平潭海域夜光藻对几种饵料藻的利用与生长
2022-08-17张海艳李晓东杨桂琴路玮静常阳王紫阳
张海艳,李晓东*,杨桂琴,路玮静,常阳,王紫阳
( 1.福建农林大学 海洋研究院 福建省海洋生物技术重点实验室,福建 福州 350002)
1 引言
有害藻华(Harmful Algal Blooms, HABs)指因微藻等生物大量繁殖,导致水生生物死亡,生态系统功能被破坏,甚至威胁人类健康和生命安全的一种生态灾害[1],其频发与人类社会发展导致的海洋富营养化与全球气候变化有着密切的联系[2-3]。夜光藻(Noctiluca scintillans)藻华是目前世界上分布最为广泛的有害藻华之一,各大洋近岸都有夜光藻暴发的记录,对当地环境与经济造成了不同程度的负面效应[4-5]。依据是否与夜光柄胞藻(Pedinomonas noctilucae)共生,夜光藻藻华分为红色与绿色两种,其中绿色藻华主要分布在中东-南亚沿岸,而红色藻华则分布在各大洋近岸海域,尤其以墨西哥湾、欧洲近海与东亚近海最为严重[4,6]。
夜光藻藻华同样是我国近岸常见的有害藻华之一[7-8],相关报道最早能够追述到1933年,20世纪80年代之后更是每年都有夜光藻藻华暴发的记录,其中渤海与东海近岸海域暴发最为频繁,并有暴发区域逐年扩大的趋势[7,9-11]。虽然在部分地区,如福建平潭与厦门,夜光藻形成的“蓝眼泪”现象已成为当地知名旅游景点,但是高密度夜光藻对沿岸环境与养殖业有着明显的不利影响,如1988年南海大鹏湾夜光藻藻华暴发,导致大量养殖鱼类死亡[12-13];2002−2003年,渤海与东海近岸夜光藻藻华导致水产养殖业损失总计超过1.3亿元[8]。国外也有夜光藻藻华暴发导致珊瑚缺氧死亡的报道[14]。但是现今对夜光藻藻华的形成机制认识尚不清楚。
目前对夜光藻藻华相关研究主要集中于对藻华现象的原位分析,重点关注现场气象、水文等条件对藻华形成与发展的影响[5,15-18]。万艳[19]认为,较高的水温(20.7~22.8℃)、弱风与低日照有利于平潭海域夜光藻藻华的发生;Xiang等[20]认为,盐度低、温度高、光照弱的环境,结合海水辐散上升的水文条件,更有利于夜光藻在海洋表层形成藻华;田达玮等[21]发现,胶州湾夜光藻密度除与水温呈显著负相关之外,还与水体叶绿素浓度呈显著正相关,说明该藻藻华和水体中食物丰度有关,这与尹翠玲等[22]对渤海湾夜光藻藻华生消过程与叶绿素浓度关系的研究结果相吻合。但目前对于夜光藻营养相关的研究较少,并且多集中在现场调查上,如Nishitani等[23]利用单细胞测序分析夜光藻食物泡中食物组成,Kitatsuji等[24]在显微镜下观察夜光藻食物泡中的藻类。实验室内定量的相关研究较少,仅有几种常见饵料藻(如扁藻、金藻等)或者甲藻(如链状裸甲藻Gymnodimium catenatum等)被探究是否能够被夜光藻摄食或利用[25-29],其中关于我国海洋中夜光藻对常见硅藻的摄食研究鲜见报道。
本实验选取福建平潭海域分离的夜光藻,在几种常见饵料藻和当地海域常见的硅藻作为单一食物来源的培养条件下,探究夜光藻增长速率的差异以及模拟海水混合对其增长速率的影响。同时探究在不同的营养盐水平下,夜光藻和扁藻共培养时种群动态的变化,并通过拟合初步构建夜光藻的种群增长模型。实验结果将会为东海近岸海域夜光藻暴发原因的探究提供科学依据。
2 材料与方法
2.1 藻类培养
夜光藻(N.scintillans)于2021年6月分离自福建平潭实验区(25°37′33″N,119°47′17″E,站位见图1),实验室条件下灭菌海水保存。亚心形扁藻(Platymonas subcordiformis)、三角褐指藻(Phaeodactylum tricornutum)、球等鞭金藻(Isochrysis galbana)、微小海链藻(Thalassiosira minima)、小球藻(Chlorellasp.)、骨条藻(Skeletonemasp.)由福建农林大学海洋研究院藻种库提供。以上藻种均使用f/2海水培养基[30]单种培养,海水 pH 为 8.09±0.05,盐度为 30±1,藻类培养环境温度为(20±1)℃,光照强度为(2 650±100)lx,光暗比为14 h∶10 h。自然海水使用0.45 μm混合纤维滤膜过滤,经121℃,25 min高温高压灭菌后用于藻类培养与后期实验。玻璃器皿使用蒸馏水冲洗3遍后经121℃,25 min高温高压灭菌后备用,量筒与塑料器皿经超纯水冲洗3遍后备用。
图1 夜光藻采样站位Fig.1 The sampling position of Noctiluca scintillans
2.2 不同藻类培养条件下夜光藻种群增长
实验分别设置对照组以及实验组,其中除夜光藻之外的6种微藻培养至指数生长后期,显微镜下计数后使用灭菌海水稀释并转移至24孔板中(实验体积为1 mL)作为实验组,每孔一种藻,每种藻终密度(生物量,以碳计)均为5.0×107pg/mL[31];亚心形扁藻液氮速冻10 min灭活后解冻,作为冷冻扁藻组(与实验组终密度相同);等体积的灭菌海水作为对照组;解剖镜下用吸管将夜光藻随机加入到以上各孔板中,每孔10个。每组设置3个平行。培养条件分别设置静置培养和模拟海水混合培养两种培养方式。上述实验组与对照组放置于培养环境下进行静置培养;制备相同实验组与对照组,将平板置于小型摇床上(120 r/min)培养,使得海水充分混匀从而模拟海水混合的情况,其他条件同静置培养组。每天在解剖镜下对夜光藻进行计数,持续20 d。藻类平均增长率采用Hallegraeff等[25]的公式:

式中,μ为平均增长率;t为培养时间;Nt为第t天的藻种密度;N0为实验开始藻种密度。
2.3 不同营养盐条件下的微藻种群动态分析
夜光藻与扁藻分别在解剖镜和显微镜下计数,无菌海水稀释后置入150 mL锥形瓶中(实验体积为50 mL),终密度分别为 1 cells/mL 与 1×104cells/mL,实验组设置营养盐浓度分别为f/2、f/4、f/8、f/16、f/32海水培养基,无菌海水作为对照。每组设置3个平行,放置于藻类培养环境下进行培养,每2 d分别在显微镜与解剖镜下对两种藻类的密度进行计数,持续20 d。
2.4 夜光藻种群动态模拟
将2.3节中所得夜光藻种群动态数据导入到Ori-gin pro中进行非线性曲线拟合,初步比较后选取Boltzmann 拟合(式(2)),拟合采取 Levenherg-Marquardt优化算法,最大迭代400次。对拟合得到的结果进行进一步的分析,分别采用高斯拟合(式(3))与线性拟合(式(4),式(5))探究营养盐水平与夜光藻种群动态参数之间的关系。
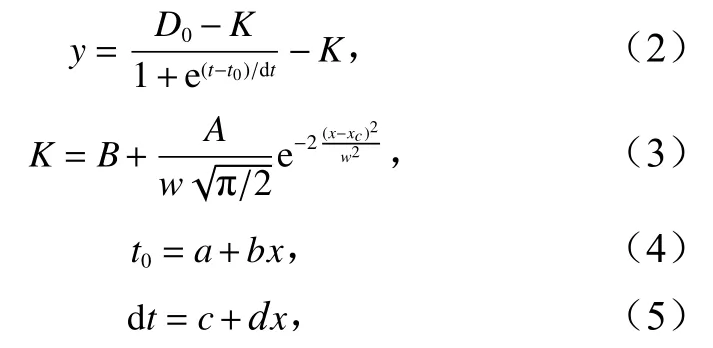
式中,y为夜光藻密度(单位:cells/mL);D0为夜光藻初始密度(1 cells/mL);K为环境容纳量(单位:cells/mL);t为培养时间;t0为达到K/2的培养时间;dt为时间系数;B为基线值;A为曲线下面积;w为σ置信区间;x为营养盐稀释系数;xc为K最大时x的取值;a,c为直线截距;b,d为直线斜率。
3 结果
3.1 不同藻类培养条件下夜光藻种群增长
如图2所示,静置条件下夜光藻在亚心形扁藻组中生长最为快速,在 20 d 的培养期内,夜光藻由(10±0)个上升到(205±4)个 ,平均增长率为(0.151±0.001)d−1(表1),显著高于其他实验组。而冷冻扁藻和骨条藻组中夜光藻在相同的时间内分别增加至(84±8)个与(86±5)个 ,平均增长率分别为(0.106±0.005)d−1与(0.108±0.003)d−1,显著低于亚心形扁藻培养组(p<0.01)。而在其他微藻作为唯一食物来源的情况下,夜光藻基本没有增长,和空白对照组之间没有显著的差异(p>0.05)。这表明静置条件下,夜光藻并不能够利用球等鞭金藻、三角褐指藻、小球藻、微小海链藻这4种藻作为唯一食物来源。

图2 静置条件下不同藻类培养的夜光藻种群增长Fig.2 Population growth of Noctiluca scintillans feeding by different algae in a standing situation
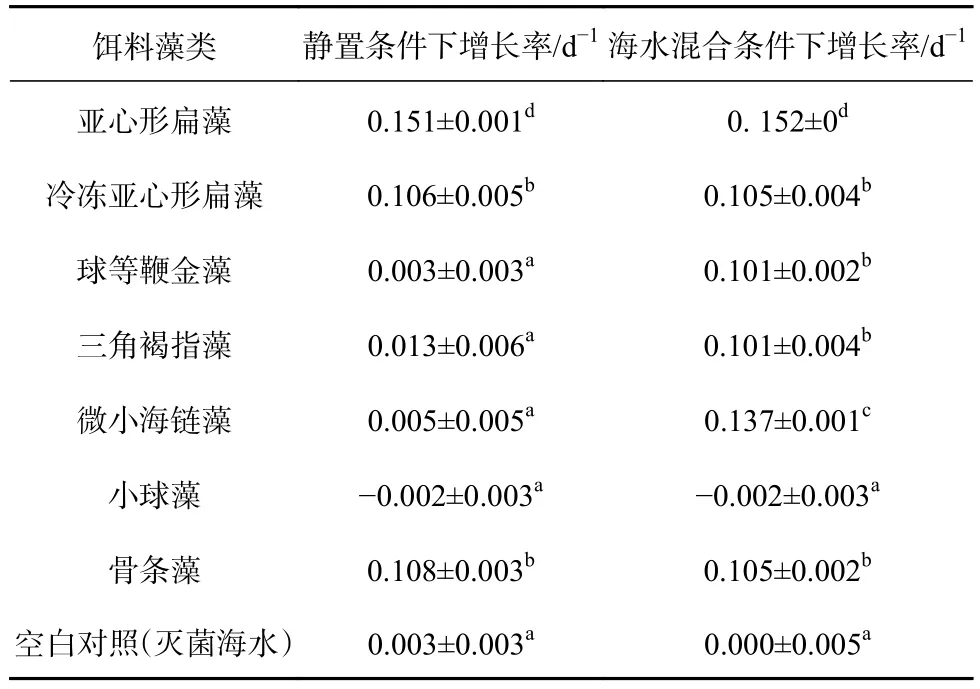
表1 不同培养条件下夜光藻种群平均增长率Table 1 Mean growth rate of Noctiluca scintillans feeding by different algal in standing and mixing simulated situation
如图3所示,在模拟海水混合的条件下,夜光藻对不同饵料藻的利用出现了明显的变化。虽然夜光藻在亚心形扁藻培养组的生长仍然最快,但和静置条件下并没有显著的差异(p>0.05)。冷冻亚心形扁藻培养组和骨条藻培养组与静置条件下的没有显著差异(p>0.05)。但微小海链藻培养组中夜光藻在20 d内上升至(153±4)个,平均增长率为(0.137±0.001)d−1,显著高于除亚心形扁藻组之外其他实验组(p<0.01)。在海水混合条件下,三角褐指藻和球等鞭金藻同样能够作为夜光藻的食物支持其生长,这和静置条件下有着显著区别(p<0.01)。这表明海水混合可能在夜光藻摄食利用浮游藻类的过程中起着重要作用。与此同时,海水混合条件下,小球藻培养组中夜光藻的种群数量依然没有增长,这说明夜光藻可能不能够利用小球藻。

图3 海水混合条件下不同藻类培养的夜光藻种群增长Fig.3 Population growth of Noctiluca scintillans feeding by different algal in a seawater mixing simulated situation
3.2 不同营养盐条件下的微藻种群动态
如图4所示,夜光藻和亚心形扁藻的种群增长曲线在不同营养盐条件下有着明显的差异。在f/2培养基培养的情况下,扁藻在10 d内迅速生长至(1.38±0.09)×106cells/mL,期间平均增长速率为(0.493±0.006)d−1,显著高于其他实验组(p<0.01),但是该营养盐条件下夜光藻生长缓慢,20 d内密度由(1±0)cells/mL增长至(37±2)cells/mL,和对照组夜光藻密度((39±6)cells/mL)之间并没有显著差异(p>0.05)。随着营养盐浓度的下降,亚心形扁藻种群平均增长速率显著降低(p<0.01),低于f/2营养盐水平的实验组中,除f/4实验组中扁藻最高密度达到(5.83±0.32)×105cells/mL,其余实验组中扁藻密度都在短暂上升之后迅速回落到1×105~2×105cells/mL的水平。夜光藻则在除f/2组的其他组中逐渐占据优势。
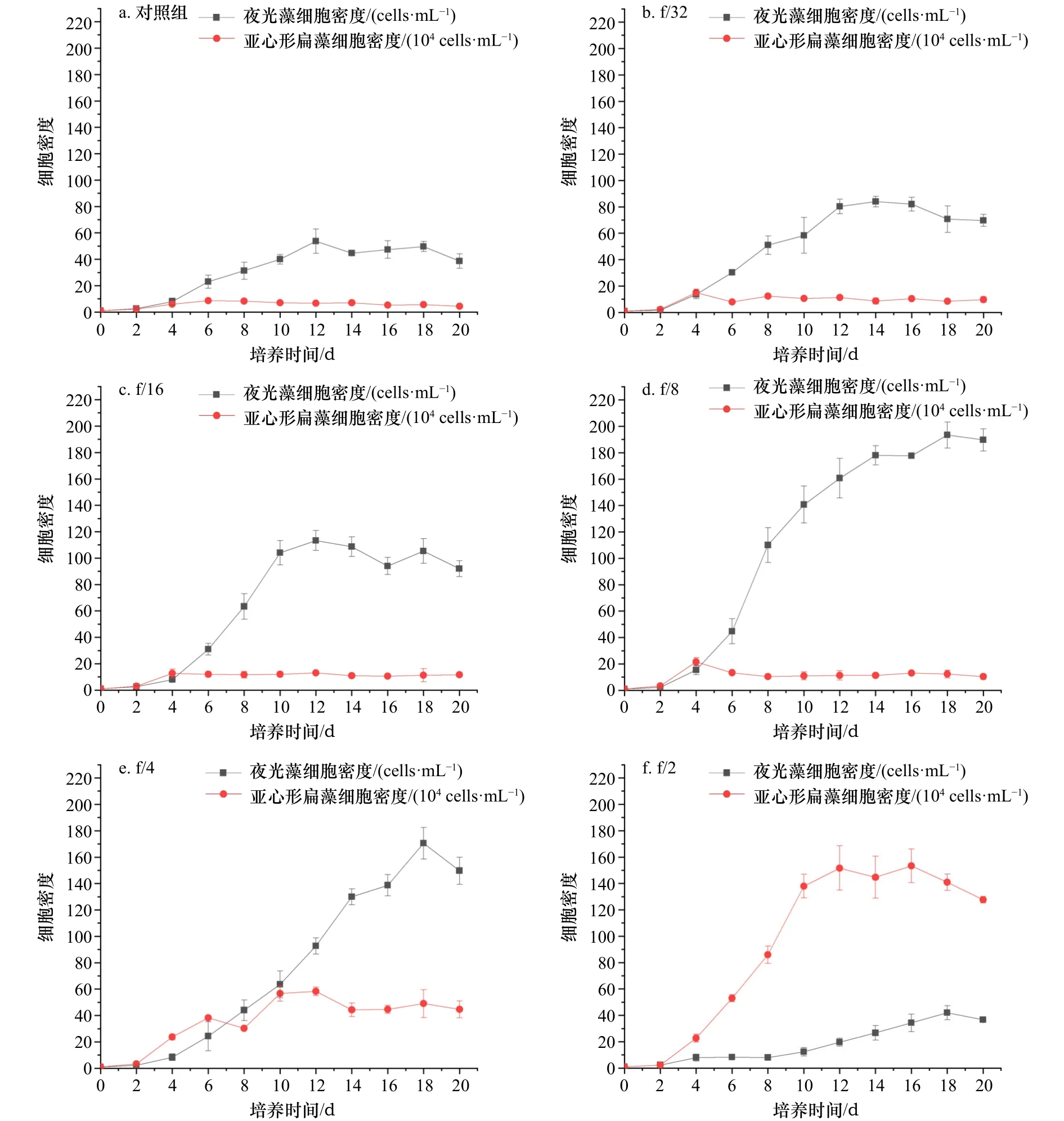
图4 夜光藻和亚心形扁藻在不同营养条件下的种群增长曲线Fig.4 Population growth curves of Noctiluca scintillans and Platymonas subcordiformis under different nutritional conditions
结合拟合结果(图5),各实验组中夜光藻种群的动态变化较好地符合Boltzmann增长模型,各实验组与对照组R2都在0.9以上(表2)。随着营养盐水平下降,夜光藻的种群增长出现先增强后减弱的趋势,其中最大环境容纳量K的峰值((184.65±3.19)cells/mL)出现在f/8组,和实际情况下夜光藻密度的最大值((193±10)cells/mL)并没有显著性的差异(p>0.05)。拟合结果还表明,夜光藻种群增长最快时间t0与时间系数dt都随着营养盐水平的降低而出现降低的趋势。进一步对Boltzmann增长模型中各参数与营养盐稀释系数进行相关性拟合之后,结果表明K、t0与dt与其有着较强的相关性,分别符合高斯分布、线性分布与线性分布,R2分别为 0.994 0、0.941 7、0.863 0(表3)。这表明在与亚心形扁藻的共培养体系中,夜光藻的种群增长表现为受单一因素(即营养盐水平)影响的函数,两种藻类表现出互相抑制的特点,其抑制能力与营养盐水平相关,高营养盐水平反而不利于共培养条件下夜光藻的生长。
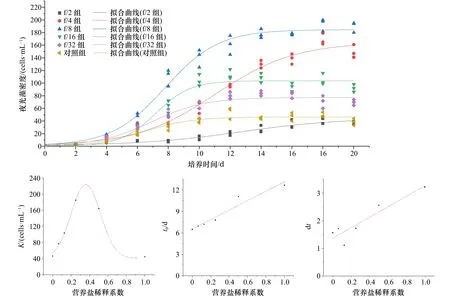
图5 夜光藻种群动态拟合Fig.5 Fitting of population dynamics of Noctiluca scintillans

表2 夜光藻种群动态Boltzmann拟合参数Table 2 Parameters of Boltzmann fitting on population dynamics of Noctiluca scintillans

表3 夜光藻种群动态Boltzmann拟合参数与营养盐稀释倍数的拟合Table 3 Fitting of dilution ratio with parameters of Boltzmann fitting on population dynamics of Noctiluca scintillans
4 讨论
夜光藻生长和其食物来源有着明显的关系,Hallegraeff等[25]发现不同藻类喂养下,夜光藻平均增长率差异(0~0.69 d−1)显著,但不同实验组之间饵料生物量差异较大,横向比较较为困难。本实验结果表明,即使在相同生物量情况下,不同微藻培养的夜光藻种群增长之间依然存在显著差别。较于其他微藻,亚心形扁藻组中夜光藻平均增长率更高,说明该藻可能是夜光藻较好的食物。这与之前的研究结果相似[27-28,32]。相较于活细胞,冷冻死亡的亚心形扁藻投喂下的夜光藻增长率显著下降,说明死亡藻体(或其滋养的细菌)并不是夜光藻较好的食物。海水混匀条件下的微小海链藻同样能够支持夜光藻较高的增长率,这和当前已有的海链藻能够培养夜光藻的结论相符[33],如Hallegraeff等[25]发现,伪矮海链藻(Thalassiosira pseudonana)培养下夜光藻的平均增长率为0.16~0.23 d-1,与本实验微小海链藻组夜光藻的增长率较为相似。骨条藻、球等鞭金藻与三角褐指藻在一定条件下虽然能够维持夜光藻种群增长,但是增长率较低,且相互之间并没有显著的差别。这说明夜光藻在食物质量较差的情况下也能够维持低水平的种群增长,这同样与现有结果相吻合[28,33-34]。但Gomes等[35]发现,绿色夜光藻在三角褐指藻培养下增长率比等生物量威氏海链藻(Thalassiosira weissflogii)培养下的增长率要高,这与本实验结果和之前其他的研究结果相差较大,说明混合营养的绿色夜光藻和完全异养的红色夜光藻对食物的选择和利用可能存在差别。
夜光藻对饵料利用上的差异有着多种解释。Nakamura[34]认为,夜光藻在不同饵料投喂下增长率的差异与饵料藻大小(等效直径ESD)有关,ESD小于5 μm的衣藻(Chlamydomonas parkeae)与球等鞭金藻不能支持夜光藻生长,而较大的饵料(ESD>10 μm)投喂下夜光藻增长率较高,这同样与本文的结果相符,亚心形扁藻与微小海链藻直径均大于10 μm,而其他藻类都比较小。徐韧等[32]发现,种群增长率高的实验组中夜光藻食物泡多而充盈,而增长率低的实验组中夜光藻食物泡较少,认为增长率差异可能与其摄食偏好相关。但现有研究表明,虽然夜光藻对不同食物的摄食率存在显著差异,但高摄食率并不等同于高增长率[26-27,36],如夜光藻对海洋丝尾虫(Uronema marinum)摄食率显著高于其他食物,但其增长率反而是负值[27]。本实验也发现,静置条件下球等鞭金藻组中夜光藻在解剖镜下能观察到多而充盈的食物泡,但种群几乎没有增长,这说明夜光藻的生长和其对食物的利用更为相关。Zhang等[36]发现夜光藻在混合营养的情况下,短期摄食率和饵料生物量呈正相关,但长期更趋近于摄取营养价值更高的食物,而且营养条件充足的藻类培养下夜光藻的增长率显著高于其在等密度的营养缺乏(氮磷不足)的同种藻类培养下的增长率。这表明食物的营养价值在很大程度上决定了夜光藻的生长。
海水混合可能对夜光藻的生长有着重要作用,模拟海水混合条件下,部分实验组中夜光藻平均增长率显著上升,以微小海链藻组最为明显。观察发现静置条件下三角褐指藻和微小海链藻有明显的沉淀,而海水混合条件下则处于悬浮状态,这表明海水混合可能通过增加藻之间接触几率从而提高夜光藻的摄食率。观察还发现混合条件下夜光藻更容易形成集群。集群的夜光藻能够分泌黏性物质形成黏液网,更容易黏住食物,提高摄食率,从而获得更高的增长率[37-38]。海水混合导致夜光藻增长率的变化可能有助于解释平潭海域多发的藻华现象,该地区海流较强,海水混合较为强烈,为夜光藻利用海洋中硅藻等作为食物提供了良好条件。环境检测数据表明,夜光藻藻华多发的时间(3月底至5月中旬)与该海域硅藻藻华发生的时间互相重叠,同区域可能同时出现夜光藻藻华与微小海链藻藻华(福建政府数据https://hyyyj.fujian.gov.cn/xxgk/tzgg/),而夜光藻藻华和前期硅藻藻华有着密切的关系[5,20,39-40],Kitatsuji等[24]对藻华发生区的夜光藻镜检发现,其食物泡里面主要为角毛藻属、海链藻属等硅藻。结合本文实验结果表明,微小海链藻等硅藻在平潭海域春季的大量繁殖可能是导致该海域夜光藻藻华暴发的原因之一。
近海海水富营养化是导致浮游植物大量繁殖的主要原因之一,为了探究营养盐水平对夜光藻藻华的贡献,本文探究了在不同营养盐条件下,夜光藻与最佳饵料亚心形扁藻的种群动态变化。结果表明,在共培养体系下,夜光藻的种群增长与营养盐水平有关,周成旭等[29]同样发现,添加较低浓度营养盐的培养基(f/40)能够促进夜光藻-扁藻共培养体系中夜光藻的种群增长。拟合结果表明,夜光藻种群动态符合Boltzmann增长模型,其各参数都与营养盐水平有较强的相关性,这说明无机营养水平能够影响夜光藻种群的增长速度与环境容纳量。但是作为完全异养性的藻类,夜光藻种群动态与营养盐水平的相关性属于间接相关,Zhang等[36]认为,夜光藻种群增长率随饵料密度上升而上升,但同时存在边际效应。实验中夜光藻只有在扁藻种群增长趋近于0的时候开始增长,说明高增长率与高密度的饵料藻对夜光藻增长可能有抑制作用,这可能是与亚心形扁藻快速生长时产生他感作用有关。这表明夜光藻的种群动态可能是亚心形扁藻的营养价值与抑制能力共同作用的结果,而这两个因素都与营养盐水平相关,这就导致最终夜光藻种群动态表现为受营养盐单一因素影响的函数。这种现象可能为微藻-夜光藻藻华演替提供一个可能的解释,很多原位观察表明,夜光藻藻华往往跟随在硅藻等浮游植物大量繁殖之后[20,24,41],这可能是夜光藻的生长在硅藻大量繁殖的时候受到一定的抑制,直到硅藻繁殖进入后期之后才开始大量繁殖。营养盐的变化可能显著改变微藻-夜光藻的种群动态关系,从而对夜光藻藻华的形成起到重要的作用,而陆源无机营养输入可能在夜光藻藻华形成机制中有着重要的地位。
5 结论
本文探究了福建平潭海域夜光藻在几种饵料藻与硅藻培养条件下的生长情况,结果表明不同饵料投喂下夜光藻的增长率有显著差异,其中亚心形扁藻培养下的夜光藻生长最快。海水混合对夜光藻生长有着重要的作用,模拟混合条件下,夜光藻能够利用微小海链藻等硅藻快速生长。营养盐水平对“夜光藻-亚心形扁藻”共培养体系中两种藻类的种群动态有着显著的影响,其中夜光藻种群增长符合Boltzmann模型,其各项参数与营养盐水平显著相关。实验结果表明,夜光藻对不同饵料藻的利用存在显著的差异,营养盐水平可能在夜光藻藻华形成机制中起着重要的作用。
