荣成马山里海域海草床分布现状及其生态特征
2022-08-17邓筱凡张宏瑜吴忠迅李文涛张沛东
邓筱凡,张宏瑜,吴忠迅,李文涛,张沛东*
( 1.中国海洋大学 海水养殖教育部重点实验室,山东 青岛 266003;2.长岛国家海洋公园管理中心,山东 烟台 265800)
1 引言
海草是一类可完全生活在海水中的被子植物,是浅海水域重要的初级生产者,年净生产力(干重,下同)达1 012 g/m2,由其构筑的海草床是地球最有效的碳封存系统之一,碳储量可达海洋碳储量的10%以上,其还具有净化水质、防风固堤、稳定底质等重要的生态功能[1-3]。然而,近几十年来受自然环境变化和人类活动的影响,全球海草床面积急剧衰退,超过1/3的海草床已完全消失,海草床的有效保护与高效修复亟待开展[4-5]。
中国近海有海草22种,约占全球海草种数的30%[6]。我国海草床的退化状况非常严峻。例如,胶州湾芙蓉岛海域海草床由20世纪80年代的1 334 hm2退化至目前仅有零星分布[7];1990年前威海沿岸广布鳗草(Zostera marina)平均生物量(干重,下同)达1 500 g/m2,但2016年调查显示,其平均生物量降至约504 g/m2,仅为1990年平均生物量的1/3左右[8-11];2017年广东省流沙湾海域海草床的分布面积和平均生物量分别仅为26.90 hm2和3.74 g/m2,均不足2002年的1/10[12];2015−2019 年,海南岛海草床的海草盖度由25.9%下降到16.7%,平均茎枝密度由1 035 shoots/m2降至579 shoots/m2,平均生物量下降36.1%[13]。
明确海草床的海草种类和面积等生态分布状况,是海草床保护与恢复的基础。近年来,我国学者对温带海域山东省的威海沿海[10,12-13]、莱州湾[14]、黄河口[15],河北省的曹妃甸[16-17],辽宁省的獐子岛[18]、长山岛[19]、兴城−觉华岛[20]等海区以及亚热带和热带海域广西壮族自治区的竹山[21]、合浦[12]、北部湾[22],广东省的柘林湾[23]、流沙湾[24],海南省的黎安港[25]、东塞港[26]、高隆湾[27],台湾省沿海[28],香港特别行政区的下白泥、荔枝窝[6]等海区的海草床分布进行了调查,逐步探明我国海草资源现状。然而,仍有部分海域的海草床分布状况至今缺少基础数据,严重影响了对海草资源的评估与保护工作。
荣 成 马 山 里 海 域 (37.30°~ 37.34°N, 122.56°~122.64°E)位于山东半岛东端,介于天鹅湖和养鱼池湾之间,其沿岸村落大量存在的海草房证明该海域曾广布海草床。为查明马山里海域海草床的分布现状及其生态特征,于2015年10月对马山里海域的海草碎屑开展调查,发现岸边堆积大量海草干叶,结合渔民走访,初步判定马山里海域分布有较大面积的海草床。随后,于2016年8月对马山里海域的海草床开展系统调查,并通过分析荣成马山里海域海草床可能面临的主要威胁因素,提出相应的保护和修复建议,以期为我国海草分布状况和海草生态学研究提供基础资料,也为我国温带海草床的科学管理提供参考。
2 调查方法
2.1 海草种类和分布调查
对马山里6 m以浅的海域进行大面调查,设置21条断面,间隔30 m,调查总面积为1 092 hm2,如图1所示。现场调查选择大潮低潮时段,由潜水员携带象拔潜水观察,发现有海草分布后沿草床边界间隔释放象拔,并使用水下相机拍摄记录,以海草覆盖度不小于5%为草床边界划分标准[11]。调查船上人员发现象拔后,用GPS记录边界拐点坐标,在记录本绘制海草分布示意图,标注近岸标志性建筑位置。返回实验室后,对拍摄录像进行判读分析,结合定点坐标和实时卫星测距,估算海草床分布面积。

图1 马山里海域海草床调查区域Fig.1 Investigation area of seagrass beds in the Mashanli sea area
2.2 海草生物学特征调查
在海草床分布区设置10个采样站位,使用25 cm×25 cm样方框采样,每站位设置3组重复。将样方内所有海草植株完整取出,冲洗干净,置入样品袋,低温保存,带回实验室进行测定与分析。
首先通过外部形态和组织切片等方式鉴定海草种类;随后统计各样方植株数量,计算茎枝密度;然后在每个站位随机选取15株完整植株,将其表面附着物与根部泥沙冲洗干净,进行形态学测量,包括株高、叶鞘长、叶鞘宽、叶宽和最大根长,其中株高、叶鞘长和最大根长使用直尺测量,叶鞘宽和叶宽使用游标卡尺测量;最后,将样方内的全部海草植株分为地上组织和地下组织,于60℃恒温烘干至恒重,测定生物量。测量标准和计算方法如下:(1)株高(cm):自分生组织至最长叶片顶端的高度;(2)叶鞘长(cm):包围着茎的叶基部的长度;(3)叶鞘宽(cm):叶鞘中间位置的宽度;(4)叶宽(cm):最长叶片中间位置的宽度;(5)最大根长(cm):单株最长根的长度;(6)单株地上生物量(g/shoot):单株地上组织干重;(7)单株地下生物量(g/shoot):单株地下组织干重。
2.3 海草床关键环境因子调查
在各站位,使用YSI 650(美国YSI公司)现场测定水温、盐度、pH和溶解氧含量;使用SM-5A测深仪(美国Speedteach公司)测量水深;使用SD20海水透明度盘(北京普力特仪器有限公司)测量透明度。沉积物理化性质主要监测的是沉积物的粒度和有机质含量,样品采集方法按照《海洋调查规范 第8部分:海洋地质地球物理调查》(GB/T 12763.8−2007)规定执行,粒度分析采用筛析法,有机质含量的测定采用烧失量法[29-31]。
2.4 海草床主要威胁的调查与分析
对马山里海域及天鹅湖、养鱼池湾周边的沿岸环境进行调研。通过现场调查,对空地、耕地、居民区、工厂以及养殖场等不同环境类型的位置和规模进行统计;通过对相关公司和渔民的走访,了解马山里海域海水养殖种类规模、捕捞渔船网具规格、码头建设位置、航道路线及其变迁等具体信息。
2.5 数据处理与分析
运用ArcGIS 10.2进行海草分布状况的绘图和海草分布面积的估算。运用SPSS 25.0进行描述性统计分析、独立样本t检验、聚类分析(CA)和典型相关分析(CCA),绘制系谱图。利用R语言的Corrplot软件包中进行Spearman相关性分析,确定海草形态学特征指标和海区环境因子之间的潜在相关性。利用ArcGIS 10.2克里金插值法评估海草生长状况和周边环境类型,分析海草床潜在威胁来源。实验数据用平均值±标准误表示,显著性水平为p<0.05。
3 结果与分析
3.1 海草种类、分布范围与面积
通过植株形态学观察和组织切片分析,发现荣成马山里海域共分布2种海草,隶属于1科2属,分别是鳗草科虾形草属的红纤维虾形草(Phyllospadix iwatensis)(图2)和鳗草属的丛生鳗草(Zostera caespitosa)(图3)。马山里海域海草床总面积为58.26 hm2,其中红纤维虾形草分布面积为54.50 hm2,占该海域海草床总面积的93.5%,丛生鳗草分布面积为3.76 hm2,占海草床总面积的6.5%。

图2 红纤维虾形草水下状态(a)、外部形态特征(b)及其叶片组织切片(c)Fig.2 Underwater state (a), external morphological characteristics (b) and leaf transection (c) of Phyllospadix iwatensis

图3 丛生鳗草水下状态(a)、外部形态特征(b)及其叶片组织切片(c)Fig.3 Underwater state (a), external morphological characteristics (b) and leaf transection (c) of Zostera caespitosa
红纤维虾形草海草床分为北部和南部两个带状区域,北部分布区宽约85 m,自西向东延伸约855 m,分布面积约为6.55 hm2;南部分布区面积较大,宽约173 m,自东北向西南方向延伸约3 500 m,至养鱼池湾湾口码头处消失,分布面积约为47.95 hm2。丛生鳗草呈现斑块状分布,与红纤维虾形草交错而生,如图4所示。由此可见,马山里海域海草床以红纤维虾形草为主。

图4 马山里海域海草分布Fig.4 Distribution of seagrasses in the Mashanli sea area
3.2 海草的茎枝密度、生物量与形态学特征
调查显示,马山里红纤维虾形草平均茎枝密度为(368.0±18.2)shoots/m2,平均生物量为(297.0±41.5)g/m2,平均株高为(89.2±4.2)cm;丛生鳗草平均茎枝密度为(691.2±17.1)shoots/m2,平均生物量为(534.0±70.7)g/m2,平均株高为(78.4±8.3)cm。形态学特征如表1所示。
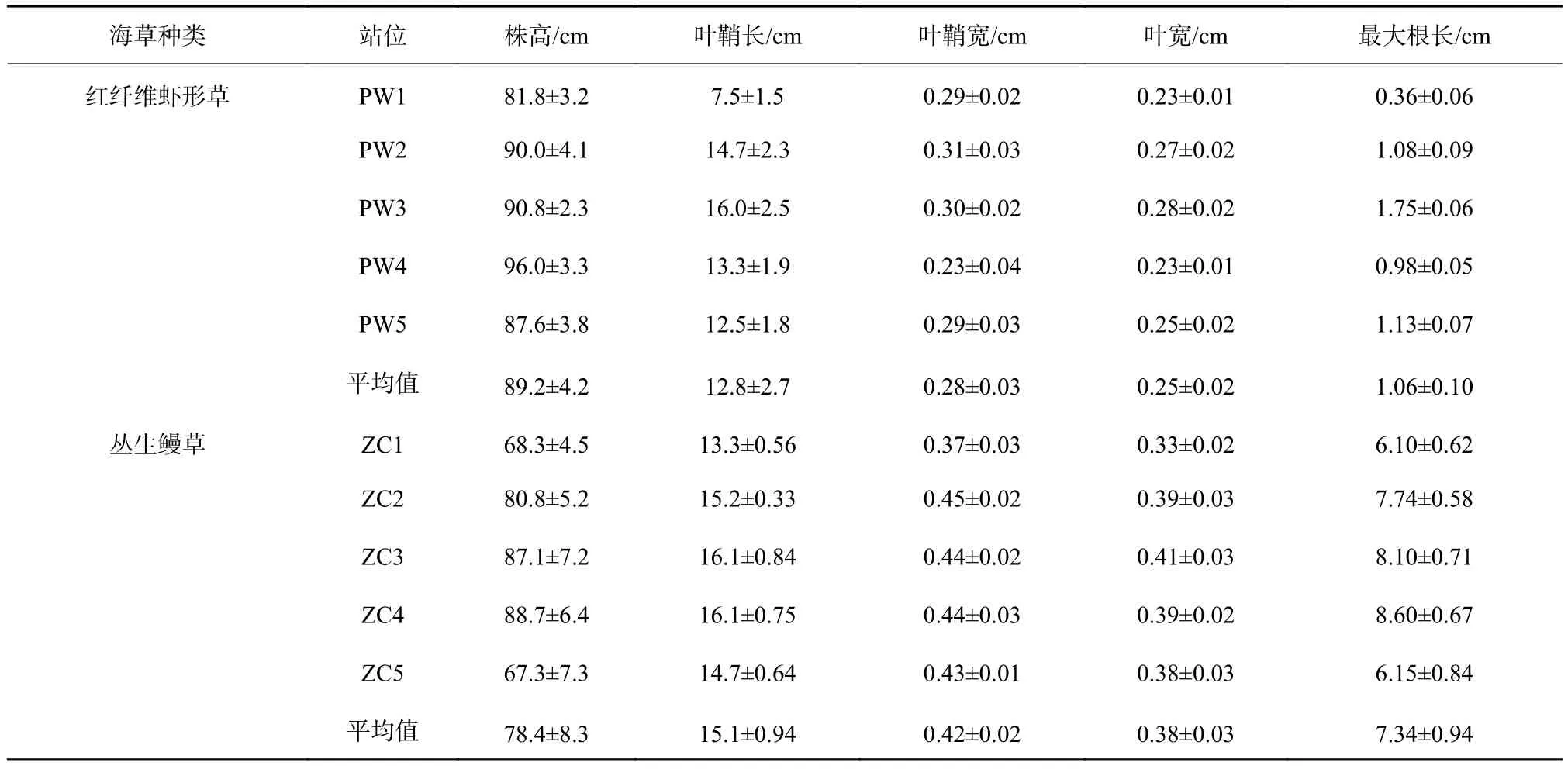
表1 马山里海域海草形态学指标统计Table 1 Statistics of morphological indexes of seagrasses in the Mashanli sea area
3.3 环境因子
监测结果发现,海草床分布水体平均水深为(2.8±0.3) m,透明度为(1.7±0.2) m,盐度为(31.7±0.2),底质以岩礁底质为主,间有少量黏土质砂,沉积物平均有机质含量为(1.12±0.05)%,如表2所示。海草生境地形复杂多样,有大量的海底凸起和凹陷。

表2 马山里海域海草床主要环境因子Table 2 The main environmental factors of the seagrass beds in the Mashanli sea area
3.4 海草生长状况分析
对马山里海域2种海草的主要形态学特征指标进行独立样本t检验,发现2种海草的株高、叶鞘长、单株地上生物量和单株地下生物量等差异不显著(p>0.05),故选择这4个指标对马山里海域10个站位的海草形态学特征指标进行均值聚类分析,如图5所示。马山里海域海草可按生长状况分为两个聚类中心,第一类为生长较好,包括 PW2、ZC3、ZC4、PW3、PW4、ZC2、ZC1共7个站位,第二类为生长较差,包括PW1、PW5和ZC5共3个站位,两者最终聚类中心的距离为20.086。第一类的4个形态学特征指标的平均值均高于第二类,是第二类的1.1~1.4倍。根据海草形态学特征指标谱系绘制马山里海域海草生长状况评估图(图6),可以看出,马山里海域中部海草长势优于南北两侧。

图5 海草形态学特征指标谱系Fig.5 Morphologic indexes intergroup pedigree of seagrasses
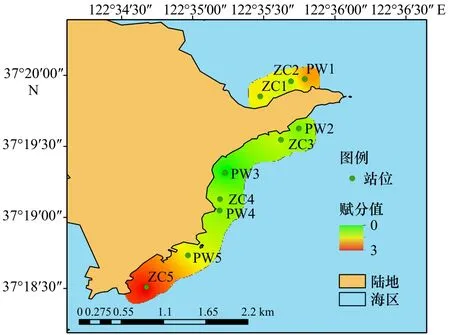
图6 马山里海域海草调查站位及生长状况评估结果Fig.6 Seagrass survey stations and evaluation results of growth status in the Mashanli sea area
3.5 海草生长影响因子分析
3.5.1 Spearman相关性分析
对马山里海域海草形态学特征和环境因子进行Spearman相关分析,结果表明,海草形态学指标间具有很强的相关性(图7A),如单株地上生物量(AB)与株高(SH),单株地上生物量(AB)、单株地下生物量(UB)与叶鞘长(LSL)均为显著正相关(p<0.05);同时环境因子之间也有很强的相关性(图7B),如水温(WT)、溶解氧含量(DO)、透明度(SWT)、沉积物有机质含量(OM)之间均有很强的相关性。然而,Spearman相关分析发现,海草形态学特征指标与环境因子指标之间无显著相关性(图7C),这可能是因为分析方法不适用。因此,进一步采用典型相关分析探究海草形态学特征指标和环境因子指标之间的相关性。

图7 海草形态学指标与环境因子间的相关性分析Fig.7 Correlation analysis among morphological indexes of seagrass and environmental factors
3.5.2 典型相关分析
对海草的全部生态学特征指标与环境因子指标进行典型相关分析,结果发现,典型变量的相关性系数较低,无法以典型变量之间的关系表示两类指标之间的关系。然后进行了海草的不同生态学特征指标与不同环境因子指标的多种组合,并分别开展典型相关分析,发现株高、叶鞘长、茎枝密度和单株生物量等海草生态学特征指标和水深、水温、透明度和沉积物有机质含量等环境因子指标的组合,第一典型变量相关性系数较高,可以通过第一典型变量之间的关系表示两类指标之间的关系,而且这8个海草的生态学特征指标与环境因子指标亦能够较好反映海草的生长分布和环境状况(图8)。
典型相关分析共得到4对典型变量(ρ1(U1、V1)、ρ2(U2、V2)、ρ3(U3、V3)和ρ4(U4、V4)),其相关系数分别为0.962、0.556、0.451和0.126。第一典型变量间的相关系数为0.962(p<0.05),因此海草形态学指标与海区环境因子之间相关性分析可简化为只研究第一对典型变量ρ1(U1、V1)之间的关系。其中,U1为海草生态学的综合指标,V1为海区环境因子的综合指标。U1、V1标准化典型结构如图8所示。

图8 U1、V1典型结构示意图Fig.8 Canonical correlation structure diagram of U1 and V1
第一典型变量与原始变量转换公式为
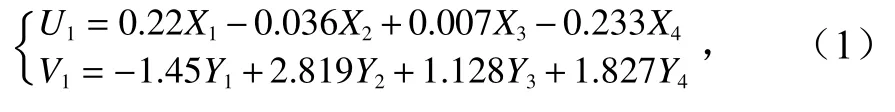
式中,U1为生态学指标第一典型变量;V1为海区环境指标第一典型变量;X1为株高;X2为叶鞘长;X3为茎枝密度;X4为单株干重;Y1为水深;Y2为水温;Y3为透明度;Y4为沉积物有机质含量。
典型载荷分析表明,在海草生态学指标中,茎枝密度典型负载系数高,达0.984,为U1主影响因子,故茎枝密度为生态学指标的主要影响因素;在环境指标中,水温和水深典型载荷系数均较高,分别为0.957和−0.873,为V1主影响因子,即水温和水深为环境指标的主要影响因素(图9a)。
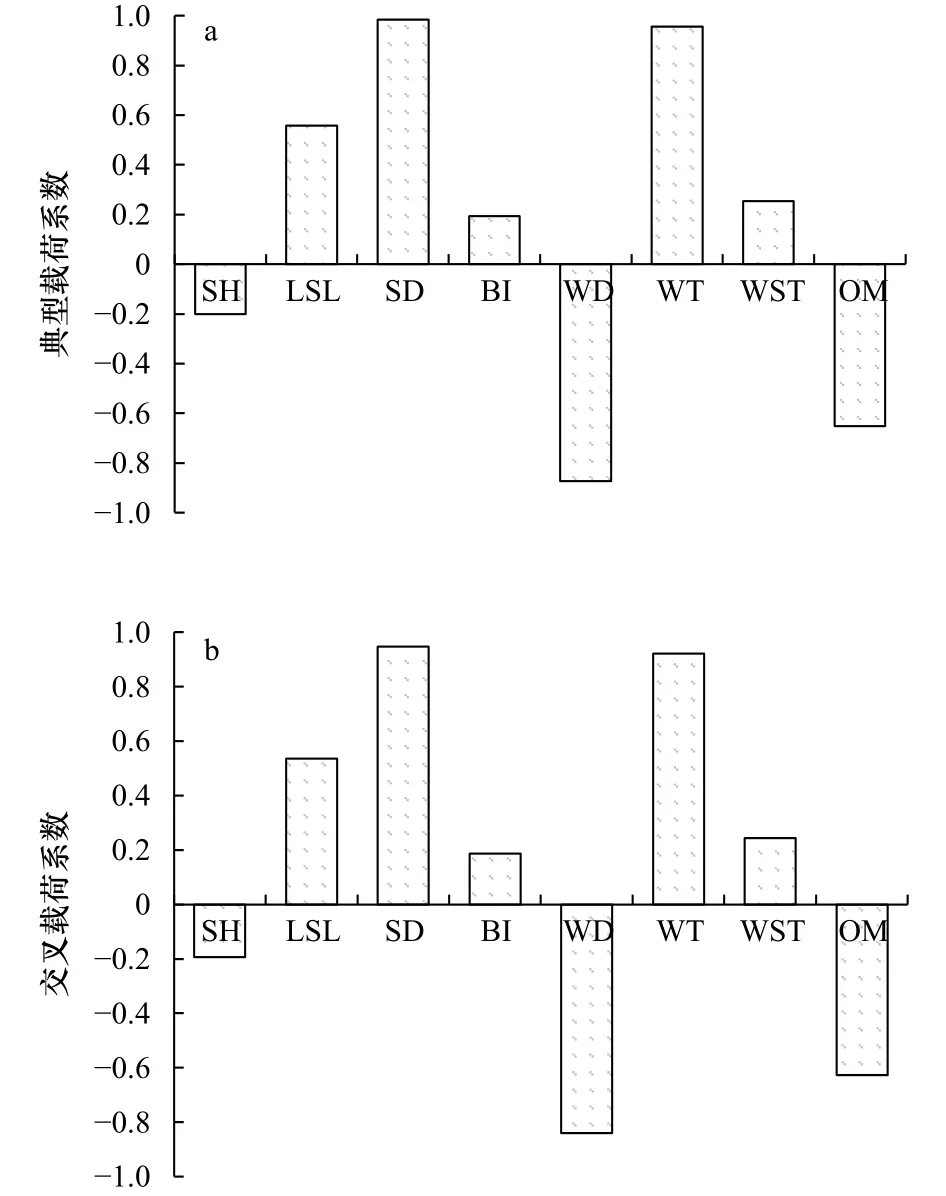
图9 变量典型载荷分析(a)和变量交叉载荷分析(b)Fig.9 Variable typical load analysis (a) and variable cross load analysis (b)
交叉载荷分析表明,U1对V1的影响中,茎枝密度交叉载荷系数较高,为0.947,V1受其影响较大,且为正相关;V1对U1的影响中,水温和水深的交叉载荷系数较高,分别为0.921和−0.840,U1受这两个环境指标的影响较大,且U1与水温为正相关,与水深为负相关(图9b)。
综上,夏季马山里海域,温度较高以及水深较浅的区域海草生长状况较好,海草茎枝密度较高。
典型冗余分析组内解释度表明,第一典型变量U1可解释4个海草生态学指标33.9%的组内变异,第一典型变量V1可解释4个海洋环境指标54.2%的组内变异,如图10所示。

图10 典型冗余分析组内、组间解释度Fig.10 Interpretation of typical redundancy analysis within and between groups
典型冗余分析组间解释度表明,环境第一典型变量V1对海草形态学特征的解释度为31.4%,4个环境典型变量(V1−V4)对海草形态特征的解释度共计41.2%,说明马山里海域还存在其他因素影响海草生长及分布状况;生态指标第一典型变量U1对海区环境因子的解释度为50.2%,4个生态典型变量(U1−U4)对海区环境因子的解释度共计55.3%,表明海草床对海区环境具有一定的影响作用,但影响程度一般(图10)。
综上,马山里海域海草床分布区中,水温和水深是影响海草床茎枝密度等生态学指标的关键因子,海草的生长状况与水温呈正相关,与水深呈负相关。同时,尚存在其他因素能影响海草生长与分布状况。
3.5.3 陆源影响分析
为进一步查明马山里海域海草生长状况差异的原因,对马山里海域周边环境进行分析。马山里海域通过涨潮、退潮与天鹅湖和养鱼池湾两处海域进行频繁水体交换。因此,同时对马山里海域、天鹅湖和养鱼池湾周边陆地环境进行调查,将自然保护区、耕地、居民区和工厂(养殖场)按污染强弱分别赋分,赋分标准见表3。
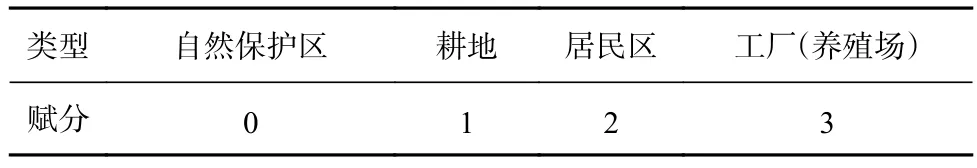
表3 环境类型及赋分标准Table 3 Environment type and assignment standard
对赋分结果采用克里金插值法分析,如图11所示。结果发现,马山里海域周边以自然保护区为主,赋分最小,即受到的陆源污染最少;其次天鹅湖周边以自然保护区和居民区为主,赋分居中,受到的陆源污染居中;养鱼池湾周边工厂(养殖场)数量众多,赋分最高,受到的陆源污染最多。赋分结果与马山里海域海草生长状况评估结果(图6)相一致,故马山里海域海草生长状况的差异可能与陆源污染程度有关,南北两端来自于天鹅湖、养鱼池湾的陆源污染物较多,海草生长状况较差,马山里中部海域陆源污染较少,海草生长状况较好。
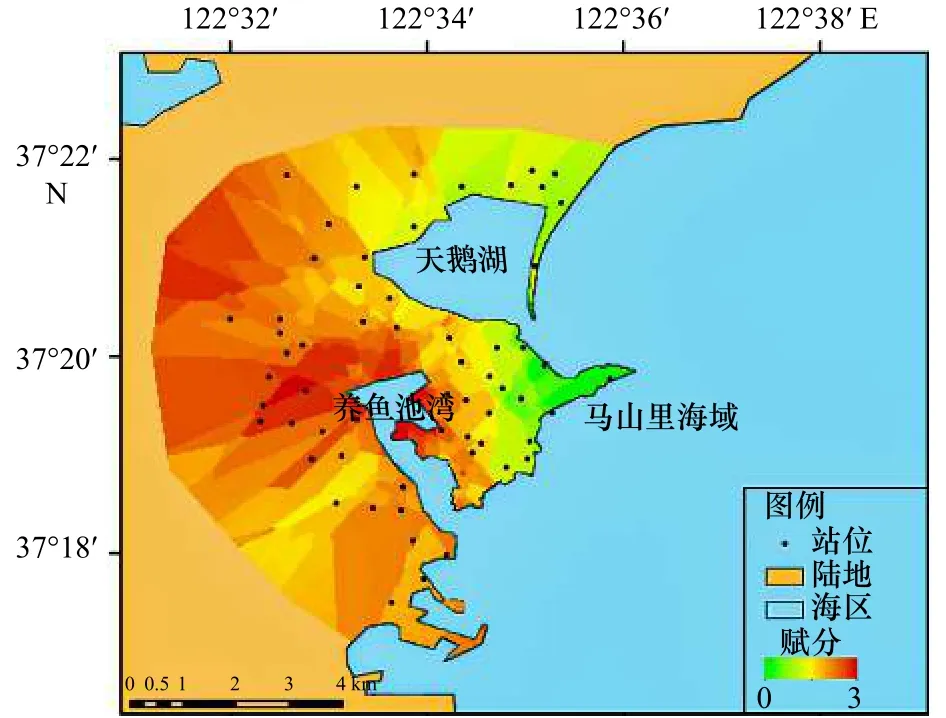
图11 环境类型赋分结果示意图Fig.11 Schematic diagram of environment type assignment results
4 讨论与建议
4.1 海草床的分布及资源量现状
马山里海域有红纤维虾形草和丛生鳗草两种海草,种类的分布与底质类型有直接关系。红纤维虾形草附着生长在岩石或礁石上,受底质条件限制,大多呈现带状分布[32-33],而丛生鳗草偏好细沙或黏土含量较高的沉积环境,如黏土质砂等[34-35]。类似地,马山里海域红纤维虾形草受海区礁石分布影响从而呈现南北两个狭长的带状分布区域;马山里海域由于黏土质砂底质的不连续分布,丛生鳗草难以形成连续草床,从而呈现零星斑块状分布。
在2011年世界自然保护联盟(IUCN)濒危物种红色名单灭绝风险的分类与评估标准中,红纤维虾形草和丛生鳗草均被列为易危种[5]。马山里海域属外海海域,自然条件恶劣,加之近年来人为活动的干扰日渐加剧,海草面临一定程度的退化风险。由于缺乏马山里海域海草床的历史调查数据,无法对其进行历史对比。但据同期调查资料显示,马山里海域红纤维虾形草的平均茎枝密度和生物量均低于周边镆铘岛、桑沟湾和双岛湾海域,其现存植株平均数量(分布面积与平均茎枝密度的乘积)仅为镆铘岛的7%,桑沟湾的18%,如表4所示。Yabe等[36]调查显示,夏季(7月)日本千叶县沿海红纤维虾形草的平均茎枝密度和生物量高达1 809 shoots/m2和2 226 g/m2,分别是相同季节(8月)马山里海域红纤维虾形草的4.9倍和6倍。这些对比结果表明,马山里海域红纤维虾形草的资源量较低,海草床生态系统处于脆弱状态,存在相当程度的退化风险。

表4 荣成近岸红纤维虾形草资源量Table 4 Resources of Phyllospadix iwatensis in the Rongcheng coastal
马山里海域丛生鳗草海草床平均茎枝密度((691.2±17.1)shoots/m2)和生物量((534.0±70.7)g/m2)略大于桑沟湾的丛生鳗草海草床((513.0±17.9)shoots/m2、(479.1±17.9)g/m2)[11],但马山里海域丛生鳗草零星分布,总面积为3.76 hm2,仅为后者的一半。因此,马山里海域丛生鳗草面临的主要风险是生境破碎化,这些海草斑块不仅会对海草床的生物多样性[37-38]、生态系统功能[39]产生负面影响,而且抵御环境变化和人为扰动的能力较差,极易因受到自然环境和人类活动的影响而消失,急需加强生境保护与修复[6,40-41]。
4.2 关键环境因子对海草床分布与生长的影响
海草对环境变化敏感,其生长发育受多种环境因子影响。典型相关分析表明,马山里海域海草生态学指标整体变化由水温和水深等环境因子所主导,与水温呈正相关,与水深呈负相关。
红纤维虾形草和丛生鳗草均属于温带海草,最优生长温度为11.5~20℃[42],典型相关分析结果显示,海草生态学指标与水温呈正相关,因此水温较高区域,海草生长状况较好,茎枝密度和单株生物量均较高。Yabe等[36]在日本铫子市对海岸海草生长状况跟踪调查显示,水温升高能够提高红纤维虾形草的生产力,与本文调查结果类似。
水深主要通过影响到达海草冠层的光照强度控制海草的分布与生长。太阳光进入海水后因吸收和散射而快速衰减,而光照强度则会影响海草叶片的叶绿素a、叶绿素b、类胡萝卜素、可溶性蛋白、非结构性碳水化合物等含量,是海草光合作用的决定性因素[43-44]。水深越浅,底层光照越充足,越有利于海草光合作用,因而能够支持更高的茎枝密度和生物量[43]。马山里海域红纤维虾形草海草床分布水域平均水深为2.6 m,丛生鳗草海草床分布水域平均水深为3.0 m,数值均为低潮时测得,当地潮差约为1.5 m,符合海草的适宜生长水深(6 m以浅)[44],而两者分布水深的差异可能是由于两者对光照的适宜生长条件不同导致。
4.3 人类活动对海草床分布与生长的影响
结合走访调查与环境赋分分析的结果发现,人为因素是导致马山里海域海草床退化的主要原因,主要包括渔业生产、养殖活动和陆源污染的排放。
渔业生产方面,马山里海域曾有相当面积的填海工程,且建有码头。填海工程会直接掩埋海草,侵占其生长海域;码头建设施工过程会造成水体悬浮物增加,引起光照衰减,影响海草的光合速率[28];船舶航行对海草床进行反复切割[45-46],会导致海草床的斑块化和生境破碎化;拖网、吸贝等生产方式则在短时间内造成海草的大面积死亡[28]。海南岛南部铁炉港海域,由于破坏性的挖贝方式,2008−2014年间,海菖蒲(Enhalus acoroides)海草床面积由1.18 hm2降至0.04 hm2,且呈现零星分布,几近消失[27]。
养殖活动方面,马山里海域养殖场数量众多,主要包括贝类养殖、海珍品养殖和海带浮筏养殖。养殖活动主要从两方面影响海草的生存生长:一方面,开掘池塘、围海养殖等挤占海草生存空间;另一方面,养殖活动对水体干扰较大,使海草失去适宜的生存环境。20世纪90年代初,在广东省流沙湾海域海草床中开挖虾池,目前该范围内海草床已完全退化[24];曾在广西合浦海域海草床大规模吊养贝类,截至目前已有100 hm2海草床消失[12]。
陆源污染方面,马山里海域直接输入的陆源污染物较少,但受到来自养鱼池湾陆源污染的间接影响,同样存在污染压力。随着污染的加重,海水出现富营养化状态,直接抑制海草叶片叶绿体的磷酸化过程,对海草产生毒性作用,进而影响海草的茎枝密度和形态学特征[47-49],且植株铵态氮的过量富集会引起植株中毒,导致生长受损[50]。此外,海水富营养化会促使大型藻类、叶片附生藻类和浮游植物大量暴发,遮挡阳光,抑制海草光合作用,影响其生长[51-52]。另外,陆源污染中的除草剂、重金属等有毒物质进入海域,会对海草产生长期毒害作用[53-54]。
4.4 保护建议
马山里海域海草资源量较少,加之底质坚硬、风浪较大,海草种子定栖和植株扎根的难度大,一旦海草床遭到严重破环,自然恢复的可能性较低。因此,急需开展海草床的保护与修复工作。针对马山里海域海草床所受威胁的类型和强度,提出以下建议:
(1) 加强马山里海域海草床的保护。特别是合理规划海带养殖规模,降低养殖密度;严控周边海域陆源污染物的排放;提高政府重视程度,科学规划海岸建设等活动,减少对海草床的破坏。
(2) 开展马山里海域海草床生态修复工程。海草床自然恢复周期漫长,应当辅以必要的人工恢复措施。近年来,海草的修复研究已取得一定成果[55-56],但主要适合生长在泥沙底质的鳗草等种类,对于红纤维虾形草和丛生鳗草并不适用。截至目前,仅程冉等[57]利用构件开展了红纤维虾形草植株的移植,但过程复杂,劳动量和成本较高,无法推广应用,而丛生鳗草的修复技术更为鲜见。因此,应当进一步加强红纤维虾形草和丛生鳗草修复技术的研发,科学有效开展海草床的修复工作。
(3) 加强红纤维虾形草和丛生鳗草的基础研究。尤其是两者的生活史、生长发育以及对环境胁迫的响应机制研究,从而为两种海草的保护和修复提供数据基础和理论基础。
