树种多样性对土壤微生物群落结构和元素生物地球化学循环的影响研究进展
2022-08-31施秀珍王建青黄志群贺纪正
施秀珍, 王建青,黄志群,*,贺纪正
1 福建师范大学地理科学学院, 福州 350007 2 福建师范大学湿润亚热带山地生态国家重点实验室培育基地, 福州 350007
森林生态系统是全球生物多样性的重要聚集地[1]。生物多样性在维持生态系统功能和稳定性等方面发挥重要作用[2]。森林生态系统包括地上和地下两大组成部分,二者相互作用,共同发挥森林生态系统功能[3]。关于树种多样性对森林生态系统功能影响的研究主要集中在地上部分[4—5],而较少关注到地下土壤微生物。近年来森林生物多样性研究取得系列重大进展,证实树种多样性增加能显著提高森林生态系统生产力[6—8]。生态位互补效应(Niche complementarity effects)是解释树种多样性促进森林生产力提高的一个最常用的生态机制,即当生态系统的生态位被不同树种占用时,生态系统中的资源(如空间、光照、养分等)能够被更有效地获取和利用,从而促进森林生产力的提高[9—10]。但研究发现,去除土壤微生物后,植物多样性对生产力的促进作用显著下降[11],这预示土壤微生物在调控树种多样性与生态系统功能中发挥关键性作用。
森林土壤微生物群落结构和功能受到生物与非生物因素的共同影响。目前,有关非生物因素,如火烧、氮沉降等对森林土壤微生物的影响已开展较多的研究[12—14]。然而,森林树种多样性作为森林生态系统最重要的生物因子,其对地下土壤微生物及其介导的元素生物地球化学循环的影响研究较少。近些年来,随着微生物免培养技术即分子生物学技术的迅速发展和广泛应用,极大地推动了作为“黑箱”的森林土壤微生物的发展[15],森林树种多样性对地下土壤微生物及生态功能的影响研究备受关注。本文首先介绍树种多样性对土壤主要微生物类群,细菌和真菌群落结构和多样性的影响;在此基础上,深入分析由功能微生物群介导的碳、氮和磷元素生物地球化学循环对树种多样性的响应机制;最后提出需要进一步深入研究的方向,以期为阐明森林生态系统地上地下联系及其对生态系统功能的影响提供新思路。
1 树种多样性对土壤细菌的影响
土壤微生物包括细菌、真菌、病毒、原生动物和显微藻类等多个类群。鉴于细菌和真菌是土壤微生物中数量及生物量最高的类群,且目前关于树种多样性与土壤微生物关系的研究主要涉及土壤细菌和真菌。因此,本文着重综述树种多样性对土壤细菌和真菌多样性和群落结构的影响。目前研究表明,树种多样性有利于促进土壤细菌生物量和多样性增加。树种多样性水平不同的森林生态系统在凋落物数量、质量和根系分泌物输入等方面存在较大差异,成为影响土壤细菌多样性和群落结构的重要因素[16—17]。Zhang等[18]通过对亚热带森林五个单一树种纯林和三个混交林(两个树种)土壤中细菌系统多样性的分析指出,混交林显著提高了土壤细菌系统发育多样性和细菌群落稳定性(表1)。
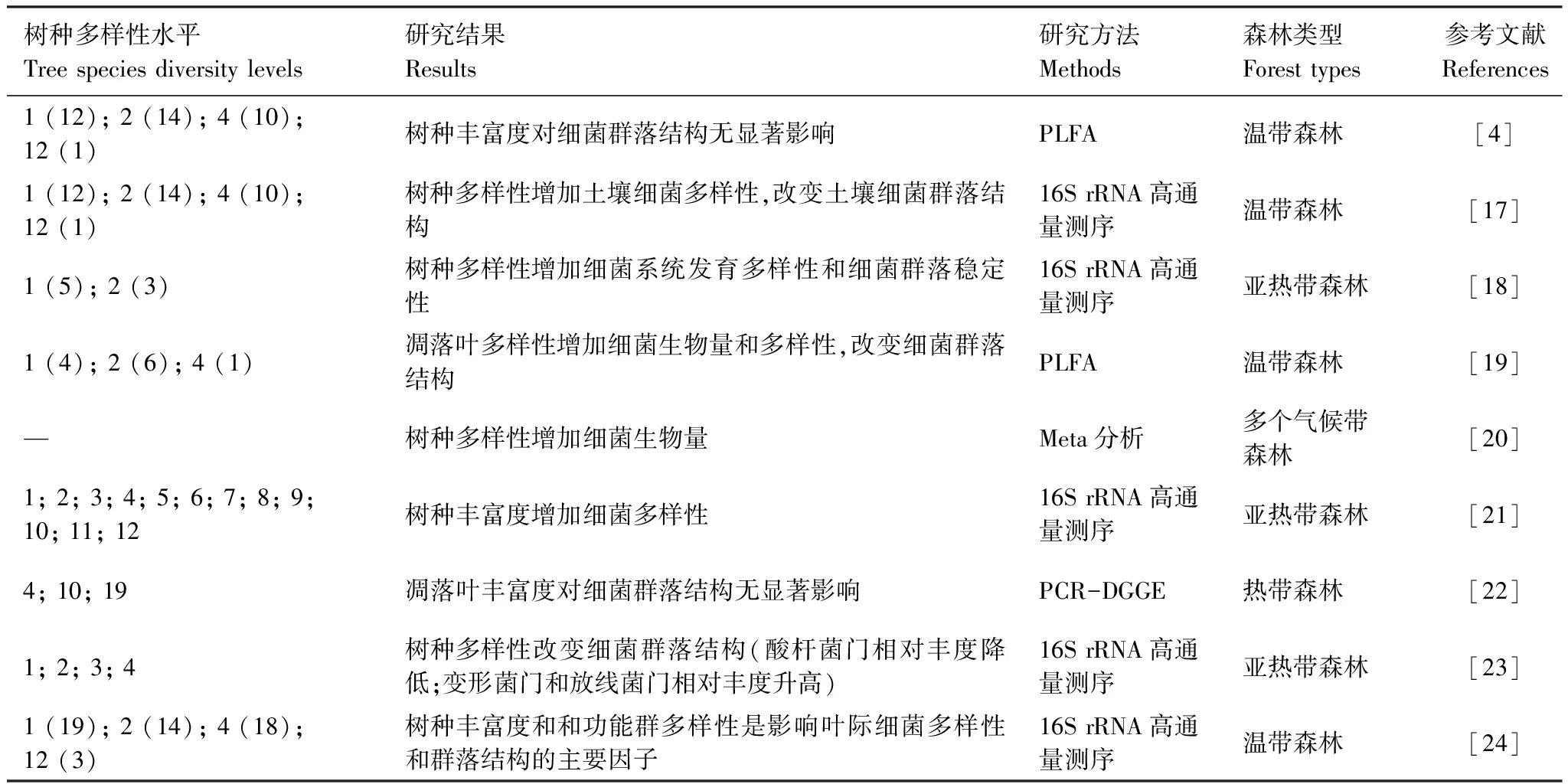
表1 树种多样性对细菌多样性及群落结构的影响
括号外数字表示树种多样性水平,括号内数字为相同多样性水平时树种组合数,“—”表示无相关数据;PLFA: 磷脂脂肪酸Phospholipid Fatty acid;DGGE: 变性梯度凝胶电泳Denaturing gradient gel electrophoresis
细菌系统发育多样性(Phylogenetic diversity)旨在描述一个群落中不同种细菌进化和系统发育距离,可用于评估树种多样性对生态系统功能和稳定性的影响[25]。系统发育多样性增加意味着细菌群落具有更多的生态位分化和资源互补利用能力,对外界环境变化具备更高的抵御能力。树种多样性对土壤细菌系统发育多样性的促进作用源于具有高度养分异质性的凋落物混合物,其能为土壤细菌提供更加多样化的生态位[19]。Wardle[26]指出混交林中不同树种叶片功能性状(比叶片面积、叶干物质含量等)多样化水平越高,其凋落物混合效应越强。鉴于树木地上叶片和地下根系的功能特性通常具有协调一致性,叶片功能性状的多样性通常代表根系结构和分泌物的多样性[27],后者可为根际微生物生态位分化提供空间和物质基础。研究者通过分析土壤微生物磷脂脂肪酸(PLFA)发现,树种(凋落物)多样性增加能促进土壤细菌生物量的增加[19—20]。近期,Rivest等[17]和Wu等[21]对细菌16S核糖体RNA高通量测序分析表明,温带和亚热带森林树种丰富度增加均显著提高土壤细菌香农指数多样性。
细菌群落结构是表征细菌群落动态及生态功能的另一个重要指标。目前关于土壤细菌群落结构对树种多样性的响应仍没有定论。一部分研究显示树种多样性对土壤细菌群落结构没有显著影响。如吴艺雪[22]研究结果表明,热带森林凋落叶多样性对土壤细菌群落结构(PCR- DGGE)无显著影响。在加拿大的温带树种多样性同质园试验中,Khlifa 等[4]发现树种丰富度对土壤细菌群落结构(PLFA)没有显著差异。但是更多的试验研究发现树种多样性对细菌群落结构具有显著影响。陈琴[23]通对亚热带森林研究发现,随树种丰富度增加,酸杆菌门Acidobacteria相对丰度降低;而变形菌门Proteobacteria和放线菌门Actinobacteria相对丰度增加。这可能归因于树种丰富度增加时,提高了凋落物的输入量,有利于富营养型细菌,如变形菌门和放线菌门成为优势菌群,而不利于寡营养型细菌如酸杆菌门的生长[28]。通过对已建立七年的温带森林树种多样性样地研究,Rivest 等[17]发现树种多样性显著影响土壤细菌的群落结构。研究还显示树种丰富度和功能群多样性是影响叶际细菌多样性和群落结构的重要因子[24]。不一致的研究结果除了归因于树种、林龄及分析微生物群落结构所采用方法的差异,还需考虑到土壤微生物本身对树种多样性响应的时滞性(Time lag)[17,29]。例如,Eisenhauer等[2]设计了一个连续六年的野外试验研究土壤微生物对植物多样性的响应,结果发现植物多样性样地建立初期,土壤微生物量和呼吸速率在不同植物多样性水平间无显著差异,但在多样性样地建立四年后随植物多样性增加。这种时滞性是由于大多数土壤微生物为异养型,需要依靠植物源养分的输入而获取生长所需要的养分和能量[3]。
2 树种多样性对土壤真菌的影响
真菌是土壤中一类重要的真核微生物。自然界中绝大多数植物能够与菌根真菌(Mycorrhiza)形成共生关系,包括内生菌根和外生菌根真菌两种类型[30]。其中,丛枝菌根是最为常见的内生菌根真菌,能够与80%的陆地植物形成共生关系[31]。研究表明,土壤菌根真菌是调控和维持树种多样性的重要生物因子[30, 32—33]。这主要由于当更多的树种与不同类型/物种的菌根真菌共生时,有利于增加资源利用互补性,减少树种之间的竞争。例如,外生菌根能产生胞外酶来降解土壤中复杂的有机氮,从而为植物提供氨基酸作为氮源;丛枝菌根主要协助植物获取土壤无机氮[34]。Jansa 等[35]报道丛枝菌根的不同物种对无机磷具有不同的亲和力及获取策略,较高的丛枝菌根丰富度可以促进不同树种对磷的吸收。菌根真菌多样性增加有助于提高生态系统生产力[30,36]。这种促进效应,一方面归因于菌根真菌生命活动中产生的代谢产物,如有机酸能促进土壤中矿质元素的释放,进而增强植物对矿质养分的吸收[37];另一方面是菌根真菌能增强植物对病原性真菌的抵抗能力[38]。
不同于菌根真菌,病原性真菌则可引发寄主树木组织坏死、幼苗死亡等[32]。研究发现树种多样性增加可以显著降低病原性真菌的数量及丰富度,对森林生态系统功能产生积极影响[39—40]。主要由于土壤病原性真菌大多为专性寄生,传播孢子能力有限,对同种寄生植物具有距离和密度依赖性[33]。增加树种多样性能够降低寄主植物的密度(稀释作用),而且局部微环境可能发生改变,均不利于病原性真菌孢子的传播和萌发[39]。
鉴于树种与真菌广泛存在的共生或寄生关系,树种与真菌之间的相互作用能促进二者协同进化。因此,不同的树种在其系统发育过程中与土壤真菌群落建立了密切联系[41]。Wang等[33]研究发现亚热带森林树种系统发育特性对土壤菌根真菌和病原性真菌的丰富度和群落结构变化的解释度高达77%,而环境和空间因素的解释度均低于4%。Nguyen等[42]通过研究也发现,温带森林树种系统发育多样性与外生菌根真菌物种丰富度呈显著正相关,树种组成对外生菌根真菌群落结构具有显著影响。Schroeder等[32]也提出,热带森林树种系统发育多样性可用于预测病原性真菌和菌根真菌的多样性及群落结构(表2)。
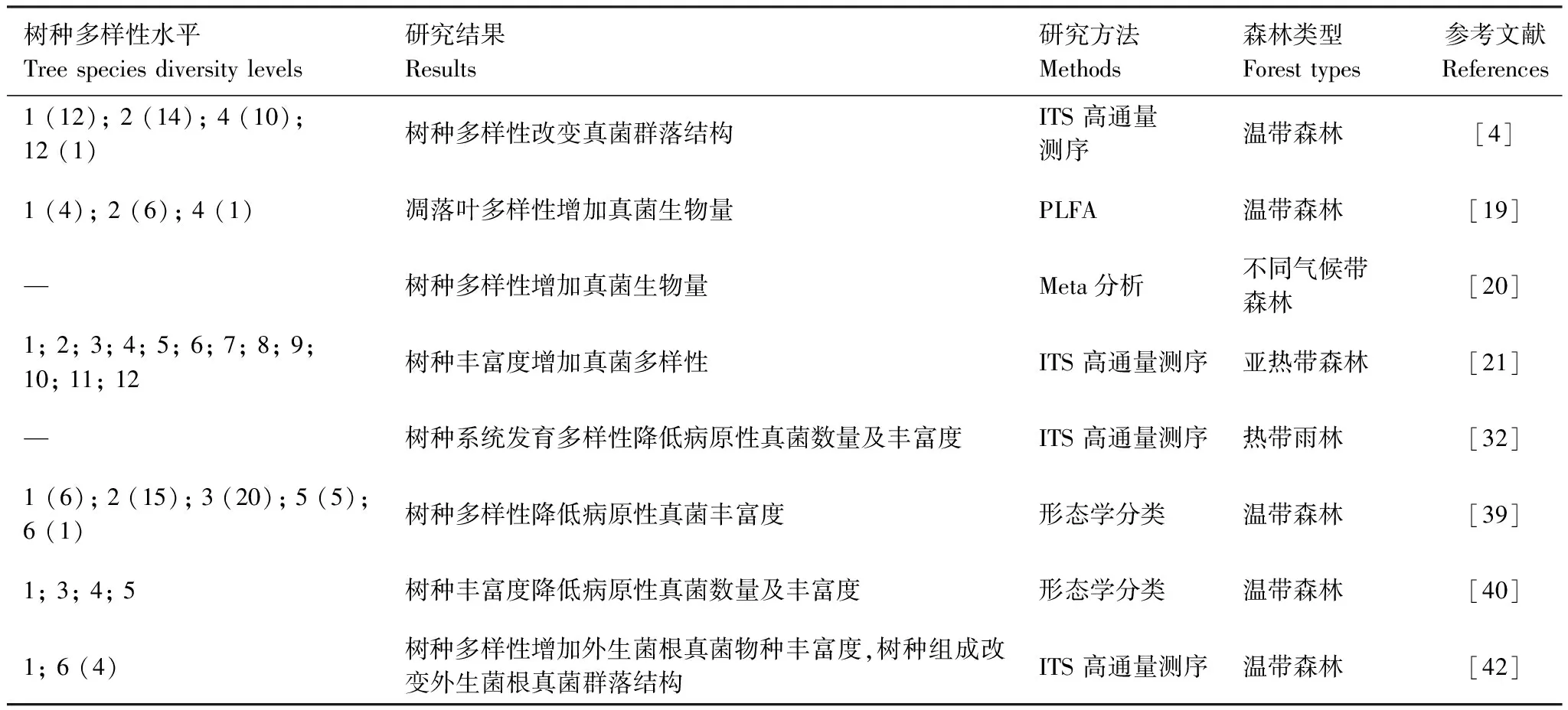
表2 树种多样性对真菌多样性及群落结构的影响
3 树种多样性对碳、氮和磷生物地球化学循环的影响
若将上述土壤微生物的多样性和群落结构作为对微生物群落动态的‘定量描述’,那么接下来元素生物地球化学循环则是对微生物群落的‘定性分析’。其中,土壤功能微生物群(Functional microbial guilds)是推动碳素、氮素和磷素生物地球化学循环的引擎[15](图1)。伴随着越来越多的土壤功能微生物被用作物质循环转化速率的生物指标(Bio-indicator),揭示土壤功能微生物对树种多样性的响应,有助于深入理解森林生态系统地上地下相互作用及其对生态系统物质循环的影响。
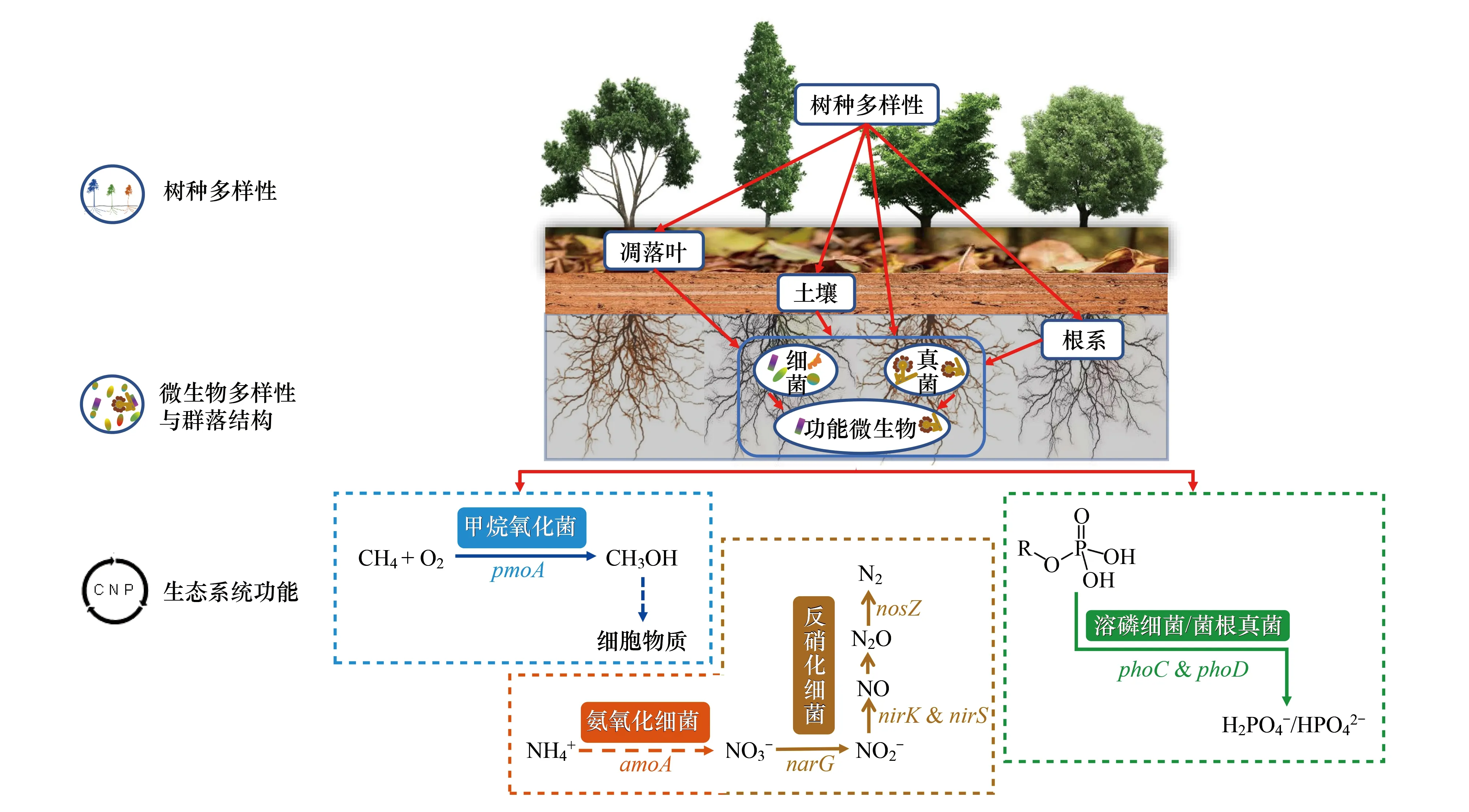
图1 树种多样性对碳、氮和磷生物地球化学循环的影响Fig.1 Effects of tree species diversity on the biogeochemical cycling of carbon, nitrogen and phosphorus
3.1 树种多样性对碳素生物地球化学循环的影响
森林生态系统植被和土壤中碳储量占陆地有机碳总量的39%,将森林作为全球碳平衡管理的杠杆潜力巨大[43]。了解树种多样性对森林有机碳储存的影响机制有利于准确评估森林生态系统应对全球气候变化的潜力[44]。当树种多样性增加时,树木以凋落物、根系分泌物和细根周转的形式向土壤中输送的有机质增加[45]。土壤微生物在调控新输入的植物源有机质向土壤有机碳转化过程中发挥重要作用。不同气候带森林生态系统研究均表明树种多样性增加能促进土壤有机碳储量积累[8, 10,21,46]。主要由于植物多样性增加时,植物性碳源输入(如凋落物和根系生物量)增加,能够为土壤中以有机碳作为碳源的大量异养型微生物提供营养和能量,促进其生长繁殖和生物量的积累[47]。当微生物死亡后,微生物残体转化为新增的土壤有机碳[48]。据Liang等[49]统计,经微生物合成代谢所固定的有机碳占土壤有机碳总量一半以上。其次,树种多样性增加有利于维持适宜的土壤含水量,从而减轻干旱或全球气候变暖对土壤微生物生长的不利影响[50]。值得注意的是微生物生物量增加的同时也将加强微生物呼吸作用(土壤异养呼吸)。近期,通过对全球包括森林在内的多个陆地生态系统类型植物多样性的整合分析指出,土壤异养呼吸作用随植物多样性增加而加强[20],但呼吸作用引起的碳释放远低于由微生物合成代谢而固定的碳[20,48]。因此,土壤微生物生物量增加是树种多样性提高森林生态系统土壤碳储量的一个重要途径。
森林是陆地生态系统最大的甲烷(CH4)汇,维持树种多样性有利于增强森林生态系统CH4氧化能力[51]。相比于在分类学和系统学上分布广泛的介导呼吸作用和分解作用的微生物类群, 甲烷氧化菌是一类专性好氧的自养型微生物,能通过氧化大气中甲烷而获得生命代谢能量,并将甲烷同化为自身生物量(图1)。因此,甲烷氧化菌不仅是一类重要的生物碳汇,而且对减少CH4排放发挥关键作用,是森林生态系统碳循环研究中的重要功能微生物[5]。东南亚热带地区近几十年来大规模的天然林向橡胶Heveabrasiliensis纯林的转变严重降低了树种多样性,近期研究报道天然林对CH4的氧化能力是橡胶纯林的2.4倍,林龄较大的橡胶纯林在雨季甚至从CH4的汇变为源[52]。相似的研究结果发现,天然林转变为牧场、咖啡或棕榈种植园后,土壤对CH4的氧化能力均显示出下降趋势[53—55]。关于树种多样性增加时,森林CH4氧化能力提高存在以下几种机制:1)维持适宜的土壤含水量,热带森林地区水热条件充足,单一树种纯林导致土壤含水量偏高,土壤厌氧环境促进产甲烷细菌的生长,而对专性好氧的甲烷氧化菌产生抑制作用,因此森林土壤CH4的排放增加[52]。Krämer 等[56]对德国中部12个温带落叶林树种多样性梯度样地的土壤水分动态研究发现,树种多样性与表层土壤水分在夏季干燥期呈显著正相关,而在非干燥期不同树种多样性水平样地土壤水分动态格局相似。因此,当树种多样性水平增加时,不同树种水分利用策略的差异可能有助于促进对土壤水分的充分利用,从而维持土壤适宜的含水量。即当环境中水分充足时,树种多样性水平高的森林系统能降低对深层土壤水分的提取,而当生境中缺水时,则以更高的速率提取深层土壤水分,且本身较高的叶面积指数也有利于保持较高的表层土壤水分[57]。这种水力策略可能是不同树种多样性水平下森林土壤CH4氧化能力差异的驱动性因素。2)土壤细菌多样性增加促进甲烷氧化菌对甲烷的氧化。据Schnyder 等[5]研究报道,土壤微生物多样性增加对甲烷氧化菌的功能基因pmoA(编码甲烷单加氧酶)丰度有促进效应,进而增强对甲烷的氧化能力。因此,树种多样性可能通过增加土壤细菌多样性[6, 17,21],间接增强土壤甲烷氧化菌对甲烷的氧化潜力。3)一些研究表明树种多样性增加能提高土壤中氮素的有效性[58—60],因此间接促进土壤对CH4的氧化。氮素可利用性增加促进甲烷氧化菌对甲烷的氧化。早期研究表明当氮素成为限制甲烷氧化菌生长的营养元素时,环境中氮素的提高能够促进甲烷氧化菌的生长,从而促进对CH4的氧化[61]。Yan等[62]研究也指出,在温带森林中氮添加处理能显著增加土壤对CH4的氧化能力。
3.2 树种多样性对氮素生物地球化学循环的影响
关于森林生态系统树种对土壤氮素生物地球化学循环的影响,多样性假说(Diversity hypothesis)提出当森林生态系统中树种丰富度增加时,不同树种凋落物形状、大小、结构和颜色的差异,有助于提高微环境的丰富度与斑块性,为土壤微生物和动物提供更多的生境;而且不同树种凋落物的分解速率不同,能提供持续的氮素来源,从而加快土壤氮循环的转化速率[63]。例如,Bengtson等[64]研究指出,随温带森林混交林中树种丰富度的增加,土壤有机氮净矿化速率和净硝化速率均提高。Reverchon 等[58]报道与单一树种的纯林相比,所罗门群岛的热带森林中混合种植柚木Tectonagrandis和白饭树Flueggeaflexuosa混交林显著提高土壤的速效氮含量,混合林土壤反硝化细菌功能基因narG、nirS和nosZ(依次编码硝态氮还原酶、亚硝态氮还原酶和氧化亚氮还原酶)的丰度显著升高。在刚果和巴西的热带森林中,Tchichelle等[59]和Voigtlaender等[60]发现相比于纯林,尾叶桉Eucalypturophylla与马占相思Acaciamangium等比例混交林显著提高土壤速效氮含量和树木生物量的积累。Chang等[65]通过研究亚热带人工湿地也发现,随植物物种丰富度的增加,土壤净硝化速率、硝酸还原酶活性和土壤氨氧化菌的丰度均显著增加。
也有不同于多样性假说结果的研究报道。例如,吴艺雪[22]通过分析热带森林凋落叶丰富度对土壤有机氮矿化的影响指出,当凋落叶丰富度较高时(十种以上),土壤净氮矿化速率显著增加,而当凋落叶丰富度较低时(低于四种),土壤净氮矿化速率急剧降低。Isobe 等[66]研究我国亚热带针叶纯林和阔叶林发现,其土壤净硝化速率分别是混交林的10和2.4倍,氨氧化古菌的功能基因amoA(编码氨单加氧酶)丰度分别是混交林的3.8和2.9倍。针对上述差异性的结果,研究者指出相比于树种丰富度,森林生态系统中优势树种的性状对土壤氮循环可能发挥决定性的作用,即质量比假说(Mass ratio hypothesis)[67]。例如,热带雨林中以美洲椴TiliaAmericana和美洲糖槭Acersaccharum为优势树种时,其土壤有机氮净矿化速率和净硝化速率均显著高于以东方铁杉TsugaCanadensis为优势树种,主要由于美洲椴和美洲糖槭的凋落物木质素与氮素的比值及碳氮比值均低于东方铁杉,所以凋落物质量高,加快了土壤有机氮矿化和硝化作用。通过遥感图像光谱学研究报道,相比于热带雨林冠层含氮量低的树种组合,冠层含氮量高的树种组合显著提高土壤净硝化速率及土壤硝态氮含量,氨氧化古细菌的amoA基因丰度显著提高,反硝化细菌的nirS、nirK和nosZ基因丰度没有差异[68]。近期,Laffite 等[69]发现高加索冷杉Abiesnordmanniana、欧洲云杉Piceaabies和以欧洲山毛榉Fagussylvatica为优势树种的温带天然林能通过凋落物和根系分泌物,抑制硝化杆菌属的生长,进而降低土壤硝化速率。综上所述,针对树种多样性对氮素生物地球化学循环的影响需要考虑多样性假说和质量比假说的相对贡献。
3.3 树种多样性对磷素生物地球化学循环的影响
磷是生物遗传信息载体DNA、生物膜以及生物能量转换和贮存物质组成中不可缺少的大量元素, 因此磷是生物的核心元素[21,70]。不同于碳、氮元素,磷元素及其化合物没有气态形式,土壤中有效磷主要来源于成土母质中不溶性磷化物,如磷酸钙和磷灰石的风化,因此磷被广泛认为是天然生态系统,如森林生态系统中限制植物生长的“瓶颈”元素[46],近些年来持续增加的氮沉降加剧了森林生态系统的磷限制[57]。因此,有研究提出土壤速效磷的增加是植物多样性与生态系统功能正相关的潜在作用机制[21]。

4 展望
近期欧洲大数据分析发现,森林生态系统树种多样性对于维持鸟类、昆虫和林下植被等生物多样性具有重大生态效益[1]。树种多样性在维持森林生态系统功能稳定性及增强对全球气候变化应对能力等方面发挥重要功能[25]。建议今后从以下几方面加强森林树种多样性对土壤微生物影响的研究。
(1)加强树种多样性对多个营养级之间相互作用(营养网络)的研究。目前关于树种多样性与生态系统功能关系的研究多集中于同一个营养级水平,但生态系统的功能在很大程度上取决于多个营养级之间复杂的相互作用。例如,当植物多样性增加时,土壤中动物分解者,如蚯蚓可以通过加快地表凋落物分解(增加营养有效性)和增加地下栖息地空间,为土壤微生物提供更多异质性的生境[35]。因此,今后的研究需要考虑多个营养级生物的相互作用,如土壤微生物和土壤动物互作对森林树种多样性的响应及对生态系统功能的影响。
(2)关注树种多样性对生态系统多功能(Ecosystem multifunctionality)的影响。目前关于树种多样性的研究多注重于对单一生态功能的影响,严重制约对树种多样性与生态系统功能关系的客观评价。例如,树种多样性能增强甲烷氧化菌对土壤CH4的氧化能力,但同时也可能导致N2O排放增加,因此需要综合评估树种多样性对温室气体排放的影响。由于生态系统多功能可以同时考虑不同生态系统功能之间的权衡(Trade-offs),其提出被认为是生态学研究的一个重要进展[78]。今后应加强树种多样性对生态系统多功能和服务能力的研究,如从不同元素生物地球化学循环耦合角度探讨树种多样性对森立生态系统多功能的影响。
(3)加强学科交叉,引入种群动态模型和气候模型等模型预测方法,研究树种多样性对全球气候变化的应对机制。目前,全球面临的气候变暖、淡水资源短缺、环境污染和极端气候频发等问题日益突出,严重威胁着人类的生存和发展。Hisano 等[44]提出“生物多样性—减缓概念”(Biodiversity-Mitigation concept),即生物多样性可能减缓全球环境变化对生态系统造成的负面影响,值得借鉴。近期,Fichtner等[79]研究亚热带森林树种多样性对干旱的响应指出,树种多样性能减缓干旱对幼龄期树木生长的不利影响。今后应在此基础上,加强学科交叉和新兴技术的应用,引入微生物种群动态模型预测的方法,进一步深入地研究树种多样性适应和缓解全球环境变化的生物机制。
