抗寒性(陈小凤等,2017;李瑞雪等,2017)。本研究利用主成分分析和隶属函数法进行综合评定,得出结果为野鸦椿幼苗抗寒性较圆齿野鸦椿更强;细胞膜透性对野鸦椿和圆齿野鸦椿的抗寒性强弱影响最大,其次是保护酶活性,渗透调节物质对二者的抗寒性强弱影响相对较小。
4 结论
POD、REC、Pro和MDA可用于野鸦椿和圆齿野鸦椿抗寒育种筛选,其中POD和REC对2个树种的抗寒性强弱影响较大。此外,野鸦椿幼苗抗寒性远强于圆齿野鸦椿,可考虑将野鸦椿作为砧木对圆齿野鸦椿进行嫁接,提高圆齿野鸦椿的抗寒性。
