油蒿叶片资源利用效率变化及其影响因素
2022-08-31姜晓燕李鑫豪魏宁宁高圣杰查天山
蒋 燕,靳 川,姜晓燕,李鑫豪,魏宁宁,高圣杰,刘 鹏,贾 昕,3,查天山,3,*
1 北京林业大学水土保持学院,北京 1000832 宁夏盐池毛乌素沙地生态系统国家定位观测研究站,盐池 7515003 北京林业大学水土保持国家林业局重点实验室,北京 100083
植物生长需要从环境中获取资源(即光合同化所需的光、水和氮),光合作用是植物获取并利用各种资源的重要过程,容易受到限制[1—3]。气候变化增加了全球极端天气的发生频率和强度,尤其是生态环境脆弱的干旱半干旱地区,可能会强烈影响该地区的资源供给,从而导致植物光合和生产能力发生变化。
植物资源利用效率(RUEs)作为连接光合作用和资源供给的关键纽带,被广泛用于理解植物对气候变化和极端气候的响应[4—5]。其中,叶片光利用效率(LUE)、水分利用效率(WUE)及氮利用效率(NUE)等是决定植物适应环境变化,协调自身碳水关系的关键指示[6]。光、氮、水等资源利用效率间的权衡(即相对变化)决定着植物最大碳吸收能力[7—8]。但植物资源利用效率权衡关系因研究环境和时间尺度的差异而不同[4,9],了解干旱区植物叶片光、氮、水利用效率相对变化特征及其对环境的响应,有助于深入理解植物对气候变化的响应,准确预测植物碳吸收能力[4,10]。
环境变化是植物叶片资源利用效率变化的主要及直接驱动因子(即非生物因子),其通过改变光、水和氮资源供给和需求对植物资源利用效率产生影响[11]。因此直接选择环境中的光、水和氮为代表性非生物因子进行分析。植物通过叶片性状间的协调和权衡来适应环境波动(即生物因子),以实现叶片资源利用和生态适应策略的优化[3,12]。在资源梯度不同的生境中,植物叶片性状发生了不同程度的适应性分化[13—14]。比叶面积(SLA)和叶片氮含量(LNC)是描述植物叶片性状的基本参数,其变化反映了不同物种在长期进化过程中对资源的利用和生存环境的适应特征[13,15],因此选择SLA和LNC为代表性生物因子进行分析。综合研究生物与非生物因子对植物叶片RUEs的影响机制,能更好地理解干旱地区植物对环境变化的适应过程。毛乌素沙地是我国西北地区典型的干旱半干旱地区,降水有限且不规律。油蒿(Artemisiaordosica)是该地区半干旱灌丛生态系统中最常见和优势的灌木种之一。为此本研究以油蒿为研究对象,利用LI- 6400XT便携式光合仪(LI-COR, Lincoln, USA)进行油蒿叶片光合参数原位测定,计算其LUE、WUE和NUE,结合同步监测的环境因子和测定的叶片SLA和LNC,分析油蒿叶片生长季RUEs的动态变化及其与生物和非生物因子的关系, 探明油蒿对毛乌素沙地干旱环境的适应机制。旨在探究: (1)油蒿叶片资源利用效率相对变化特征;(2)生物和非生物因素对油蒿叶片资源利用效率的影响机制,为阐明荒漠生态系统对环境变化的响应机制提供理论依据。
1 材料与方法
1.1 研究地概况及样地布设
宁夏盐池毛乌素沙地生态系统国家定位监测研究站(37°04′ N—38°10′ N,106°30′ E—107°47′ E,海拔1550 m)处于毛乌素沙地南缘,占地总面积2000 hm2。该区属于典型的中温带大陆性季风气候。年平均气温为8.1℃,降水稀少,年降水量292 mm,主要集中在夏季,年际变化大;年潜在蒸散量为2100 mm,无霜期约为120 d。典型植物群落为油蒿群落,优势建群种为油蒿,夹杂有老瓜头(Cynanchumkomarovii)、杨柴(Hedysarummongolicum)、赖草(Leymussecalinus)等其他物种,并伴有苔藓地衣结皮。本文选取油蒿为研究对象,生长季集中在5—9月,5月为春季,6—8月为夏季,9月为秋季。
于2017—2019年生长季通过野外调查选择油蒿分布集中区域作为研究样地,样地内灌木平均株高0.5—1.0 m,样地大小为15 m×15 m,在样地内随机选择3个5 m×5 m样方,每个样方内选择3株油蒿标记。观测期内样地中油蒿植被覆盖度为45%,平均冠幅和株高分别为0.86和0.78 m。在每个样方中选择长势较好、且植株之间遮挡较少的油蒿作为样株,为避免边缘效应影响,植株尽量选择在样方内部位置,每次生理实验在所标记的植株上进行,测量周期为7—10 d。
1.2 测量方法1.2.1 光合特征参数
在已标记油蒿样株上分别选取一簇1—2年生小枝上的成熟叶片,依据枝条和叶片生长状况,以及叶片大小选择相似叶片,之后对所选叶片进行测量。考虑植物光合随天气变化存在差异的特点,选择晴朗无风的天气,于9:00—12:00使用Li- 6400XT便携式光合仪透明叶室进行气体交换参数的测定,当测量数值小数点后一位稳定时,每个叶片记录20次读数,主要包括单位叶面积的净光合速率(Pn,μmol m-2s-1)、蒸腾速率(E,mmol m-2s-1)和气孔导度(Gs,mol m-2s-1)。
在每次气体交换参数测定时,同时利用LI- 6400XT便携式光合系统,配备LED红蓝光源叶室,进行光响应曲线测量(A-PAR曲线)。进行光响应曲线的测定前先对目标叶片进行充分诱导;叶室温度设置为25℃,CO2浓度设置为400 μmolCO2mol-1,近似环境中的浓度,相对湿度设置为55%左右。光合有效辐射(PAR)梯度共设置12个点,分别为2000、1800、1500、1200、1000、800、600、400、200、100、50和0 μmol m-2s-1。测定时应在设置的每个PAR下适应3 min测定其光合速率。测定完成后通过模型拟合,利用A-PAR曲线估算叶片光最大净光合速率(Pmax,μmol m-2s-1)和内禀量子效率(α,mol/mol)[16]。叶片的光合特征参数和响应曲线测量时间为上午9点至12点[17—18]。每次测量的叶片面积小于叶室面积,在每次测量完之后取下叶片带回实验室,使用YMJ-A托普叶面积仪测定实验样叶面积,然后对所测数据进行校准。
1.2.2植物叶片性状特征
每次气体交换测量结束后,在与标记油蒿特征相似的相邻植株上立即采集10—20片样叶,用于测量叶面积和氮含量。叶面积测量完毕后,用电子天平测量饱和鲜重,75℃烘干48 h后测定干重,比叶面积(SLA,cm2/g)为鲜叶面积与干重的比值。将每次烘干后的样叶粉碎、研磨后制成粉样,利用凯氏定氮法测定样叶的氮含量(LNC,g/kg)[8]。
1.2.3环境因子
光合有效辐射(PAR)、降水量(PPT)和30 cm土壤含水量(SWC)由样地附近的微气象观测系统监测。PAR由光合有效辐射传感器(PAR-LITE, Kipp & Zonen, the Netherlands)测得;ECH2O土壤温湿度仪(ECH2O- 5TE, Decagon Devices, USA)用于测量SWC;翻斗式雨量筒(TE- 525 WS, Campbell Scientific, USA)完成样地内PPT监测。PPT由CR200数据采集器(Campbell Scientific Inc.,USA)获取,PAR和SWC由CR3000数据采集器(Campbell Scientific Inc.,USA)获取。所有环境因子测定间隔均为30 min。每一实验周期内,在每个样方周围用土钻法取15、30、45 cm土壤样品各1份,制成土样后,凯氏定氮法测定土样的全氮含量(Nsoil,g/kg)。
1.3 资源利用效率计算
直角双曲线修正模型可以更好地反应植物净光合速率对光合有效辐射的响应过程[16],采用此模型拟合植物光响应曲线。模型表达式为:
(1)
式中,Pn(μmol m-2s-1)为净光合速率,PAR为光合有效辐射,α是光响应曲线的初始斜率,定义为植物内禀量子效率,反映植物利用光能的最大潜能,所以LUE (mol/mol) 由α值表示;β为修正系数;系数γ为内禀量子效率与植物最大光合速率之比;Rd(μmol m-2s-1)为PAR=0时的暗呼吸速率。
对式(1)两边求导数,得到植物的量子效率,即有:
(2)
当式(2)等于0时,可求出植物的饱和光照强度(LSP):
(3)
与植物饱和光照强度对应的最大光合速率Pmax(μmol m-2s-1)为:
(4)
则WUE (μmol/mol) =Pmax/Gs,表示植物的内在水分利用效率,其反映了叶片碳吸收和水分耗散的内在调控情况。
植物单位叶面积氮含量Narea=LNC/SLA,NUE (μmol g-1s-1) =Pmax/Narea,表示植物的光合氮利用效率。
1.4 数据分析
采用方差分析(ANOVA)对不同季节间的叶片光、水和氮利用效率进行比较。变异系数(CV)用于量化叶片资源利用效率、叶片性状和环境因子的季节变化。用分段平均方法将WUE和NUE数据按LUE分组平均(LUE间隔0.002 mol/mol),又将NUE数据按WUE分组平均(WUE间隔0.2 μmol/mol),每组求平均值和标准误差,通过标准化主轴分析法(SMA,smatr包中的R函数SMA)检验油蒿LUE、WUE和NUE间的相互关系。线性回归和非线性回归分析叶片资源利用效率和叶片性状特征及环境因子之间的相关关系。全子集回归法获取最优模型,以描述叶片资源利用效率随各影响因素的变化情况。同时采用结构方程模型(SEM)进一步评估叶片性状特征及环境因子对叶片资源利用效率的直接和间接影响。所有数据分析和作图通过RStudio 4.0.5完成。
2 结果与分析
2.1 环境因子与叶片性状变化特征2.1.1 环境因子季节变化特征
由图1和表1可知,三年PAR季节变化趋势一致,2017—2019年年均值分别为425.93,398.86 μmol m-2s-1和393.37 μmol m-2s-1,年际变异幅度极小(CV=3.5%)。2017—2019年Nsoil年均值分别为0.13,0.17 g/kg和0.06 g/kg,年际波动较大(CV=37.88%),波动范围是0.042—0.33 g/kg,Nsoil基本在秋季达最高值。生长季降雨量主要集中在7—9月(约72%),约占全年降雨量的60%,生长季总降雨量最高的年份是2017年(299 mm),比总降雨量最低的2018年(220 mm)高35.36% (表1)。SWC主要受降雨量影响,和降雨量季节分布一致,春季最低,夏季最高,秋季次之,2017年SWC最高(0.112 m3/m3),2018年最低(0.093 m3/m3)。
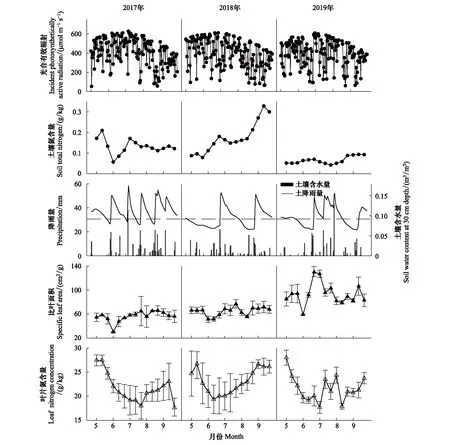
图1 研究地点2017—2019年生长季环境因子和叶片性状季节动态Fig.1 Seasonal variation in environmental factors including incident photosynthetically active radiation (PAR), soil total nitrogen (Nsoil), soil water content at 30 cm depth (SWC) and precipitation (PPT), and leaf traits including specific leaf area (SLA) and leaf nitrogen concentration (LNC) in the study site over growing season (May—September) during 2017—2019图中水平虚线表示SWC=0.09 m3/m3

表1 2017—2019年生长季环境因子均值(总和)及误差表
2.1.2油蒿SLA和LNC季节变化特征油蒿SLA和LNC随季节发生变化(图1),SLA变化幅度显著高于LNC,LNC相对稳定(SLA:CV=29%;LNC:CV=13%)。SLA和LNC日均值3年内波动范围分别是30.20—129.88 cm2/g和17.61—28.03 g/kg。SLA没有显著的季节规律性,随着叶片的生长,SLA在夏季波动更为明显,2017—2019年年均值分别为62.86,64.63 cm2/g和93.48 cm2/g。春秋季LNC高于夏季,三年年均值分别为21.65,23.08 g/kg和21.63 g/kg。
2.2 油蒿叶片资源利用效率季节变化特征
油蒿叶片资源利用效率在生长季呈现明显的季节和年际波动(P<0.01) (图2)。三年内LUE,WUE和NUE日均值波动范围分别是0.018—0.153 mol/mol,31.82—192.25 μmol/mol和3.01—16.07 μmol g-1s-1;WUE波动幅度最高(CV=48%),NUE最低(CV=39%)。LUE和WUE年均值在2017年最高,分别为0.079 mol/mol和74.69 μmol/mol;WUE和NUE年均值在2018年最低(图3)。

图2 油蒿叶片光、水和氮资源利用效率季节变化特征Fig.2 Seasonal variations in leaf-level resource use efficiencies including light use efficiency (LUE), water use efficiency (WUE) and nitrogen use efficiency (NUE) in A. ordosica
除2018年LUE在秋季最高,三年内油蒿叶片资源利用效率月均值基本在夏季出现高峰值(图2);WUE和NUE分别在2017年7月和2019年8月达最高,为104.02 μmol/mol和11.49 μmol g-1s-1,LUE在2017年6月达最高(0.12 mol/mol)。叶片资源利用效率最低值分布在生长季各时期,无规律性。研究期间油蒿WUE和LUE,WUE和NUE关系不显著(P>0.05;图4),WUE不受LUE和NUE的影响。但LUE和NUE正相关性显著(P<0.01,R2=0.17;图4),LUE和NUE的季节变化趋势基本一致,且每年最低值出现时间一致(图2)。
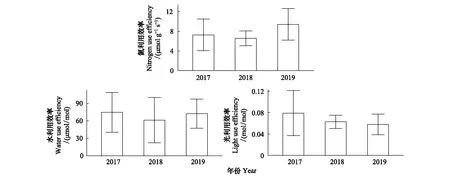
图3 油蒿叶片光、水和氮资源利用效率年均值变化特征Fig.3 Variations in annual mean of leaf-level resource use efficiencies including light use efficiency (LUE), water use efficiency (WUE) and nitrogen use efficiency (NUE) in A. ordosica
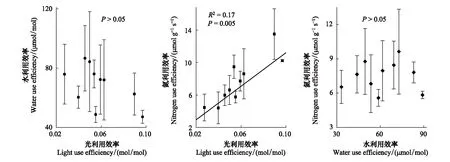
图4 油蒿叶片资源利用效率SMA回归分析Fig.4 Relationships between leaf-level resource use efficiencies (i.e., WUE, LUE, and NUE) during the growing season based on SMA regression in A. ordosicaSMA:标准化主轴分析法 Standardized major axis
2.3 油蒿叶片资源利用效率与环境因子的关系
SWC>0.09 m3/m3时,油蒿叶片资源利用效率变化不受其影响;SWC<0.09 m3/m3,WUE和SWC的关系为二次函数,拟合结果决定系数较高(R2=0.41),LUE和NUE不受SWC影响。资源利用效率对Nsoil和PAR的响应受SWC调节,SWC>0.09 m3/m3时,除LUE随Nsoil增加呈线性降低外(P<0.01),资源利用效率变化均不受Nsoil和PAR影响;SWC<0.09 m3/m3,WUE和Nsoil的关系为二次函数,有74%的结果可以被该方程解释,NUE和PAR呈倒数关系(P=0.02;图5)。
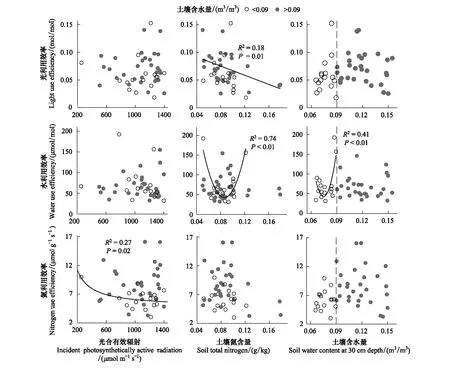
图5 油蒿叶片资源利用效率对光、水和氮资源的响应Fig.5 Relationships between leaf-level resource use efficiencies and incident photosynthetically active radiation (PAR), soil total nitrogen (Nsoil), soil water content at 30 cm depth (SWC) in A. ordosica图中垂直虚线表示SWC=0.09 m3/m3
2.4 油蒿叶片资源利用效率与SLA和LNC的关系
油蒿叶片资源利用效率与LNC无显著相关性;LUE随SLA增高而降低(P<0.01),NUE则随SLA增高而升高(P<0.01),WUE对SLA的响应不明显(图6)。由油蒿叶片资源利用效率最佳回归模型可知:SWC、Nsoil和SLA能很好地预测LUE和NUE(表2,LUE:R2=0.26,P<0.01;NUE:R2=0.32,P<0.001),但无法预测WUE。SEM模型也表明LUE和NUE变化受SWC、Nsoil、SLA和LNC综合影响;模型分别解释了LUE和NUE变化的31.3%和40.2%(图7)。SLA(λ=-0.38)和Nsoil(λ=-0.32)直接降低LUE,Nsoil(λ=0.18)还通过SLA间接降低LUE,LNC(λ=-0.19)通过SLA间接提高LUE,由SEM得到的标准化总效应表明,LUE主要受SLA和Nsoil影响,其次是SWC和LNC。SLA(λ=0.43)和SWC(λ=0.31)直接提高NUE,SWC(λ=0.05)还通过SLA间接提高NUE,Nsoil(λ=-0.38)和LNC(λ=-0.21)直接降低NUE,但Nsoil通过SLA和LNC间接提高NUE,由图7可知,NUE主要受SLA和SWC影响,其次是Nsoil和LNC。但对于WUE,也无法用所选指标得出拟合优度较好的SEM模型。
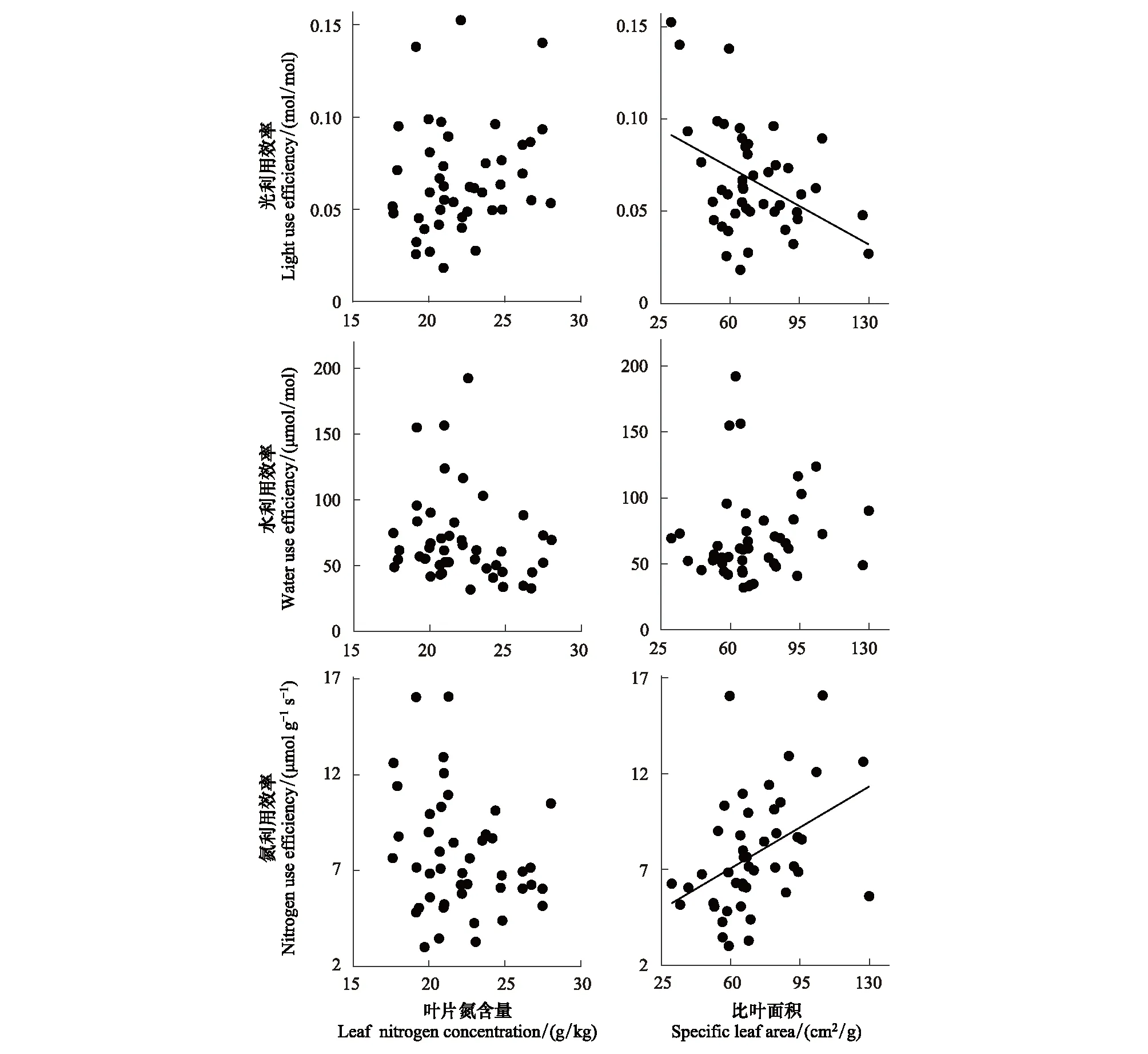
图6 油蒿叶片资源利用效率对比叶面积和叶片氮含量的响应Fig.6 Relationships between leaf-level resource use efficiencies and leaf nitrogen content (LNC), specific leaf area (SLA) in A. ordosica

图7 环境因子和叶片性状影响资源利用效率动态的结构方程模型和标准化总效应Fig.7 Structural equation modelling and standardized total effect showing the effect of environmental factors and leaf traits on resource use efficiencies in A. ordosicaLUE:光利用效率 Light resource use efficiency;NUE:氮利用效率 Nitrogen resource use efficiency;PAR:光合有效辐射 Incident photosynthetically active radiation;Nsoil:土壤全氮含量 Soil total nitrogen;SWC:30cm土壤含水量 Soil water content at 30-cm depth;LNC:叶片氮含量 Leaf nitrogen content;SLA:比叶面积 Specific leaf area。 箭头旁边的数字是标准化路径系数λ,实线箭头和虚线箭头分别代表模型中的正负效应,R2表示有关变量的共同解释度;LUE:χ2=0.51,df=2,P=0.80,RMSEA<0.001;NUE:χ2=0.59,df=1,P=0.44,RMSEA<0.001
3 讨论
3.1 油蒿叶片资源利用效率变化特征及权衡关系
很多研究表明叶片光、水和氮等资源利用效率季节差异明显,植物在生长季各阶段有不同的资源利用策略[4,19]。本研究中油蒿基本在夏季叶片光、水和氮利用效率最大(图2)。且本研究地附近其他灌木,如沙柳和杨柴的光和水利用效率也在夏季最高[20—21]。研究地夏季干旱时常出现,植物在水分亏缺时提高了自身的光合生产能力,即提高资源利用效率,加快物候更换,使得资源利用效率基本在夏季最高,在干旱限制来临前尽快完成生活史[22]。但也有研究指出温带地区乔木树种,如云杉、山松、白杨和椴树氮利用效率在夏季最高,光利用效率在春季最高,水利用效率在夏季最低[19,23—24]。这可能是由于不同地区环境限制条件不同导致,也有可能是灌木与乔木间的资源利用策略差异,表明环境因子可能是资源利用策略差异的直接驱动因子,物种的生物特性是决定因素[11,25]。

表2 油蒿叶片资源利用效率最佳回归模型
植物最大化碳吸收或利用效率理论取决于光、氮、水利用效率的权衡[7—8]。且生态经济理论认为植物会降低丰富资源的利用效率,最大化最有限资源的利用效率,从而增强碳同化能力[26]。Tarvainen等发现在针叶林混交林中,NUE和WUE、LUE负相关显著[19];Gong等对内蒙古干旱草原优势草本植物的研究也指出植物最大化水分利用效率时,氮利用效率降低[27],与该理论一致。但本研究发现3年期间油蒿光和氮利用效率正相关性显著,二者的变化趋同,且都与水利用效率无显著相关性(图4),研究结论与此理论存在差异。研究地光照充足,光资源一直处于过量状态,油蒿LUE主要受本身羧化能力的影响,而叶氮的增加会直接影响羧化酶的含量和活性,进一步增加CO2的同化速率[6],LUE由此提高,NUE也随着投入叶绿体氮含量的增加而提高[28],从而LUE和NUE显著正相关。蒸腾与PAR和Narea间无明显的关系可能是导致WUE与LUE和NUE没有显著相关性的主要原因。这与李鑫豪等对油蒿短期监测所得的光、水和氮利用效率权衡关系也不一致[6]。可能是由于时间的积累降低了饱和资源的相对重要性,引起长期和短期资源利用效率权衡存在差异[9]。所以有必要对灌木资源利用效率权衡机制进行更加深入地长期研究,增强对灌木碳同化能力的理解。
3.2 油蒿叶片资源利用效率对环境的响应
水分是影响干旱区生态系统功能和植物生长的主要环境因子,同时调控其他因子的响应[18,29—32],与本文研究结果一致。当干旱出现时(SWC<0.09 m3/m3)[33],油蒿水利用效率升高,油蒿在干旱时通过气孔限制防止叶片失水以保持正常生理活动,气孔导度降低,导致蒸腾的下降幅度大于光合,WUE升高[17,34],但WUE在严重干旱(SWC<0.075 m3/m3)时降低,因为在严重干旱胁迫时,植物光合作用主要受叶片内部CO2扩散和代谢限制,不再是气孔限制[35];水资源充足时(SWC>0.09 m3/m3),WUE变化不受其影响(图5)。WUE不存在最优模型(表2;图7),可能是由于不同水平SWC对WUE有着不同的直接调节作用(图5)。光、氮利用效率对SWC的响应不明显,但对Nsoil和PAR的响应受SWC调控(图5、图7;表2)。干旱诱导的气孔关闭和叶面积减少往往会抑制半干旱生态系统冠层光合作用,在不同时间尺度上掩盖和调节PAR对植物的影响[31];SWC影响该系统的Nsoil,且植物各器官氮吸收与根系土壤水吸收密切相关[36];因此SWC除自身对植物的影响外,同时调控其他环境因子对植物的影响,共同限制植物叶气孔、酶活性及叶根结构等生理生化及形态反应[30],导致叶片资源利用效率发生变化,影响植物生存生长,证实了现有的研究结论[12,21,28—29,33]。
水资源充足时(SWC>0.09 m3/m3)时,油蒿资源利用效率基本不受光、水和氮的影响(图5),表明即使光、氮资源不够充足,若水分能满足植物蒸腾、细胞扩张和代谢过程[29],植物仍能正常获取和利用环境中的资源维持自身生长。2017年降雨量最高,油蒿LUE和WUE在该年最高也为该结论提供了依据(图3)。但LUE在水分充足时随Nsoil上升而降低(图5)。油蒿通过根系吸收的氮随Nsoil增加而增加,在水分富足状态下,叶片将有更多的氮投资于叶绿体中以增加CO2同化速率,油蒿在较低光强时易达到光饱和点[28],研究地光照充足,通常超过油蒿光饱和点的光强,其通过光系统Ⅱ热耗散调节机制耗散过剩光能,甚至强光使得光抑制现象出现[20],导致油蒿光能利用能力降低。SWC<0.09 m3/m3时,油蒿受干旱胁迫,NUE随PAR升高而下降,降到一定程度后趋于平稳。首先光合作用会随PAR升高而增强,但受到干旱胁迫的植物会将叶片中更多的氮投资于保卫构造以防止失水过多和强光伤害,分配给光合系统的氮减少,导致光合可利用的氮减少,氮利用效率降低[8,28];其次当PAR达到或超过油蒿光饱和点时,光合速率和氮分配基本不变,NUE趋于稳定。WUE同时受氮水限制(图5)。因此油蒿WUE受干旱胁迫时对环境响应敏感,变化复杂(图7;表2),需要在进一步研究中明确干旱区物种WUE对环境的响应机制,探寻典型干旱植物对干旱环境胁迫的响应及其适应策略。
3.3 油蒿叶片资源利用效率与叶片性状特征关系分析
植物通过叶片功能性状的变异,以改变资源获得和保存之间的权衡关系来适应环境变化[3,12,33],获得其最适生态策略位点[12,14]。夏季降雨量最高,SWC随之升高,高SWC使得油蒿从土壤中获取更多的氮水。此时正值油蒿完全展叶期,生理活动旺盛,充足的养分促进了叶片细胞的伸长,体积增大,叶片密度下降,SLA增大,对资源的获取能力增强[13,37],油蒿消耗更多的光、水和氮资源进行光合同化,以满足自身生长,是光氮利用效率在夏季最高的主要原因之一。SEM结果也表明SWC和Nsoil通过SLA和LNC间接影响LUE和NUE,证实了其他学者的研究结果[8,12,28]。
SLA可以反映植物获取资源的能力,主要反映了植物对碳获取与利用的平衡关系,被认为是指示植物养分利用策略的关键指标[38],我们的研究表明油蒿LUE和NUE主要受SLA调控(图7),支持了现有的研究结果。SLA和LUE呈显著负相关,和NUE呈显著正相关(图6)。高比叶面积一般叶组织密度较小,叶片较薄[39—40],使得叶片内部水分向叶表面扩散的距离或阻力减小,增加了植物内部水分散失[41],强光与水分胁迫并存会打破叶绿体内光合作用固定CO2和吸收光能的平衡,导致植物叶片光利用效率下降。同时SLA和LNC负相关关系明显(图7),表明高SLA有利于降低叶片建成成本,用于构建防护结构的氮较少,则分配较多的氮于类囊体和RuBP羧化酶中使其具有较高的光合能力[42],氮利用效率升高[6],油蒿NUE和SLA变化趋势高度一致也验证了NUE随SLA增加这一点。
LNC对油蒿叶片资源利用效率的影响并不显著(图6),可能是由于LNC在生长季相对稳定(CV=13%),植物资源利用主要由其光合能力决定[19]。但有研究指出LNC对资源利用效率的影响较为复杂,与本文略有不同[28,43],主要原因是植物叶氮分配的复杂性和干旱对氮分配的影响,叶片中的氮一般分配在叶片各细胞结构及游离化合物中,分配到叶绿体中的氮才会直接决定光合效率的高低[44],且研究地植物易受干旱胁迫,水分胁迫下叶绿体膨胀,排列紊乱,类囊体层肿胀或解体,叶氮的分配倾向及叶绿体中氮的利用受到干扰[44],从而扰乱了叶氮对植物资源利用效率的影响。因此未来应该着重研究叶片氮分配比例的变化,以期更好地了解干旱地区叶片氮对植物资源利用的影响。
4 结论
三年内油蒿叶片光、水和氮利用效率基本在夏季最高,最低值分布在生长季各时期,无规律性;叶片光、氮利用效率同增同减(P<0.01),但叶片水利用效率变化与光、氮利用效率无显著关系。水分和土壤氮是影响油蒿叶片光、水和氮利用效率季节和年际变异的主要非生物因子;油蒿通过调控比叶面积改变其对环境中光、水和氮资源的利用效率,以适应环境变化和保持正常的生理活动。叶片氮含量对叶片资源利用效率无明显影响,未来的研究应侧重于细化叶片氮分配对植物叶片资源利用效率的影响。研究结果为植物-气候反馈机制的探索提供了基础,对了解干旱地区植物对气候变化的适应能力具有重要意义。
