川中丘陵区冬水田种植模式转旱作土壤呼吸组分特征及碳平衡
2022-08-31岳雪萍吕天星周高子贾媛丽任万军
岳雪萍,王 淘,吕天星,周 涛,刘 琦,唐 磊,周高子,贾媛丽,周 伟,任万军,陈 勇,*
1 四川农业大学农学院/作物生理生态及栽培四川省重点实验室, 温江 611130 2 浙江大学农业与生物技术学院/农药与环境毒理研究所, 杭州 310012
作为陆地生态系统最活跃的碳库,农业土壤碳库对全球碳循环的影响十分重要[1]。农田生态系统与大气的二氧化碳交换过程是影响大气二氧化碳浓度的关键过程[2]。土壤呼吸是土壤碳库以CO2形式回归大气碳库的主要途径,占陆地生态系统与大气之间碳交换总量的2/3,对气候变化和碳循环起着不可忽视的重要作用,土壤呼吸强度的微小变化可能对大气中二氧化碳的浓度产生很大影响[3]。土壤呼吸(Rs)是由土壤产生并向大气中释放CO2的过程,主要包括异养呼吸(Ra)和自养呼吸(Rh)两个组分,前者主要来自于土壤动物和微生物呼吸,后者属于根源呼吸[4—5]。精确地测定陆地生态系统土壤呼吸及其组分特征,对于揭示大气“失踪的碳汇”之谜以及寻找土壤固碳增汇的措施具有重要的指导意义[6]。研究表明,自养呼吸对土壤呼吸的贡献可达到50%—65%,但其并非土壤自身的碳损失,在核算土壤碳平衡时一般扣除自养呼吸[6—7]。自养呼吸由植物光合产物控制,异养呼吸则由土壤有机质控制,它们各自呼吸所利用的碳源不同,对土壤呼吸的贡献、对全球变化的作用也不同[8]。土壤呼吸是农田碳平衡研究的基础[9],农田系统碳平衡常采用净生态系统生产力(NEP)衡量,即净初级生态系统生产力(NPP)与土壤异养呼吸碳释放量(Rh)的差值[10—11]。种植模式改变、作物配置不同、养分管理措施改变等均影响农田土壤碳平衡[12—15]。因此,通过土壤呼吸组分测定并分析系统碳平衡对理解系统固碳增汇具有重要的意义。
冬水田-水稻种植模式是川中丘陵区传统的以蓄水为中心的种植制度。随着农业水利工程和机械化的发展,该种植模式逐渐演变为水旱轮作或旱作多熟种植模式,提高了周年生产效益[16]。研究表明[17],冬水田是重要的CO2排放源,具有很大的温室气体减排空间。由于环境剧烈变化,冬水田-水稻种植模式转变为旱作过程的土壤呼吸及组分会受到极大影响,但其具体排放特征仍不明确。为了探索冬水田-水稻模式转变为旱作过程中作物季及季后休闲期土壤呼吸组分及其影响因素,本研究设置冬水田-水稻、冬水田-水稻转为旱作玉米、冬闲—玉米3种种植模式的试验,探明冬水田-水稻转旱作过程中作物生长季及季后休闲期土壤呼吸的变化特征和组分贡献,并阐明温度敏感性,以期为冬水田种植模式转变后的农田生态系统的土壤碳平衡评估提供理论依据。
1 材料与方法
1.1 研究区域概况
研究地点位于四川省成都市金堂县转龙镇大桥村(30°40′N,104°45′E),该区属于亚热带季风性温润气候,年均降雨量900—1300 mm,年均日照1042—1412 h,年均温度16℃,土壤属于潜育水稻土。该区属于典型川中丘陵区,立体气候明显,形成了冬水田-水稻、油菜—水稻、冬闲田—玉米、马铃薯—玉米等多元化种植模式。试验期间当地气象资料如图1,其中,平均气温18.2℃,最低气温出现在12月,为0.6℃,最高气温出现在8月,为36.9℃,累积降雨量859.7 mm,集中分布在7—8月。

图1 试验点气温和降雨量分布Fig.1 Temperature and precipitation distribution in the experimental site
1.2 试验设计
试验于2019年4月20日至2019年12月31日期间开展,其中4月20日至9月16日为作物季,9月16日至12月31日为作物季后休闲期。试验设置冬水田-水稻(FR)、冬闲田—玉米(FM)、冬水田-水稻转旱作玉米(FTD)3种种植模式,形成水水连作、旱旱连作、水作转旱作3种类型,单因素随机区组设计,3次重复,共9个小区,小区面积为58.5 m2(6.5 m×9.0 m)。试验开始前,当地农户已经在FR模式田块上实行了20年以上的冬水田-水稻种植模式,FM田块为附近(<50 m)实行了5年以上的冬闲田—玉米种植模式,FTD种植模式为冬水田-水稻种植模式下从稻季开始转旱作玉米。本研究的作物季为3种模式的玉米和水稻生长期,玉米和水稻收获后为季后休闲期,不种植其它作物,由于本试验中FTD模式次年种植马铃薯,为了研究冬水田转旱作利用的水旱转换过程,本研究设定3种种植模式的季后休闲期均截止于当年12月31日。三个处理的作物的播期和播栽密度见表1,耕作及管理措施见表2。

表1 各作物播栽期和播栽密度
1.3 气体样品采集与测定
采用根排除法和密闭气室-气相色谱法测定土壤呼吸及其组分,2019年6月10日开始,每隔10d选择晴天(遇雨适当延后)进行土壤呼吸测定,取样时间为上午9:00—11:00。取样装置由顶箱和底座构成,规格根据各作物行穴距设置,采集水稻土壤呼吸的顶箱和底座长宽规格为20 cm×40 cm,采集玉米土壤呼吸的顶箱和底座长宽规格为30 cm×40 cm,底座的高度设置为5 cm和30 cm,其中底座高度为5 cm的采样箱用于测定土壤总呼吸(Rs),底座高度为30 cm的采样箱用测定异养呼吸(Rh),顶箱由亚克力板制成,高度为20 cm,顶端安装1个小型风扇、温度计插口和气体样品取样接口。
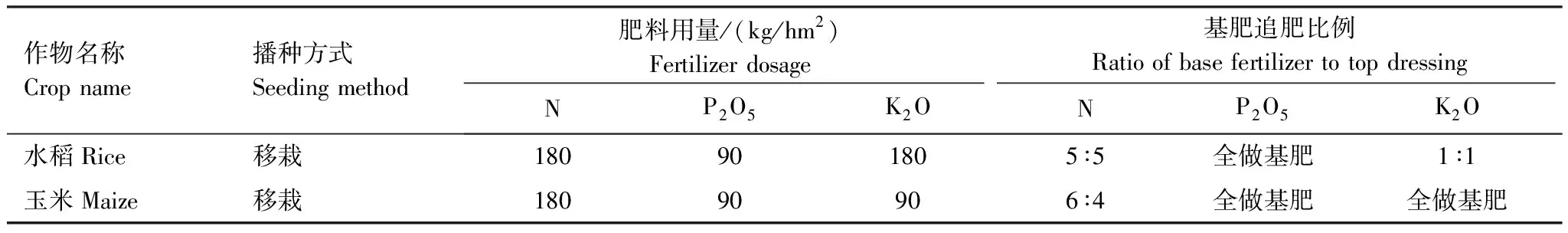
表2 各作物施肥和管理措施
采样装置的底座于水稻和玉米移栽当天安置于行间,并用泥土对外部进行敷边防止呼吸外溢,整个采样期间底座均不移动。采样时底座水槽内加水以密封顶箱和底座之间的空隙,并打开风扇使采样箱内气体充分混合。顶箱扣入底座后立刻用20 mL 注射器采集第1次样品,然后每10 min采集1次,共采集4次,每次采集气体样品20 mL。每次取样前一天,去除底座内土壤表层的一切活体,为了避免由于采样时间差异而导致的系统误差,每次采样时三个重复同时进行。采样后立即将样品带回实验室,使用日本岛津气相色谱仪(2010 Plus)测定CO2浓度。
1.4 土壤呼吸速率计算
土壤呼吸速率计算公式[18]如下:
(1)
式中,F为土壤呼吸速率,mg m-2h-1;ρ为标准大气压下的CO2密度,为1.97 kg/m3;T为采样过程中取样箱内的平均温度,℃;H是取样箱内的净高度,m;dC/dt是采样气体浓度的变化率。自养呼吸计算公式[19]如下:
Ra=Rs-Rh
(2)
式中,Rs为土壤总呼吸,Ra为自养呼吸,Rh为异养呼吸。
土壤呼吸累积排放量计算公式[20]如下:
(3)
式中,中,M为土壤呼吸累积排放量(kg/hm2);F为土壤呼吸速率(mg m-2h-1);i为第i次测定;(ti+1-ti)为相邻两次测定间隔天数。
1.5 水热生态因子测定
于每次土壤呼吸气体取样时,同步测定土壤温度、箱内气温,有水层期间测定箱内水层高度和水温。土壤温度、箱内气温和水温均采用便携式温度测定仪(MITIR-TP678)测定,土壤温度分0—5 cm和5—10 cm分别测定。田间水层高度用直尺测量。
1.6 温度敏感系数(Q10)计算
土壤呼吸速率与土壤温度之间的关系采用指数模型拟合[21],公式如下:
R=aebt
(4)
式中,R为土壤呼吸速率(mg m-2h-1),t为土壤温度(℃),a是温度为0℃时的土壤呼吸速率,b为温度反应系数。温度敏感系数(Q10)计算公式如下:
Q10=e10b
(5)
式中,b同上式(4),Q10指温度升高10℃时土壤呼吸速率变化的倍数,该指标可以衡量土壤呼吸速率对温度的响应程度。
1.7 作物生长季的碳平衡测算
采用净生态系统生产力(NEP)衡量生态系统碳平衡[14]。其计算公式如下:
NEP=NPP-Rh
(6)
式中,NPP为净初级生态系统生产力,是作物全生育期地上部碳积累量与地下部碳积累量的总和,常采用成熟期作物地上部和根的生物量来计算NPP,作物地上部各器官和根的碳含量一般取45%,即作物光合过程中合成1 g有机质需要吸收0.45 g C[22]。土壤异养呼吸碳排放量(Rh)为各处理下排除根系的土壤异养呼吸在整个作物生育期间CO2的累积排放量。NEP为正值时,表示该系统是大气CO2的吸收汇;反之为大气CO2的排放源。
1.8 数据处理
采用SPSS13.0 进行数据处理,LSD法进行多重比较,origin 2020进行作图。
2 结果分析
2.1 不同种植模式下土壤总呼吸通量特征
通过对作物季和季后休闲期农田土壤呼吸速率的连续测定,结果表明3种种植模式下农田土壤总呼吸速率在作物生长季和季后休闲期具有较大差异。在作物生长季,冬水田-水稻转旱作模式(FTD)和冬水田-玉米模式(FM)的土壤呼吸速率均表现为单峰变动趋势,峰值出现在玉米籽粒灌浆期(7月下旬—8月中旬),冬水田-水稻模式(FR)土壤呼吸速率总体维持在较低的排放水平(图2)。

图2 不同种植模式下作物季及季后休闲期土壤呼吸速率变化Fig.2 Changes of soil respiration rate in crop season and fallow period under different planting patternsFR:冬水田-水稻 Paddy field-rice;FM:冬闲田-玉米 Fallow field-maize;FTD:冬水田-水稻转旱作 Paddy field-rice to upland cropping
从3种种植模式土壤呼吸速率比较来看,6月20日前,土壤呼吸速率表现为FM>FTD>FR,6月20日—6月30日,FM的土壤呼吸速率升高,FTD次之,FR最小;6月30日后,均表现为FTD>FM>FR。3种模式下全生育期土壤呼吸速率平均值表现为FTD>FM>FR(P<0.05),平均排放速率分别为400.67 mg m-2h-1、234.38 mg m-2h-1和59.87 mg m-2h-1。在季后休闲期,3种种植模式的土壤呼吸速率较小,变化平稳,其中FTD的土壤呼吸速率高于FM和FR,FM和FR的土壤呼吸速率的部分时点为负值,表现为碳吸收(图2)。
2.2 不同种植模式下土壤呼吸组分特征
各处理土壤呼吸组分在作物季表现出不同的排放特征。总体来看,各处理土壤自养呼吸速率均表现为双峰变化趋势,但呼吸速率大小和峰值出现的时间有较大差异。FM自养呼吸速率峰值出现在6月30日和7月28日,FTM的峰值出现在7月28日和9月1日,FR模式的自养呼吸速率峰值出现在水稻拔节期(7月28日)和灌浆期(9月1日)。从整个生育期来看,FM和FTD的自养呼吸速率均显著高于FR,而FTD的自养呼吸速率在7月10日前低于FM,之后迅速上升并高于FM。整个作物季各处理平均自养呼吸速率表现为FTD>FM>FR(P<0.05),呼吸速率值分别为191.43 mg m-2h-1、153.08 mg m-2h-1和26.63 mg m-2h-1。季后休闲期,各处理自养呼吸速率均较低,其中FTM的自养呼吸速率表现为逐渐降低直至平稳状态,而FR和FM的自养呼吸速率一直维持很低水平,FTD、FM和FR处理在整个休闲期的平均呼吸速率分别为11.49 mg m-2h-1、-13.21 mg m-2h-1和2.41 mg m-2h-1(图3)。

图3 不同种植模式作物季及季后休闲期土壤自养呼吸与异养呼吸变化Fig.3 Changes of autotrophic and heterotrophic respiration in crop season and fallow period under different planting patterns
作物季各处理土壤异养呼吸速率呈现波动变化的特征,其中FTM波动较大,呈现出移栽期(6月10日)和灌浆期(8月21日)两个排放峰值,FM总体呈现单峰变化,峰值出现在7月28日,FR全生育期波动较小。从呼吸速率大小来看,FTM的异养呼吸速率显著高于FM模式和FR模式(P<0.05),FTM、FM和FR模式的平均呼吸速率分别为209.24 mg m-2h-1、81.30 mg m-2h-1和33.24 mg m-2h-1。季后休闲期,各处理异养呼吸速率均较低,且波动小,但FTM模式的异养呼吸速率较FM和FR稍高(图3)。
2.3 不同土地利用方式土壤呼吸累积排放量特征
各处理在作物季的土壤总呼吸累积排放量表现为FTD>FM>FR(P<0.05),其排放量分别为8948.69 kg/hm2、5910.46 kg/hm2和1117.64 kg/hm2;季后休闲期的土壤总呼吸累积排放量表现为FTD>FR>FM。其中,在作物生长季自养呼吸累积排放量表现为FTD>FM>FR,排放总量分别为4334.57 kg/hm2、3586.99 kg/hm2和353.92 kg/hm2,季后休闲期的自养呼吸累积排放量表现为FTD>FR>FM,其中FTD和FR表现为净排放,排放量分别为266.40 kg/hm2和19.52 kg/hm2,而FM表现为净吸收,其吸收量为319.91 kg/hm2;异养呼吸累积排放量在作物生长季的排放规律与总呼吸和自养呼吸一致,表现为FTD>FM>FR,其排放量分别为4614.12 kg/hm2、2323.46 kg/hm2和763.72 kg/hm2,季后休闲期异养呼吸排放量表现为FTD>FM>FR,其中FTD表现为净排放,排量为46.15 kg/hm2,FM和FR表现为净吸收,吸收量别为160.63 kg/hm2和482.27 kg/hm2。就作物生长季和休闲期整体来看,无论是自养呼吸、异养呼吸还是总呼吸均表现CO2的净排放,处理间表现为FTD>FM>FR,累积排放量分别为9261.23 kg/hm2、5429.92 kg/hm2和654.89 kg/hm2;其中,自养呼吸的累积排放量分别为4600.96 kg/hm2、3267.08 kg/hm2和373.44 kg/hm2;异养呼吸的累积排放量分别为4660.27 kg/hm2、2162.84 kg/hm2和281.45 kg/hm2(图4)。
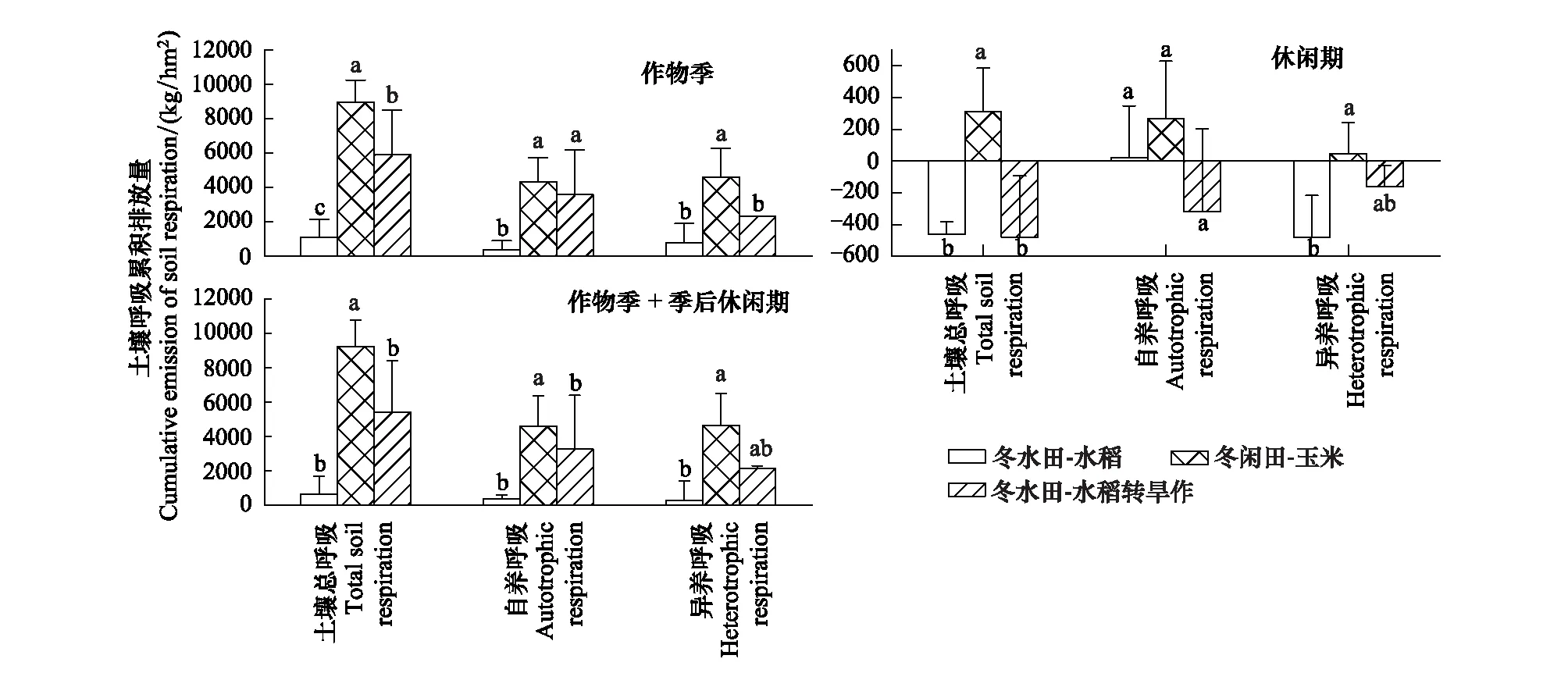
图4 不同种植模式作物季及季后休闲期土壤呼吸累积排放量特征Fig.4 Characteristics of soil respiration cumulative emission in crop season and fallow period under different planting patterns不同小写字母标注表示处理间在0.05水平上差异显著
根据土壤呼吸组分对总呼吸的贡献来分,作物生长季土壤自养呼吸对土壤总呼吸的贡献表现为FM>FTD>FR,其贡献量分别为60.69%、48.44%和31.67%;就休闲季来看,FTD表现为向大气释放CO2,而FR和FM表现为吸收CO2;就作物生长季和休闲期整体来看,自养呼吸对总呼吸的贡献表现为FM>FR>FTD,其贡献量分别为60.17%、57.02%和49.68%。因此,冬水田-水稻模式转旱作后显著提高了土壤呼吸及其自养和异养呼吸,增强了异养呼吸对土壤呼吸的贡献率。
2.4 不同土地利用方式土壤呼吸的温度敏感性
通过对土壤呼吸速率与土壤温度的回归分析发现,不同模式下土壤呼吸速率与0—5 cm和5—10 cm土壤温度均呈现极显著的正相关(P<0.01),表明土壤呼吸速率随着土壤温度的升高而增高。从不同土层来看,0—5 cm的Q10在3.7—27.6之间,5—10 cm的Q10在3.9—45.7之间,3种种植模式Q10均表现为0—5cm<5—10 cm,表明土壤呼吸速率与土壤温度的敏感性随深度增加而增大。从不同组分呼吸速率的Q10来看,表现为自养呼吸<总呼吸<异养呼吸,表明受温度影响较大是异养呼吸,受温度影响较小的是自养呼吸。从不同模式来看,在0—5 cm土层中,土壤总呼吸的Q10表现为FM>FR>FTD,其大小分别为14.9、8.9和8.4;异养呼吸的Q10表现为FR>FM>FTD,其大小为27.6、19.3和9.6;自养呼吸的Q10表现为FM>FR>FTD,其大小分别为7.7、4.5和3.7。在5—10cm土层中,土壤总呼吸的Q10表现为FM>FR>FTD,其大小分别为45.7、21.2和12.6;土壤异养呼吸的Q10表现为FR>FM>FTD,其大小为37.8、30.2和12.1;自养呼吸的Q10表现为FTD>FR>FM,其大小分别为5.8、5.1和3.9。这表明:冬水田-水稻模式转旱作后降低了土壤呼吸的自养呼吸、异养呼吸和总呼吸的温度敏感性(表3)。
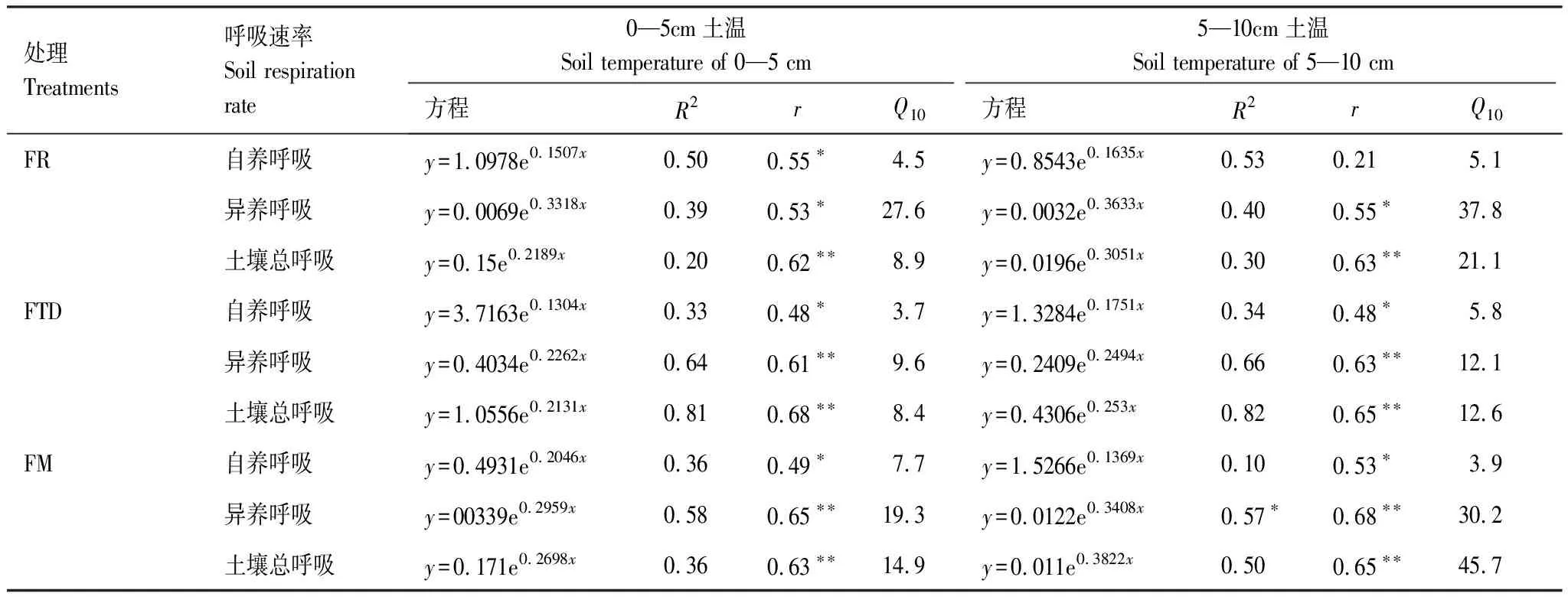
表3 不同种植模式作物季及季后休闲期土壤呼吸的温度敏感性
2.5 不同种植模式下农田系统碳平衡
不同种植模式下农田系统碳平衡测算结果如表4所示。结果表明,各模式的净初级生态系统生产力(NPP)表现为FTD 表4 不同种植模式作物季的农田系统碳平衡 稻田是受人类活动干扰频繁的一种农田生态系统,冬水田-水稻模式转旱作后,其作物类型及田间管理措施也会发生改变[23],冬水田由于长期淹水,土壤湿度较大,不利于CO2的排放,厌氧微生物较多。种植模式转变为旱作过程中,土壤水分降低,逐渐转变为氧化状态,厌氧微生物减少,好氧微生物增多,微生物活性增强,有利于有机质的氧化分解,促进CO2的排放[24—25]。此外常年旱作模式一直处于较为干旱的环境,由于其土壤长期与氧气接触,微生物活跃,且具有较高的温度使之具有较强的有机碳矿化能力和CO2排放能力[26]。 本研究结果表明,FTD>FM>FR(P<0.05)(图2),表现为冬水田-水稻模式的土壤呼吸小于旱作的土壤呼吸。其原因是由于在水稻季田间长期处于淹水状态,抑制了土壤呼吸作用,导致水稻田的土壤呼吸速率小于旱作环境[27—29]。将冬水田-水稻模式转为旱作后,富含碳氮的淹水土壤水分减少, 刺激好氧微生物生长,微生物分解的底物丰富,促进微生物活动并降解有机质,从而使CO2排放显著增加[24]。研究表明[30—31],土壤水分是影响土壤呼吸的重要因素,土壤含水量在一定范围时,随着土壤含水量的增加,土壤呼吸速率相应上升,当土壤含水量超出这个范围时,则会抑制土壤呼吸速率。在本试验的7月—8月间土壤呼吸出现了明显的波动现象,主要是由于该时期正值雨季。降雨后土壤含水量增高,透气性变差,土壤微生物的活动受到一定抑制,降低了旱作系统土壤呼吸速率。从作物季及季后休闲期的总呼吸速率来看,作物季的土壤呼吸速率显著高于季后休闲期(图2、3),这主要由于作物季土壤温度较高和根系生物量增加,土壤微生物活性增加,根源呼吸也十分活跃,进而提高了土壤总呼吸速率[3]。 土壤自养呼吸和异养呼吸是土壤呼吸的重要组成成分,自养呼吸对土壤呼吸的贡献变化很大,在整个生物群落,整个气候区以及不同年份之间在10%到90%之间,导致自养呼吸的时空变化很大[32—33]。研究表明[19,34]土壤自养呼吸的变化是由土壤温度、覆盖作物的光合能力和根系性状的共变异解释的,而土壤水分、温度和土壤碳对异养呼吸的变化有较强的解释力。本研究发现冬水田-水稻模式转旱作过程中,土壤自养呼吸和异养呼吸均显著增大,从而增强了土壤总呼吸。其原因是由于冬水田模式转变为旱作过程中,土壤湿度减小,温度升高,土壤微生物活性增强,增强了土壤碳的矿化能力,从而增强了异养呼吸[12]。此外,在模式转变过程中,覆盖作物类型也由原来的C3作物水稻变成了C4作物玉米,光合能力增强,温度升高增强了根系的活性,从而增强了自养呼吸[35]。 温度是土壤呼吸速率的主要影响因素之一。研究表明[36—37],土壤温度主要通过影响土壤微生物代谢和植物根系生长来调控土壤呼吸通量,土壤温度能解释土壤呼吸季节变化的14%—96%。土壤呼吸与土壤温度并不是简单的线性关系,单一因子模型无法全面的解释土壤呼吸[38]。大多数学者用采用指数模型,用Q10表示土壤温度与土壤呼吸的关系,Q10表示温度每增加10℃土壤呼吸增加的倍数。本研究表明,土壤温度与土壤呼吸及其自养和异养呼吸速率呈显著正相关关系,3种种植模式的Q10变化范围为3.7—45.7,并且自养呼吸的温度敏感性介于异养呼吸和土壤总呼吸之间,这表明土壤异养呼吸随土壤温度变化的幅度更大,温度主要通过影响异养呼吸的变化来调节土壤呼吸的变化。前人研究表明,Q10值变化范围一般在1.3—3.3之间。本实验结果偏大,并且随着土层的加深,Q10增大。这与赵爽凯等[39]的研究一致,原因可能是在湿地生态系统中,表层的土壤温度高于更深土层的温度,更深层的温度变化幅度小,很微小的变化就能引起土壤呼吸的剧烈变化。 本研究还发现,异养呼吸比自养呼吸具有更高的温度敏感性(表3),表明有机质分解比根源呼吸对温度的变化更为敏感。前人研究[5]表明,温度升高对自养呼吸没有显著影响,但是能显著影响异养呼吸,这与本试验的研究结果一致。这是由于温度升高能够提高土壤微生物和酶的活性,刺激土壤异养呼吸,从而促进土壤CO2的排放[40]。关于异养呼吸和自养呼吸温度敏感性差异,前人的研究结果存在较大分歧[41]。 农田土壤固碳在温室气体减排中占有重要地位。研究表明[42],农田生态系统可以在较短的时间尺度上对碳库起调节作用。据估算,中国农田土壤固碳潜力占土壤总固碳潜力的20%[43],张宇等[8]研究认为不同耕作下小麦田土壤均表现为大气CO2排放的汇。本试验结果表明,3种种植模式的生态系统在作物季均表现为碳汇(表4),说明作物光合作用固定CO2的量高于土壤异养呼吸释放的CO2量。然而,冬水田-水稻模式转旱作过程中的碳汇强度显著低于冬水田-水稻模式和冬闲田—玉米模式,主要是种植模式改变导致土壤碳排放量增大,抵消了部分系统净初级生态系统生产固定的碳,从而导致净生态系统生产力减小,弱化了系统的碳汇功能。因此,在冬水田种植系统管理过程中,应构建稳定的农田生态系统,或者在不同种植模式的转变过程中,配置固碳效率高的作物,结合高效低排栽培管理技术以实现碳汇强度的增加。 冬水田-水稻是川中丘陵区传统的稻田种植模式,通过与冬水田-水稻模式和冬闲田—玉米模式对比,对冬水田-水稻转旱作的土壤呼吸组分特征和农田系统碳平衡进行研究,结果表明,(1)冬水田-水稻转旱作后的土壤呼吸速率(400.67 mg m-2h-1)最大,其累积排放量(9261.23 kg/hm2)也显著高于冬水田-水稻模式和冬闲田—玉米模式(P<0.05)。冬水田-水稻转旱作后显著提高了土壤自养呼吸和异养呼吸,从而显著提高了土壤总呼吸速率(P<0.05)。(2)3种种植模式的土壤温度与土壤呼吸速率均呈现极显著的正相关关系,并且对温度变化较为敏感,其Q10的变化范围为3.7—45.7,其中FM对温度变化最为敏感,FR次之,FTD最小,说明冬水田模式转旱作后降低了其温度敏感性。(3)3种种植模式的净生态系统生产力均为正值,表现为大气CO2的碳汇,但冬水田-水稻转旱作后,由于异养呼吸显著增加导致净生态系统生产力显著小于冬水田-水稻和冬闲田—玉米模式,呈现出较弱的碳汇。因此,在川中丘陵区冬水田利用管理中,应减少冬水田种植系统盲目转换,要通过碳高效品种及高效低排栽培措施,并构建稳定的农田生态系统基础上实现冬水田利用方式转换,以达到固碳增汇的目的。
3 讨论
3.1 冬水田-水稻模式转旱作对土壤呼吸及其组分的影响
3.2 土壤温度对土壤呼吸及组分的影响
3.3 冬水田-水稻种植模式转旱作对农田碳汇强度的影响
4 结论
