马尾松幼苗根系和光合碳供应对土壤性质变化和温室气体排放的驱动作用
2022-08-31赵泽尧赵天宇
赵泽尧,张 雪,陈 桐,赵天宇,徐 帅,梅 莉,*
1 华中农业大学园艺林学学院,湖北省林业信息工程技术研究中心,武汉 430070 2 襄阳市自然资源和规划局,襄阳 441003
温室气体排放导致的全球气候变化问题是当前人类面临的最严峻课题之一。森林土壤是温室气体的重要来源,主要包括土壤自养呼吸和异养呼吸排放的二氧化碳(CO2)[1]以及土壤硝化反硝化作用产生的氧化亚氮(N2O)[2]。研究表明,森林植被遭受自然灾害、氮沉降、人为活动及土地利用方式改变等干扰,导致森林土壤温室气体排放量增加[3]。如:森林采伐及更新等经营措施,可通过改变林分光照强度、土壤温湿度、土壤有机质分解速率、土壤理化性质等直接或间接地影响土壤温室气体排放[4—7]。森林土壤呼吸排放CO2受植被类型、经营方式、环境温湿度、土壤养分状况及土壤碳储量等多种环境因子影响[8],一直是森林生态系统研究的热点与难点问题[9]。N2O的排放也受土壤环境因子的调控,如土壤温度、水分、pH值和土壤活性碳氮等[10—11],森林破坏导致的土壤环境因子变化,进一步影响了土壤N2O排放通量[12—13]。由于野外环境的复杂性,当前研究中关于不同干扰类型导致的CO2及N2O等温室气体排放变化的研究结果及其机理解释不尽一致,有待进一步研究。
无论森林植被遭受了哪种干扰,其直接作用都将表现在输入森林土壤的根系数量/生物量和组成发生变化[14—15],以及向地下部分的光合碳供应变化。例如:气候变暖导致细根生产量增加和周转加快[16],从而使输入到土壤的活细根和死细根量都在不断增加;大气N沉降及森林经营中施用N肥[17]等因素提高林地土壤氮含量,导致细根生长、死亡和周转率发生相应的变化[18],进而影响根系输入量和组成变化;森林植被采伐、火灾、病虫害等状况也会导致森林土壤中活根生物量降低、死细根数量在短期内快速增加,光合碳向地下供应急剧下降。因此,弄清输入土壤的细根生物量及光合碳供应变化对森林土壤理化性质、微生物及温室气体排放的影响,可以从不同干扰类型下导致的根系输入和光合碳供应变化上预测和解释其对土壤温室气体排放的影响及其机理,这对于预测全球森林植被干扰变化背景下的森林温室气体排放通量变化及其生态功能具有重要意义。
马尾松(Pinusmassoniana)是我国南方最重要的造林树种之一,当前面临严峻的松材线虫(Bursaphelenchusxylophilus)危害,受害后林的根系死亡、地上光合碳供应降低,研究受灾林分土壤理化和生物环境以及土壤温室气体排放的变化是预测灾后森林生态功能的重要依据。为此,本研究采用盆栽模拟试验方法,通过不同栽植密度控制根系输入量、通过环割和截干模拟光合碳供应降低,模拟研究灾后根系输入量和光合碳供应降低对土壤特性及土壤呼吸的影响及其驱动机制,为灾后马尾松林的科学经营提供依据。
1 材料与方法
1.1 试验材料与处理
2016年10月,将湖北省太子山林场管理局苗圃培育的2年生马尾松播种容器苗移栽于华中农业大学校内温室。栽培容器规格:上口直径275 mm,下底直径180 mm,高200 mm。栽培基质为等体积的河沙、蛭石、泥炭土、黄泥土均匀混合物,基质全氮含量(2.39±0.09)g/kg,全磷含量(0.56±0.01)g/kg,全钾含量(6.43±0.26)g/kg,有机质含量(64.32±4.33)g/kg。分别1株/盆和3株/盆栽植,2017年6月处理时,3株/盆栽植的平均苗高(59.03±1.11)cm和地径(5.7±0.06)mm均显著低于1株/盆的苗高(65.08±0.54)cm和地径(8.5±0.12)mm,但3株/盆的每盆苗木总叶、茎、根及总生物量均显著大于1株/盆。
2017年6月,选取长势基本一致的马尾松幼苗,在距地面5 cm高处分别进行环割和截干处理,环割宽度1 cm。共6个处理:1株/盆对照(SCK),1株/盆环割(SG),1株/盆截干(SJ),3株/盆对照(TCK),3株/盆环割(TG),3株/盆截干(TJ),每个处理30盆重复,共计180盆。每个处理随机选取5盆安装静态气室,用于土壤温室气体取样;每次每个处理破坏性取5盆重复,用于土壤和植株理化性质分析等。
1.2 植物和土壤样品采集与分析
环割和截干处理后,每隔1周取植物和土壤样品。取样时将花盆倒扣取出整棵植物,避免破坏根系结构。植物样品分根、茎、叶分别取样,取样后用微波炉高火5 min灭活,以避免根系呼吸造成碳损耗。用Epson(Expression 10000XL)数字扫描仪对灭活后的根系进行扫描,扫描的图片用根系形态分析软件(WinRhizo 2004b)分析根系长度、总表面积以及总体积。然后于65 ℃烘干、称重、粉碎,过0.5 mm(40目)筛,用于测定根、茎、叶的非结构性碳水化合物(Total nonstructural carbohydrate,TNC)和全氮(N)含量。
植物样品TNC含量测定:称取植株各部分粉末各 0.1000 g,采用蒽酮比色法分别测定其可溶性糖和淀粉含量,根据葡萄糖标准曲线换算为相应的百分比含量(%),可溶性糖与淀粉含量之和为TNC含量[19]。
植物全氮含量采用凯氏定氮法测定:称取0.1000 g植物样品(0.5 mm,40目筛)装入50 mL消化管的底部,加蒸馏水2—3滴润湿,用移液器加入浓硫酸5 mL,轻轻摇匀,静置过夜后消煮,消煮过程中间断性加入6滴H2O2溶液直至消煮液由黑色变为无色透明,冷却后用全自动凯氏定氮仪测定(SHN220N石墨消解仪,K9860全自动凯氏定氮仪)。
土壤样品测定:植株取出后,将土壤样品混匀,过2 mm土壤筛,土壤pH值采用水浸提电位法测定;土壤铵态氮与硝态氮用流动注射分析仪(ICP-MS)进行测定;土壤微生物量碳(Soil microbial biomass carbon, SMBC)和土壤微生物量氮(Soil microbial biomass nitrogen, SMBN)采用氯仿熏蒸氯化钾浸提,用TOC仪进行测定[20];土壤群落结构采用磷脂脂肪酸方法(Phospholopid Fatty Acid, PLFA)进行测定和分析。
1.3 土壤温室气体排放测定
土壤CO2和N2O排放通量采用静态箱-气象色谱法进行测定[21]。静态箱由聚氯乙烯(PVC)管制成,周围土壤紧密压实防止漏气,于2017年6月18日—8月18日期间取样,于8:00—11:00 am密封PVC管,培养2 h后取气,将取出的气体装入真空瓶带回实验室,立即用气相色谱仪(Agilent 7890A)测定其中CO2、N2O的浓度,并计算得出土壤CO2、N2O的排放通量。
土壤CO2、N2O排放通量计算方法:F=ρ×V/M×Δc/Δt×273/T×α。
式中,F为CO2、N2O的排放通量,正值为排放,负值为吸收;ρ为标准状况下气体的密度,CO2、N2O的密度分别为1.978 kg/m3,1.98 kg/m3,V是采样箱体积(m3),M为采样箱底部土壤干重(g);△c/△t为在特定时间内的气体浓度变化速率;T为采样点的绝对温度;α分别为N2O换算到N(28/44)、CO2换算到C(12/44)的转换系数。
1.4 数据处理与统计分析
应用SPSS 19.0软件进行单因素方差分析,当方差分析结果表现为显著性差异时,通过Tukey检验进行不同处理间的多重比较,用Pearson法对土壤呼吸CO2、N2O与各影响因子进行相关性分析,用Origin 8.0软件绘图。
2 结果与分析
2.1 马尾松幼苗生物量和根系形态结构及养分特征
通过采用单株/盆和3株/盆种植控制根系输入量,3株/盆种植的苗木叶、茎及根系总生物量均显著高于单株/盆种植(表1)。无论是单株/盆还是3株/盆种植,环割处理30 d后,苗木地上部分生物量均没有显著变化,而根系生物量比对照显著降低,其中单株/盆和3株/盆根系生物量分别比对照降低47.98%和15.64%;分别比处理开始时降低51.75%和9.65%。截干处理30 d后,根系生物量也显著降低,其中单株/盆和3株/盆根系生物量分别比对照降低40.59%和44.87%;分别比处理开始时降低44.89%和40.96%。环割处理30 d后,单株/盆种植的根系总根长、表面积及体积均显著下降;而3株/盆种植的根系与对照无显著差异。无论是单株/盆还是3株/盆种植,截干处理30 d后的根系总根长、表面积及体积均比对照显著降低(表1)。
3株/盆种植的根系TNC含量略低于单株/盆,但差异不显著;而3株/盆种植的土壤氮含量显著低于单株/盆(图1,表2)。环割处理后,单株/盆种植的根系TNC在第7 d开始显著下降,之后一直低于对照;根系氮含量在第7 d显著高于对照,之后与对照无显著差异;3株/盆种植的根系TNC在处理14 d后显著下降,而根系氮含量与对照没有显著差异。截干处理后,单株/盆和3株/盆种植的根系TNC均在处理14 d后显著下降,而单株/盆种植的根系氮含量在处理后第7 d显著高于对照,3株/盆种植的根系氮含量在处理21 d和30 d后显著上升(图1)。

图1 不同处理苗木根系非结构性碳水化合物和氮含量(n=5)Fig.1 Root total nonstructural carbohydrate and nitrogen contents in seedling roots of different treatment (n=5)
2.2 根系输入量和光合碳供应对土壤化学性质与微生物群落结构的影响
3株/盆种植比单株/盆土壤铵态氮和硝态氮含量低(表2)。环割和截干处理30 d后,土壤pH值有升高趋势,其中单株/盆截干处理的土壤pH显著提高。无论是单株/盆还是3株/盆种植,环割和截干处理的土壤硝态氮含量均比对照显著增加,而土壤铵态氮与对照没有显著差异(表2)。环割和截干处理后,SMBC含量均低于对照,其中单株/盆环割和截干及3株/盆环割均显著低于对照(P<0.05); 环割和截干处理后,SMBN含量均高于对照,其中单株/盆环割和截干及3株/盆环割与对照有显著差异(P<0.05)(图2)。

表2 土壤pH、硝态氮、铵态氮含量(均值±标准误)(n=5)
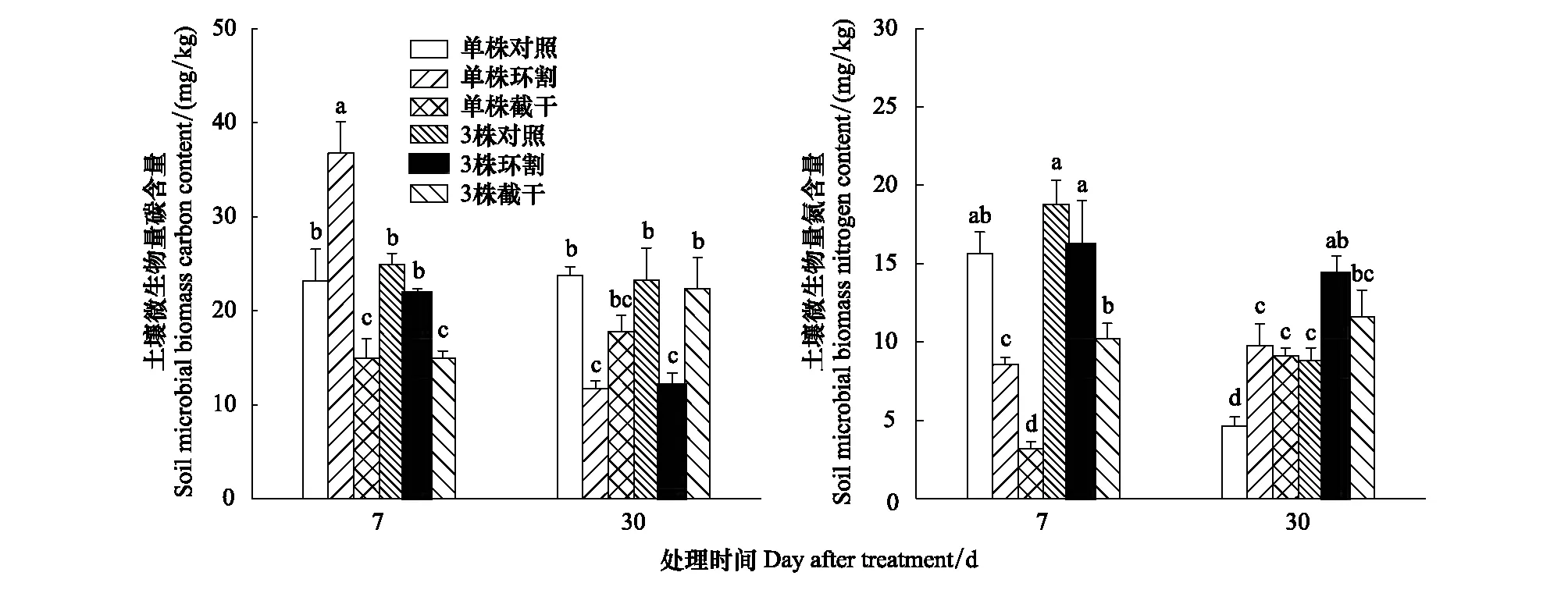
图2 不同处理土壤微生物量碳和土壤微生物量氮含量 (n=5)Fig.2 Soil microbial biomass carbon and soil microbial biomass nitrogen contents of different treatments (n=5)
土壤菌群中,细菌多样性最高,其次是放线菌、真核生物和真菌。总细菌、厌氧菌、放线菌、AM菌根真菌和真核生物多样性分别占菌群总量的68%、1.15%、14.64%、3.4%和7.1%。处理7 d时,根系输入量对土壤各微生物组成没有显著影响;处理30 d时,根系生物量高的3株/盆种植的革兰氏阳性菌、厌氧菌、放线菌及丛枝菌根真菌丰富度均比单株/盆显著增加(表3)。
无论是环割还是截干处理,土壤中各种微生物与对照相比均有下降趋势,其中单株/盆环割和单株/盆截干的真菌丰富度比对照显著下降;3株/盆种植的细菌总丰富度(包括革兰氏阴性菌及革兰氏阳性菌)、放线菌及丛枝菌根真菌均比对照显著下降(表3)。
环割和截干处理后,土壤中细菌和真菌结构发生了显著变化。细菌总量比对照显著下降,但细菌多样性占菌群总量的比例显著提高,在单株/盆环割及截干处理30 d时均显著高于对照;而真菌多样性占菌群总量的比例下降,处理30 d时,单株/盆环割和截干处理的真菌比例均显著低于对照;但3株/盆环割和截干处理与对照之间没有显著性差异(表3)。
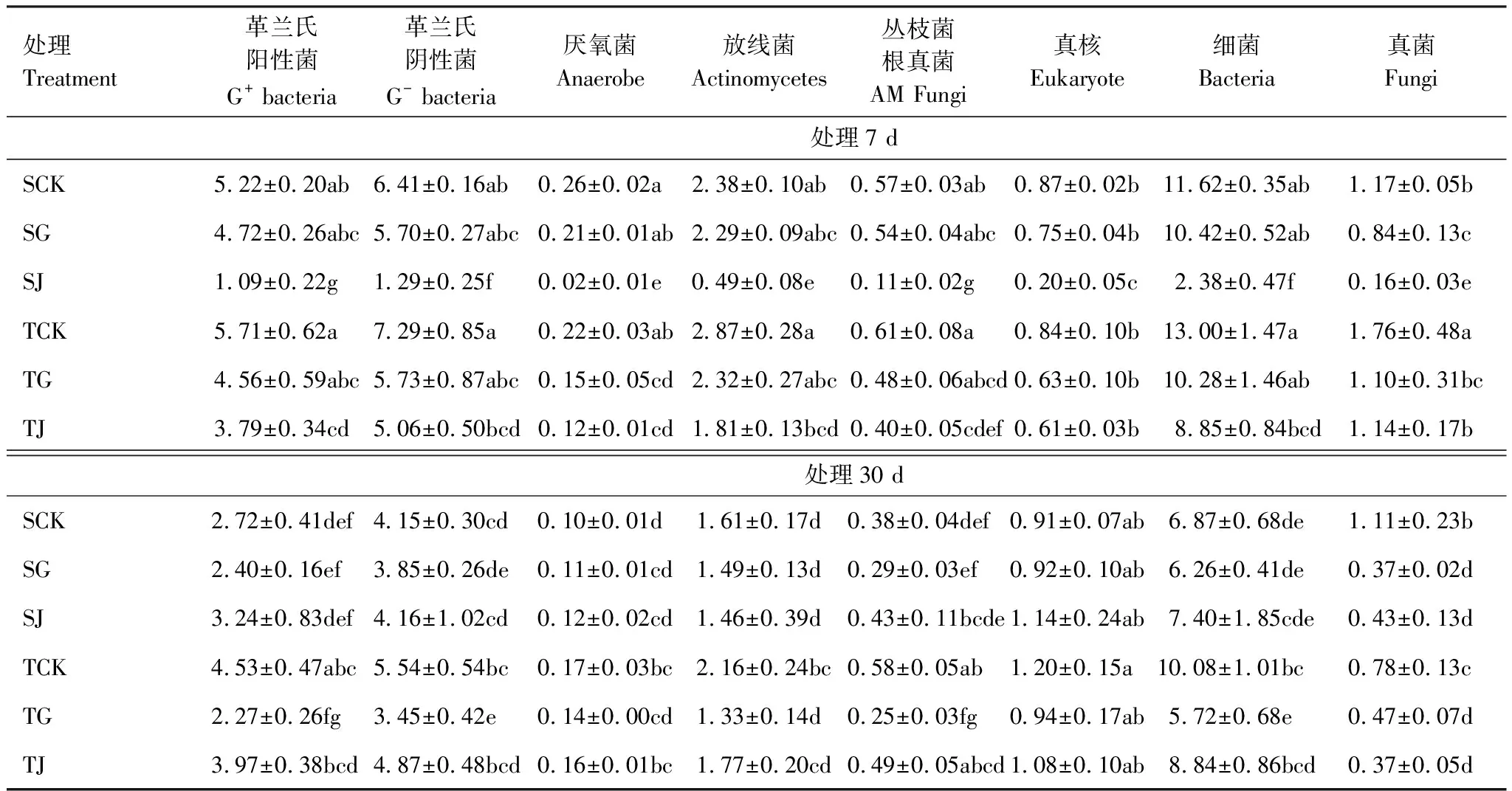
表3 不同处理土壤微生物的特征PLFAs含量(平均值±标准误)(nmol/g)(n=5)
方差分析表明,真核生物与真菌含量没有显著相关性,但其他各个微生物组成之间均呈极显著正相关关系。不同处理时间、根系输入量及光合碳供应对土壤细菌和真菌含量均有显著影响(表4)。土壤细菌含量与根系生物量、SMBC和SMBN显著正相关;土壤真菌含量与土壤温度显著负相关,与根系生物量、SMBC和SMBN显著正相关(表5)。

表4 根系输入量、处理时间及光合碳供应对土壤微生物群落结构的影响方差分析
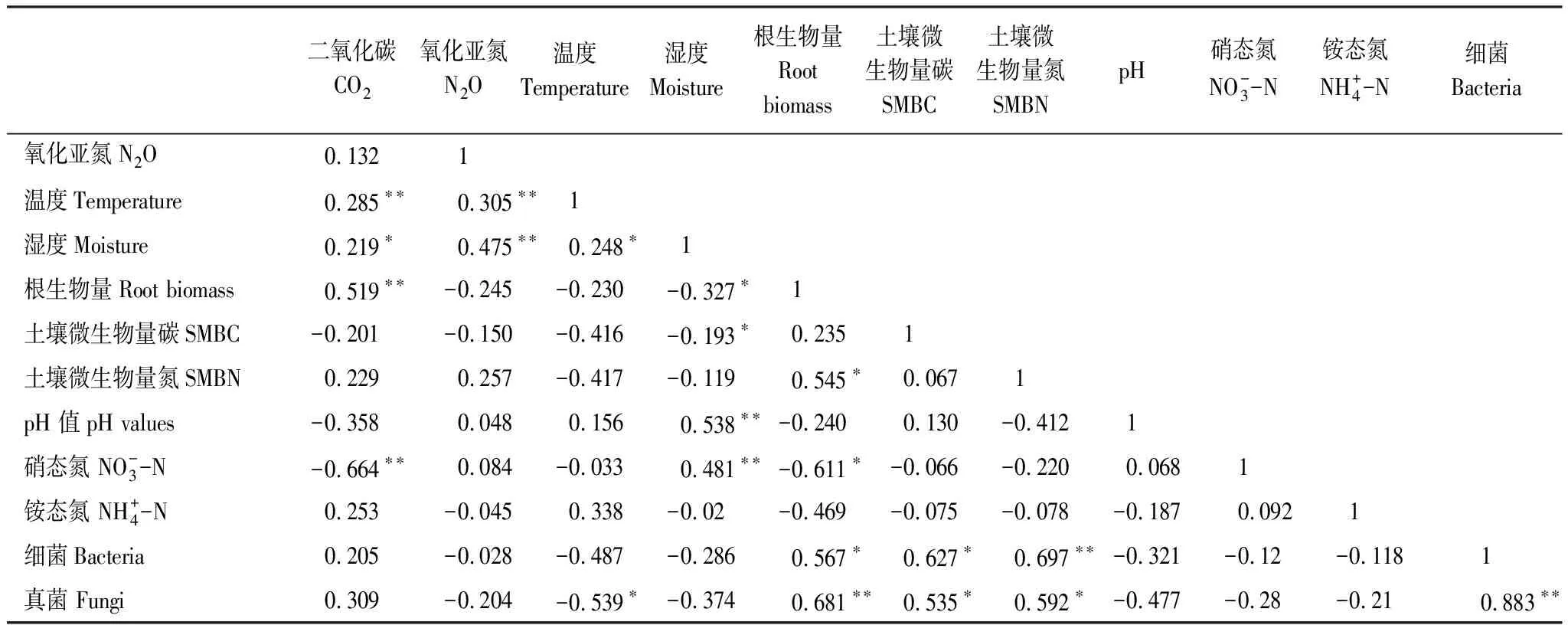
表5 土壤CO2和N2O排放通量与土壤环境因子的相互关系
2.3 根系输入及光合碳供应对土壤温室气体排放的影响
环割和截干处理后,土壤呼吸排放的CO2均比对照显著下降,在处理后的第7天、18天和26天,CO2排放速率达到峰值且差异尤其显著(图3)。环割和截干对单株/盆种植土壤CO2平均排放量影响(平均分别降低66.1%和59.2%)大于对3株/盆种植的影响(平均分别降低35.3%和41.6%)。单株/盆处理后的第5、9、11、13天,环割处理的CO2排放量显著低于截干处理。3株/盆处理后的第5、15天,环割处理的CO2排放量显著低于截干处理,而在第3、18、30、45天,环割处理的CO2排放量显著高于截干处理,其余时间没有显著差异(图3)。
环割和截干处理后,土壤排放的N2O均比对照显著提高,在处理后的第5天、18天和27天峰值时,差异尤其显著。其中环割处理下,单株/盆和3株/盆种植的N2O平均排放量分别比对照提高63.2%和75.3%;而截干处理后,单株/盆和3株/盆种植的N2O平均排放量别是对照的110倍和82倍,分别是环割的68倍和47倍(图3)。

图3 不同处理土壤CO2和N2O的排放通量Fig.3 Soil CO2 and N2O fluxes of different treatments
相关分析表明,土壤CO2排放通量与土壤温度、土壤湿度及根系生物量显著正相关,与土壤硝态氮含量显著负相关;土壤N2O排放通量与土壤温度和土壤湿度显著正相关,与根系生物量负相关,但差异不显著(表5)。
3 讨论
3.1 根系输入及光合碳供应对SMBC、SMBN含量的影响
SMBC和SMBN是衡量土壤微生物动态变化的关键性因素。本研究中增加活根系输入量对SMBC无显著影响,但显著增加了SMBN的含量,可能与根系生物量的增加提高了土壤微生物的活性有关,而根系的分解会进一步增加SMBN含量[22]。3株截干和环割降低了SMBC含量,但SMBN含量呈现先下降后上升的趋势。根系的死亡首先导致根际微生物活性降低,然后根系死亡增加土壤有机碳会导致土壤微生物活性增加[23]。环割和截干30 d后土壤SMBN含量增加,这加强了土壤有机碳的分解,随着有机碳的逐渐减少,可能会减弱土壤微生物的活性,进而降低SMBC的含量[24]。环割和截干后,土壤氮含量的增加,提高了土壤氮素循环微生物所需的底物,底物的增加进一步导致微生物活性变强、SMBN含量提高[25]。
3.2 根系输入及光合碳供应对土壤微生物群落结构的影响
土壤微生物种群、群落结构、微生物生物量等对环境变化非常敏感,可以反映土壤质量、土壤肥力的演变,并可用作评价土壤健康的生物指标[26]。细根对土壤微生物群落结构具有重要影响[27],研究表明,去除根系降低阔叶林和针叶林土壤微生物生物量[28],降低人工林土壤微生物的碳源代谢功能[29]。本研究利用PLFA技术分析土壤微生物群落结构发现,根系输入量高的3株/盆处理,其土壤革兰氏阳性菌、厌氧菌及丛枝菌根真菌均显著高于单株,这与前人的研究结果一致[27—28]。相关分析表明,活根系输入量与土壤真菌和细菌极显著相关(表5),进一步明确了根系能够促进土壤微生物群落丰富度。
通过环割阻断地上光合碳供应能改变土壤理化性质,并对土壤微生物群落结构产生重要影响[29—30]。本研究结果进一步表明,截干和环割后,微生物群落各组分的含量均有下降趋势,其中单株/盆种植下降快于3株/盆种植,截干处理下降速度快于环割。取样过程中观察到截干后植物根系死亡的速度比环割快,这可能是截干后微生物组分含量下降较快的原因。根系所产生的根际沉积和根凋落物为土壤微生物的重要碳源[31],但根系对细菌和真菌的影响不同,真菌更喜欢来源于根系的碳[29],环割和截干后活根的死亡导致真菌群落显著下降;细菌群落结构的下降可能是活根生物量下降间接作用的结果,因为细菌只有在无可替代碳源的情况下,才加强对源于根的碳利用[32]。因此,截干和环割后根系的死亡改变了根际周围的微生物活性以及根际分泌物[25],可能是降低微生物群落各组分含量的主要因素。Zhou等[33]进行大数据分析表明,土壤微生物主要通过调整群落结构而不是调整生物量来适应森林抚育间伐后导致的土壤生物和非生物变化。
3.3 根系输入量对温室气体排放的影响
根系呼吸是土壤自养呼吸的重要组分,约占土壤呼吸的10%—90%[34]。相关研究表明,根系生物量与土壤呼吸速率呈正相关关系,而去除根系则使森林土壤呼吸速率显著降低[35—37]。本研究中提高栽植密度(3株/盆)增加活根系输入量导致土壤湿度降低(表5),间接影响了土壤CO2排放通量,根系生物量与土壤呼吸呈显著正相关关系。虽然根系输入量增加能够提高土壤自养呼吸,但同时因蒸腾作用及根系对土壤水分的吸收利用差异而降低了土壤湿度,土壤湿度变化导致的呼吸降低部分抵消了因根系生物量引起的呼吸增加分。根系输入量的差异,影响生物因子、小气候和土壤理化性质之间复杂的相互作用和联系,进而通过微生物影响土壤呼吸[38]。
根系输入量与土壤N2O排放通量没有显著性相关(图3、表5),由相关分析得出土壤N2O排放速率与土壤温度、土壤湿度具有极显著的正相关关系(表5),这与菊花等[39]研究结果相一致。因根系生物量的差异,土壤N2O排放通量与根系生物量负相关,可能是较多的根系吸收利用了较多的土壤硝态氮,进而显著降低了土壤速效氮含量(表2),降低了土壤N2O排放的底物。
由于对地上地下资源的竞争作用,3株/盆种植的马尾松苗根系TNC及氮含量均显著低于单株/盆种植(图1),这种根系质量的变化可能在一定程度上削减了由于根系生物量增加而导致的土壤CO2和N2O排放量增加。Volder等[40]研究发现根系氮含量与土壤呼吸速率存在极显著的相关性。因此,在评价影响森林生态系统土壤温室气体排放因素时,不仅要考虑根系输入量的变化,还应该考虑根系质量的差异[41]。
3.4 光合碳供应对土壤温室气体排放的影响
截干与环割光合碳供应终止,降低了土壤CO2的呼吸速率,这与Laporte等[42]的研究中择伐及间伐降低土壤CO2排放速率的结果一致。土壤自养型呼吸主要来源于植物根系和根际微生物,植物光合作用所产生的底物是其主要来源[43—44];异养型呼吸主要来源于土壤微生物和土壤动物,土壤碳储量是异养呼吸的主要来源[45]。截干与环割降低了土壤CO2的呼吸速率,截干与环割处理阻止了地上向根系运输碳水化合物,降低了根系的非结构性碳含量,根系活性降低,因而减弱了土壤呼吸的自养呼吸部分;另一方面,活根系的减少伴随的土壤微生物群落丰富度降低[46],降低了土壤的异养呼吸部分,也是土壤呼吸速率减弱的重要原因之一。相关性分析表明,土壤CO2排放通量与土壤湿度呈显著正相关关系,截干与环割处理导致根系吸收水分的速率下降,土壤湿度变大,但截干与环割处理土壤呼吸速率却没有因土壤湿度增加而升高。相对与土壤湿度变化的影响,本研究中根系死亡导致的自养呼吸减弱以及土壤微生物异养呼吸的减少对土壤呼吸的影响具有主导作用。
截干与环割处理终止光合碳供应,增加了土壤N2O的排放速率,截干与环割处理直接增加了死根输入量,增加了土壤微生物的分解底物,加快了土壤N2O的排放速率。另一方面土壤N2O排放速率与土壤湿度呈极显著正相关关系,这与Amadi等[47]的研究结果相类似,本研究中截干与环割处理降低了根系对土壤水分的吸收,一定程度的土壤湿度增加能促进硝化作用,进而提高土壤的N2O排放速率。
4 结论
通过不同栽植密度、环割和截干处理,模拟根系输入量及地上光合碳供应变化对土壤理化性质、土壤微生物及温室气体释放的影响。结果表明,根系输入增加能显著增加土壤微生物丰富度,提高土壤CO2排放速率,但对土壤N2O的影响不显著。地上光合碳供应终止后,SMBC含量降低,SMBN增加,土壤中各种微生物组成丰富度均随之下降,土壤CO2排放速率显著下降,土壤N2O排放速率则显著提高。研究进一步明确了森林土壤温室气体排放量受土壤理化性质和微生物群落多样性的影响,森林土壤CO2排放通量与活根系输入量及地上光合碳供应呈正相关,N2O排放通量主要受土壤温度和湿度影响。当森林植被遭受干扰破坏时,森林土壤温室气体排放受根系输入量及地上光合碳供应变化共同影响。
