天台鹅耳枥种群数量特征及其与其他树种的关联性
2022-08-18许元科赵昌高廖瑜俊吴传波吴初平王志高
许元科,赵昌高,廖瑜俊,林 坚,吴传波,吴初平,王志高
(1.景宁畲族自治县生态林业发展中心,浙江 景宁 323500;2.浙江省林业科学研究院,浙江 杭州 310023)
物种在群落内的多度、个体大小、竞争关系等特征反映了其在群落中的地位,是群落演替过程中对生境长期适应的结果。研究濒危物种在群落内的地位是判断物种能否长期在群落中生存的依据,也是揭示物种濒危机制的主要途径[1-4],为针对性开展科学有效的保护措施提供基础[5]。天台鹅耳枥Carpinus tientaiensis分布区域十分狭窄,为国家Ⅱ级重点保护野生植物,被《中国生物多样性红色名录 高等植物卷》列为极危等级(CR)。前期仅在浙江省天台县天台山和磐安县大盘山共发现了24株成树,生境地群落环境复杂,物种多样性较高,天台鹅耳枥与柳杉Cryptomeria fortuneivar.sinensis、黄山松Pinus taiwannensis和云锦杜鹃Rhododendron fortunei等共同组成了群落乔木层的优势树种[6-7]。天台鹅耳枥光照适应能力较强,其幼苗可以适应较强的光合有效辐射[8-10],海拔、坡度和干扰是影响群落结构和空间分布的主导环境因子[7]。天台鹅耳枥为雌雄异花植物,雄花始花期早于雌花,柱头可授期与雄花散粉的重叠时间约 15 d,授粉成功率受空气湿度和阴雨天气的影响较大;种子空壳率超过90%,自然更新困难[11]。播种育苗、扦插育苗及组织培养等人工繁育技术[12-13]对天台鹅耳枥种群更新效果不大,该物种濒危的状况未得到明显的改善。2019年5月,浙江省景宁畲族自治县上山头发现了新的天台鹅耳枥分布点,这是目前发现的面积最大的野生种群。本研究在对景宁上山头天台鹅耳枥群落结构进行充分调查的基础上,分析天台鹅耳枥种群数量特征及其与群落内其他树种的关系,以期从群落生态学的角度探讨该种群的濒危机制,为该物种的保护提供参考。
1 研究地区与研究方法
1.1 研究区概况
景宁上山头位于浙江省景宁畲族自治县县城南21 km,处于大漈乡、景南乡两地交界处,属洞宫山脉右支主峰,最高海拔为1 689.1 m,为景宁县境内最高峰。年平均气温为11.8 ℃,年平均降水量为1 918 mm,年日照时数为 1 717.6 h,无霜期为 196 d,森林覆盖率为 96.5%。上山头分布有近 70 hm2的云锦杜鹃林,胸径10~20 cm,树高2~5 m,树龄均在50 a以上。2019年5月,在云锦杜鹃林内共发现天台鹅耳枥354株,分布海拔为1 500~1 600 m,分布面积约1 hm2,是目前发现面积最大、分布海拔最高的天台鹅耳枥野生居群。
1.2 样地设置与调查
2019年9月,根据天台鹅耳枥实际分布情况,在上山头分布区设置140 m×60 m(样地1)和50 m×50 m(样地2)等2个样地,调查时划分成109个10 m×10 m样方。记录每个样方的木本植物(胸径≥1 cm)种类、高度、胸径。样地1和样地2中心海拔分别为1 531和1 580 m,坡度均为25°,东北坡向、上坡。土壤类型为黄壤,厚度为30~50 cm。
1.3 数据分析
以2 cm为径级,统计每个径级的个体数目。以10 m×10 m样方尺度进行重要值和种间联结值的计算。重要值Iv=(Ra+Rp+Rf)/3。其中Ra为相对多度,Ra=(某一植物种类株数/所有植物种类株数之和)×100%;Rp为相对显著度,Rp=(某一树种的胸高断面积/所有树种的胸高断面积之和)×100%;Rf为相对频度,Rf=(某一树种的频度/所有树种的频度之和)×100%。
采用层次聚类法根据种群重要值进行聚类分析,根据距离系数划分群组。
采用Godron稳定性指数评价群落稳定性。建立种百分数与累积相对频度回归方程,求回归方程与横坐标(0,100)和纵坐标(100,0)连线的交点,交点越接近种百分数与累积相对频度比值(20/80),群落就越稳定[14]。累积相对频度为乔木层出现的树种相对频度由大至小累积序列,种百分数为树种总数的倒数累积序列。建立种百分数与累积相对频度的幂函数、指数函数、二项式和Gompertz回归方程,根据R2大小选择最优模型。
采用非连续性数据的公式计算种间联结值(χ2)。公式为:
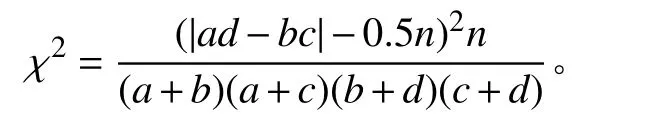
其中:n为样方总数;a为种A和种B都出现的样地数目,b和c分别为仅有种A或种B出现的样地数目,d为种A和种B均未出现样地数目。χ2<3.841,表示种A和种B种间联结独立,两者关联性不显著(P>0.05);3.841≤χ2≤6.635,表示种A和种B种间联结显著,两者有显著关联性(P<0.05);χ2>6.635,表示种A和种B存在极显著关联性(P<0.01)。联结系数(AC)计算公式为:
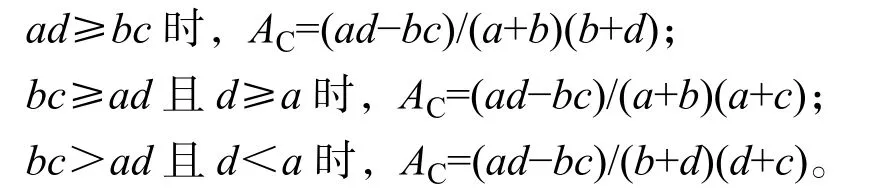
其中:AC为[-1,1],AC越接近1,表明种A与种B间的正联结性越强;AC越接近-1,表明种A与种B间的负联结性越强;AC越接近0,表明种A与种B间的联结性越弱;AC=0,表明种A与种B间完全独立,没有关联性。
共同出现百分率(PC)计算公式为:PC=a/(a+b+c)。其中:PC为[0,1],PC越接近1,表明种A与种B的正联结关系越强;PC越接近0,表明种A与种B的正联结关系越弱。
2 结果与分析
2.1 天台鹅耳枥群落木本植物数量特征
天台鹅耳枥所在群落中,云锦杜鹃重要值最大,为 26.84%,其次为白檀Symplocos paniculata(12.98%),天台鹅耳枥重要值排在第3位,为12.73% (表1)。群落中胸径1 cm以上的木本植物有51种3 348株,隶属于19科,31属,裸子植物仅松科Pinaceae松属Pinus黄山松1种。群落中个体数目排在前3位的树种分别是云锦杜鹃、白檀和天台鹅耳枥,分别占个体总数的37.19%、13.39%和10.60%。个体数目最多的前9个树种,占调查到树种个体总数的80%以上,个体数较少的21个树种仅占个体总数的1%,有12个树种仅统计到1株(图1)。群落中胸高断面积最大的12个树种的累积百分比占整个群落的90%以上,胸高断面积最大的是云锦杜鹃,占28.12%,其次为天台鹅耳枥,占17.79%,黄山松占14.44%,胸高断面积较小的21个树种,累积百分比仅占不到1%(图1)。利用重要值进行聚类分析发现:天台鹅耳枥为群落的主要优势树种;51个树种根据距离系数划分为3个群组,其中群组Ⅰ仅云锦杜鹃1个物种;群组Ⅱ包括天台鹅耳枥、白檀、黄山松和微毛柃Eurya hebeclados等4个物种,重要值为7%~13%;其余46个树种重要值<5%,归类为群组Ⅲ,包括毛果珍珠花Lyonia ovalifoliavar.hebecarpa、多脉青冈Cyclobalanopsis multinervis、水青冈Fagus longipetiolata、红果山胡椒Lindera erythrocarpa和交让木Daphniphyllum macropodum等5个重要值排在前10位的树种,还有齿叶冬青Ilex crenata、湖北海棠Malus hupehensis、山樱花Cerasus serrulata等41个重要值<1%的树种。群组Ⅲ个体总数仅占27.7%,其中28个树种的个体数<10株,12个树种仅见1株,表明群组Ⅲ主要为群落伴生种和稀有种。

图1 天台鹅耳枥群落多度和胸高断面积累积Figure 1 Cumulative percentage of species abundance and area at breast in C. tientaiensis communities

表1 天台鹅耳枥所在群落重要值排前 10 位物种Table 1 Top 10 tree species of importance value in C. tientaiensis communities
2.2 天台鹅耳枥群落稳定性特征
对天台鹅耳枥所在群落所有木本植物的累积相对频度与乔木总物种数累积倒数作散点图。由图2可知:最优Godron稳定性指数曲线回归结果为Gompertz曲线(y=98.14×0.179.91x,R2=0.992 5),交点坐标离群落稳定点(20,80)较近。群落径级呈偏正态分布,群落中小径级个体较少(图3A),群落结构较为稳定。具体来看,群落中4 cm及以下胸径的个体仅占个体总数的2.09%,其中天台鹅耳枥占1.88%,为所有4 cm及以下胸径个体的90%;4~20 cm胸径的个体占94.32%,其中天台鹅耳枥占7.97%;20 cm以上胸径的个体占3.58%,天台鹅耳枥占0.72%。群落中胸径30 cm以上的个体共18株,其中天台鹅耳枥11株,占61.11%,最大胸径为74.6 cm,是群落中胸径最大的个体。但在天台鹅耳枥径级分布中,4 cm及以下胸径的个体仅占18.00%,无1 cm以下胸径的个体;4~20 cm胸径的个体占75.42%;30 cm以上胸径的个体占3.39%(图3B)。

图2 天台鹅耳枥所在群落的 Godron 稳定性Figure 2 Godron community stability in C. tientaiensis community

图3 天台鹅耳枥所在群落 (A)和天台鹅耳枥 (B)径级分布Figure 3 Distribution of diameter at breast height of the community (A) and C. tientaiensis (B)
2.3 天台鹅耳枥与群落内其他物种之间的关系
对天台鹅耳枥和群落内其他50个树种做种间联结分析。共同出现百分率(PC)分析(图4A)表明:垂丝卫矛Euonymus oxyphyllus、光叶山矾Symplocos lancifolia、闪光红山茶Camellia lucidissima、毛鸡爪槭Acer pubipalmatum、毛脉槭A.pubinerve、石灰花楸Sorbus folgneri等7个树种与天台鹅耳枥没有相关性,其他43个树种均与天台鹅耳枥表现出正相关性,其中PC>0.3的有6个树种。联结系数(AC)分析(图4B)表明:天台鹅耳枥与23个树种表现出负联结,与27个种对表现出正相关。χ2检验(图4C)表明:天台鹅耳枥与红果山胡椒、雷公鹅耳枥C.viminea、水青冈等树种极显著相关(P<0.01),与白檀、闪光红山茶等显著相关(P<0.05)。

图4 天台鹅耳枥与群落内主要树种之间的种间联结值Figure 4 Interspecific association between C. tientaiensis and other tree species
3 讨论与结论
天台山、大盘山和上山头,天台鹅耳枥均分布在黄壤、海拔相对较高(890~1 600 m)、坡度相对较大(15.35°~38.32°)、植被覆盖率较高的中坡至山脊上,种群更新能力差。受海拔较高影响,木本植物种类组成差异较大[6-7]。陈模舜等[7]发现:天台山和大盘山等地群落结构稳定性较差,受人为干扰的影响较大;天台山18株天台鹅耳枥在群落中的重要值排在第2位,仅次于柳杉;天台鹅耳枥自然分布植株的生长受伴生乔木的影响,同时也影响伴生树种的组成和物种多样性[6]。本研究发现:景宁上山头天台鹅耳枥所在群落结构稳定,海拔较高,受人为干扰影响较少;重要值、个体数量、胸高断面积以及群落乔木层树种重要值聚类分析等均显示:天台鹅耳枥是群落的优势树种之一。群落中4 cm及以下胸径的小径级个体和30 cm以上胸径的大径级个体在群落中具有显著的优势地位,但个体数量在其种群中的占比较小,分别占18.00%和3.39%,缺少1 cm以下胸径的个体,显示景宁上山头天台鹅耳枥存在更新限制,4~20 cm胸径的个体竞争压力较大。种间联结分析表明:与毛柄小勾儿茶Berchemiella wilsoniivar.pubipetiolata[1]、白 桂 木Artocarpus hypargyreus[2]、云 南 红 豆 杉Taxus yunnanensis[15]、脱皮榆Ulmus lamellosa[16]、狭果秤锤树Sinojackia rehderiana[17]、对 开 蕨Phyllitis scolopendrium[18]和藤枣Eleutharrhena macrocarpa[19]等濒危树种在群落内表现出独立分布或随机分布的特征不同,研究区天台鹅耳枥与群落中其他树种间存在较强的种间关系,这与黄梅秤锤树S.huangmeiensis[20]的研究结果一致。群落种间关系的形成与演替过程有关,在演替初期主要种对间尚未形成一定的种间关系,随着演替的进展,逐渐表现为复杂多样的种间关系[21],但在物种丰富的热带森林中可能会出现相反的结论[22]。另外种群数量的大小和分布特征可能也会影响濒危物种与群落内其他物种的关系,如数量少、分布零散比数量相对较多且集中分布的濒危物种,其种间关系较为松散。本研究中天台鹅耳枥与群落内其他树种较为复杂的种间关系可能与数量集中分布、群落较为成熟有关。因此,从群落生态学的角度来看,天台鹅耳枥可以在适生区内作为群落的优势种长期存在,应该具有更大的分布范围。运用气象和海拔等指标对该树种潜在分布区的预测[23]也证明:天台鹅耳枥可分布在浙江南部、台湾中南部以及安徽南部等高海拔区域。
天台鹅耳枥种子空壳率极高,种子人工萌发结果非常差,空气湿度和阴雨天气影响授粉成功率[8]。目前仅在2014年采集到饱满种子约500 g,繁育了一批实生苗,而扦插和组织培养等人工繁育的效果均不理想[12-13]。天台山、大盘山和上山头3个分布区的海拔均在800 m以上,高海拔地区较大的空气湿度和降水量可能是影响种子发育的关键因素。本研究中未发现1 cm以下胸径的个体,表明种子发育障碍以及从种子到幼苗阶段的更新限制是导致该树种数量稀少、分布狭窄的主要原因。种间联结分析还显示天台鹅耳枥与群落内其他树种间存在较强的竞争关系。
陈模舜等[10,24]研究发现:天台鹅耳枥及其幼苗具有一定的阳性特征。因此在合适的气象条件下采取人工授粉可提高种子发育饱满率;可采取间伐部分4~20 cm胸径的干扰树,通过减少其竞争压力来提高群落透光率,为天台鹅耳枥幼苗生长提供条件。
本研究表明天台鹅耳枥所在群落结构稳定,其在群落构建中发挥重要的作用,但存在种群更新障碍,且中龄级个体竞争压力较大。建议通过间伐等措施改善天台鹅耳枥生长环境,提高幼苗定植成功率。后续应进一步研究天台鹅耳枥繁育特性和繁育体系,以及中龄级个体的生态需求。
