不同土地利用方式土壤氨氧化微生物和反硝化微生物时空分布特征
2022-08-15蔡玉佳沈菊培张成军冯虞彦DIHongjie贺纪正
蔡玉佳,沈菊培,张成军,冯虞彦, DI Hongjie,贺纪正
1 福建师范大学,地理科学学院,福州 350007 2 西交利物浦大学,健康和环境科学系,苏州 215123 3 北京农林科学院,植物营养与资源研究所,北京 100097 4 林业大学,环境科学与工程学院,北京 100083 5 新西兰林肯大学, 土壤与环境研究中心, Canterbury 7674
氮是植物生长的重要营养元素之一,对农业生态系统具有重要作用。硝化和反硝化过程是氮循环主要过程,均为微生物所驱动,两者共同调节植物和微生物可利用氮含量,也是土壤N2O产生的主要途径[1]。氮循环过程受多种因素的影响,包括气候,土壤类型,施肥和土地利用方式等[2—3]。土地利用方式通过改变土壤理化性质和养分有效性等影响地上地下生态系统功能。Shen等[4]研究了酸性红壤在5种不同土地利用方式下古菌和氨氧化微生物,发现土地利用方式显著影响古菌群落结构,且氨氧化古菌(Ammonia oxidizing archaea, AOA)对土地利用方式变化的响应比氨氧化细菌(Ammonia oxidizing bacteria, AOB)敏感。Liu等[5]研究发现灌区撂荒地和农耕地表层和底层土壤氮转化速率和氨氧化微生物数量存在显著差异,农耕地土壤的AOA和AOB数量显著高于撂荒地。Zulkarnaen等[6]研究了我国亚热带红壤地区林地和旱地农田土壤中的硝化作用,结果表明旱地土壤AOA和AOB数量和净消化速率高于林地土壤,但二者的N2O排放速率没有显著差异。可见,土地利用方式的改变对土壤硝化过程的影响还没有统一的定论,特别对驱动氨氧化和反硝化过程的微生物群落的季节影响还有待深入[4]。本文主要对比分析农田和林地两种土地利用方式下氨氧化微生物和反硝化微生物的季节变化特征,可为预测土壤硝酸盐淋溶和环境氮损失提供参考。
土壤是异质性较高的体系,其生物和化学性质在不同剖面上存在显著差异。深层土壤氧含量较低,温度变化小,土壤溶液或水分停留时间较长[7]。在灌溉和降水的水力驱动下,硝酸盐和活性溶解性有机碳组分将向深层土壤迁移,改变深层土壤养分组成[8—9],进而影响土壤微生物群落。由于表层土壤的微生物生物量大,活性高,当前对其生物地球化学过程的研究给予了极高的关注。有研究表明深层土壤储存着大量的有机碳,且具有相当丰富和多样的土壤微生物。40—200 cm土壤的微生物量可占200 cm土壤剖面微生物全量的35%,这些深层土壤微生物在土壤成土过程、污染物降解和维持地下水水质等生态功能方面具有重要作用[10]。因此,研究不同土地利用方式下硝化、反硝化微生物沿土壤剖面的动态变化对认识硝酸盐的垂直淋溶过程具有重要的实践指导意义。
目前对不同土地利用方式下土壤氮转化过程及微生物机制已有不少研究,但对土壤硝化和反硝化微生物的时空分布特征的认识还不够。本文通过采集华北平原典型潮土两种土地利用方式0—100 cm的剖面样品,研究玉米地和林地土壤氨氧化微生物和反硝化微生物丰度垂直分布特征,并探讨其与剖面土壤硝酸盐的关系,旨在为调控和减少硝酸盐淋溶风险、降低土壤N2O排放提供重要的理论指导。
1 材料与方法
1.1 样品采集
采样地点位于北京市延庆区某奶牛养殖场附近的玉米地和邻近的次生林地(40°29′N,115° 56′E),土壤类型均为粉砂质粘壤土(美国制),容重为1.31 g/cm3。该地区属于典型大陆性季风气候,年平均温度为8℃,年平均降水量为400—500 mm,降水集中在7—9月(夏季),占全年降水量的70%[11]。样地附近某奶牛养殖场已经营十年,圈养奶牛200多头,每年所产牛粪(约4000 t)作为基肥施用于临近玉米地。本研究选取了该处理下大小一致的三块玉米地(200 m×200 m),同时采集了邻近未施用牛粪的次生林林地土壤。该次生林在2004年实施退耕还林之前的土地利用方式和土壤母质与玉米地一致。采样时间为2018年春季玉米种植之初(5月初)和秋季收割之后(9月底)。
采用“S”型采样法在每块地选取5个采样点,用管型土钻采集0—20、20—40、40—60、60—80、80—100 cm深度的土壤,每层5个采样点分别混合均匀为一个土壤样品,共采集得到60个土壤样品(即2个土地利用方式×3个重复×5个土层×2个季节)。去除土壤样品中的植物残体和碎石后于采集当天送至实验室,过2 mm筛后分为两部分:一部分存于4℃冰箱用于土壤氨氧化潜势和亚硝酸盐氧化潜势测定和后续土壤理化分析;另一部分存于-40℃冰箱,用于提取土壤DNA。
1.2 土壤基本性质测定
土壤pH使用pH计(METLER TOLEDO FE20)以土水比1∶1测定。土壤有机质测定采用低温外热重铬酸钾氧化-比色法。利用元素分析仪(Vario EL III, Elementar)上测定土壤总氮含量。土壤溶解性有机碳和溶解性有机氮含量的测定过程如下:称取5.0 g鲜土用0.5 mol/L K2SO4溶液以1∶5的土水比浸提,离心过滤后,在总有机碳/总氮分析仪(Multi N/C Model 3100, Analytic Jena, Germany)上测定。土壤铵态氮和硝态氮用1 mmol/L KCl溶液以1∶5的土水比浸提,离心过滤后,流动分析仪(AA3, SEAL, Germany)测定。土壤基本性质见表1。

表1 春秋两季玉米地和林地土壤剖面基本性质
1.3 土壤氨氧化潜势和亚硝酸盐氧化潜势测定
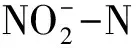
1.4 土壤DNA提取
称取0.50 g冷冻样品,利用土壤DNA试剂盒(DNeasy Power Soil Kit, QIAGEN, Germany)根据说明书提取土壤DNA并在NanoDrop ND-1000分光光度计(NanoDrop, Thermo Scientific, USA)上测定DNA浓度和纯度,随后保存于-40 ℃下用于后续分析。
1.5 土壤细菌和氮功能基因丰度测定
通过实时荧光定量PCR方法,在LightCycler 480II(Roche, Switerland)上测定细菌16S rRNA基因、AOAamoA、AOBamoA、反硝化nirS、nirK和nosZI基因的丰度。细菌16S rRNA基因丰度采用TaqMan探针法,氮相关功能基因丰度的测定采用嵌合荧光法检测法。PCR扩增体系为20 μL,内含10.0 μL Premix Ex Taq II(分别为PrimeDirect Probe RT-qPCR Mix, TAKARA, Japan和SYBR Green Supermix, BIO-RAD, USA),0.5 μL或1.0 μL前引物和后引物,1.5 μL或2.0 μL DNA模板,用超纯水补足(RNase-free)。定量PCR引物序列信息、PCR扩增程序及标线制作见袁超磊等[14]。定量PCR标线的扩增效率在90%—100%范围,标准曲线R2>0.99并通过凝胶电泳检验PCR产物。
1.6 数据分析
采用SPSS 16.0对数据进行单因子、多因子方差分析。采用R语言“vegan”,“Hmisc”,“corrplot”包等对硝化活性、氮功能基因丰度与土壤理化性质之间的关系进行冗余分析(Redundancy Analysis, RDA)和Pearson相关分析。冗余分析中膨胀因子大于10的环境因子没有呈现。采用SigmaPlot 10.0和R的“corrplot”包作图,P<0.05差异检验显著。
2 结果与分析
2.1 玉米地和林地土壤氨氧化潜势和亚硝酸盐氧化潜势时空分布特征
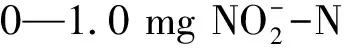
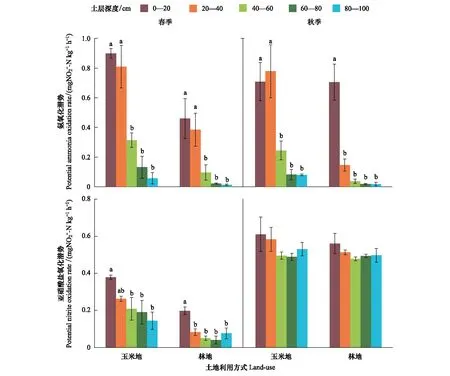
图1 春、秋两季玉米地和林地不同土层的氨氧化潜势和亚硝酸盐氧化潜势
2.2 玉米地和林地土壤细菌和氮循环功能基因丰度时空分布特征
图2为不同土地利用方式下细菌16S rRNA基因、氨氧化微生物(AOAamoA、AOBaomA)和反硝化微生物(nirS、nirK和nosZI基因)相关功能基因的垂直分布特征。土壤细菌和氮循环功能基因丰度均随土壤深度增加而显著降低(P<0.05)(图2、表2)。玉米地沿土壤剖面各氮循环功能基因丰度范围(拷贝数/g 干土)分别为AOAamoA基因 5.22×107—3.30×103,AOBamoA基因 5.62×107—1.0×103,nirS基因4.32×107—9.91×102,nirK基因4.62×107—2.97×104和nosZI基因6.25×107—2.70×103。林地土壤沿剖面则分别为AOAamoA基因7.67×107—1.39×103,AOBamoA基因5.52×106—1.0×103,nirS基因4.03×106—1.02×103,nirK基因5.49×106—2.69×104和nosZI基因4.75×106—3.29×103。整体上玉米地土壤氨氧化、反硝化微生物数量高于林地土壤。
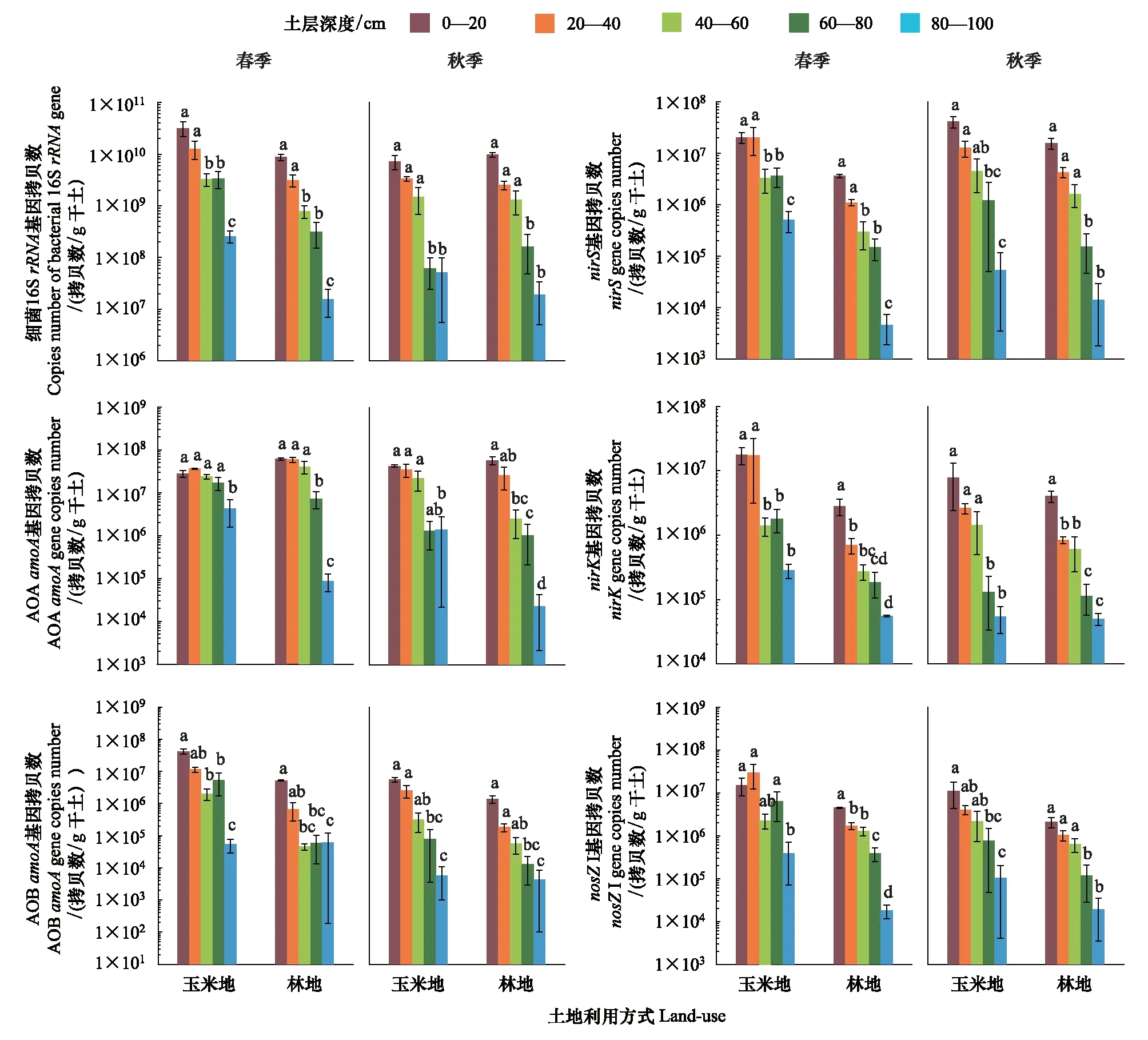
图2 玉米地和林地土壤细菌16S rRNA基因和氮循环功能基因丰度时空分布
单因子方差分析结果表明春季玉米地土壤AOBamoA、nirS、nirK和nosZI基因丰度显著高于林地(P<0.01),而秋季样品两种利用方式间无显著差异。玉米地AOBamoA(F=6.905,P<0.05)、nirS(F=9.791,P<0.01)、nirK(F=4.577,P<0.05)和nosZI(F=4.857,P<0.05)基因丰度在不同季节间差异显著,且秋季氨氧化、反硝化微生物的丰度呈下降趋势,而AOAamoA基因(F=3.017,P>0.05)在不同季节间无显著差异。另外,季节变化对林地土壤氮循环功能基因没有显著影响(P>0.05)。多因子方差分析表明土地利用方式、季节变化和土壤深度对细菌丰度和氮循环功能基因有显著影响,但各变量间的交互作用均不显著(表2)。
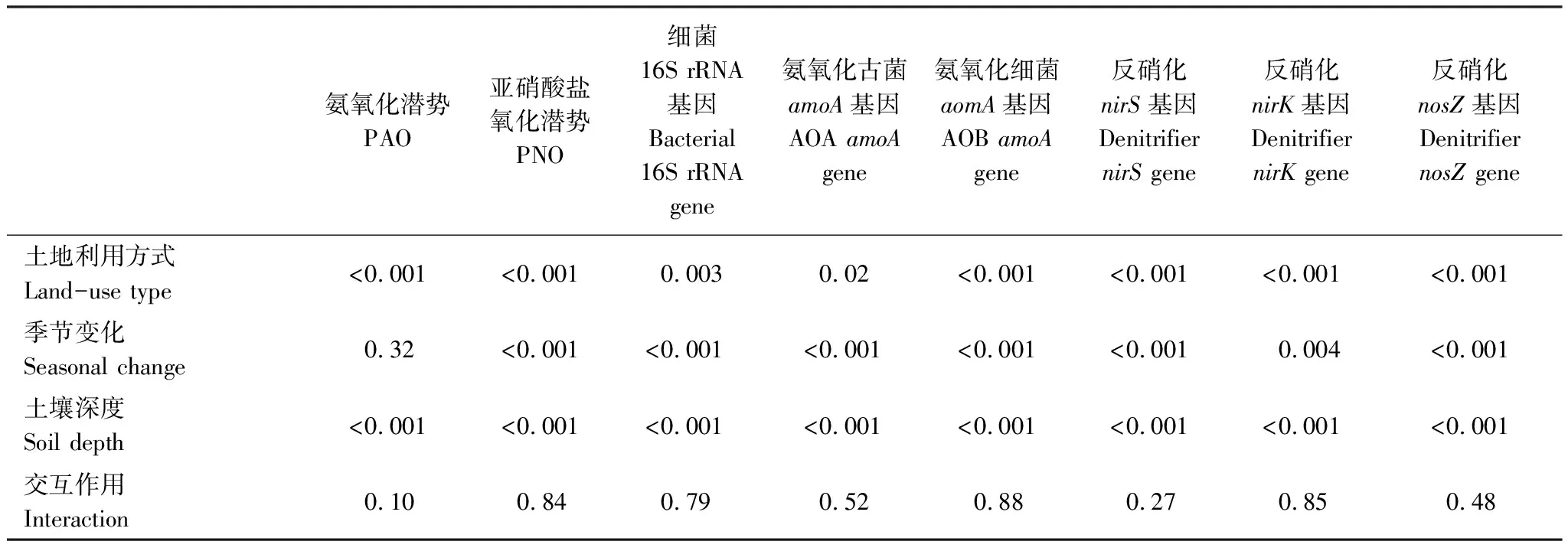
表2 三因子PerMANOVA分析土地利用方式、季节变化、土壤深度及其交互作用对氨、亚硝酸盐氧化潜势和氮循环基因的影响
2.3 土壤理化因子与氮功能微生物基因丰度和功能的相关性
图3为氨氧化、反硝化微生物丰度与土壤理化性质之间的冗余分析结果。土壤理化性质分别解释了玉米地和林地氮循环微生物差异的48.7%和72.7%。玉米地的第1和第2排序轴解释量分别为44.2%和3.5%,而林地的第1和第2排序轴的解释量分别为69.9%和1.7%。如图3所示,玉米地和林地土壤可溶性有机碳、可溶性有机氮、铵态氮、硝态氮与氨氧化、反硝化微生物丰度正相关,而土壤C/N比和pH与氨氧化、反硝化功能基因丰度显著负相关。
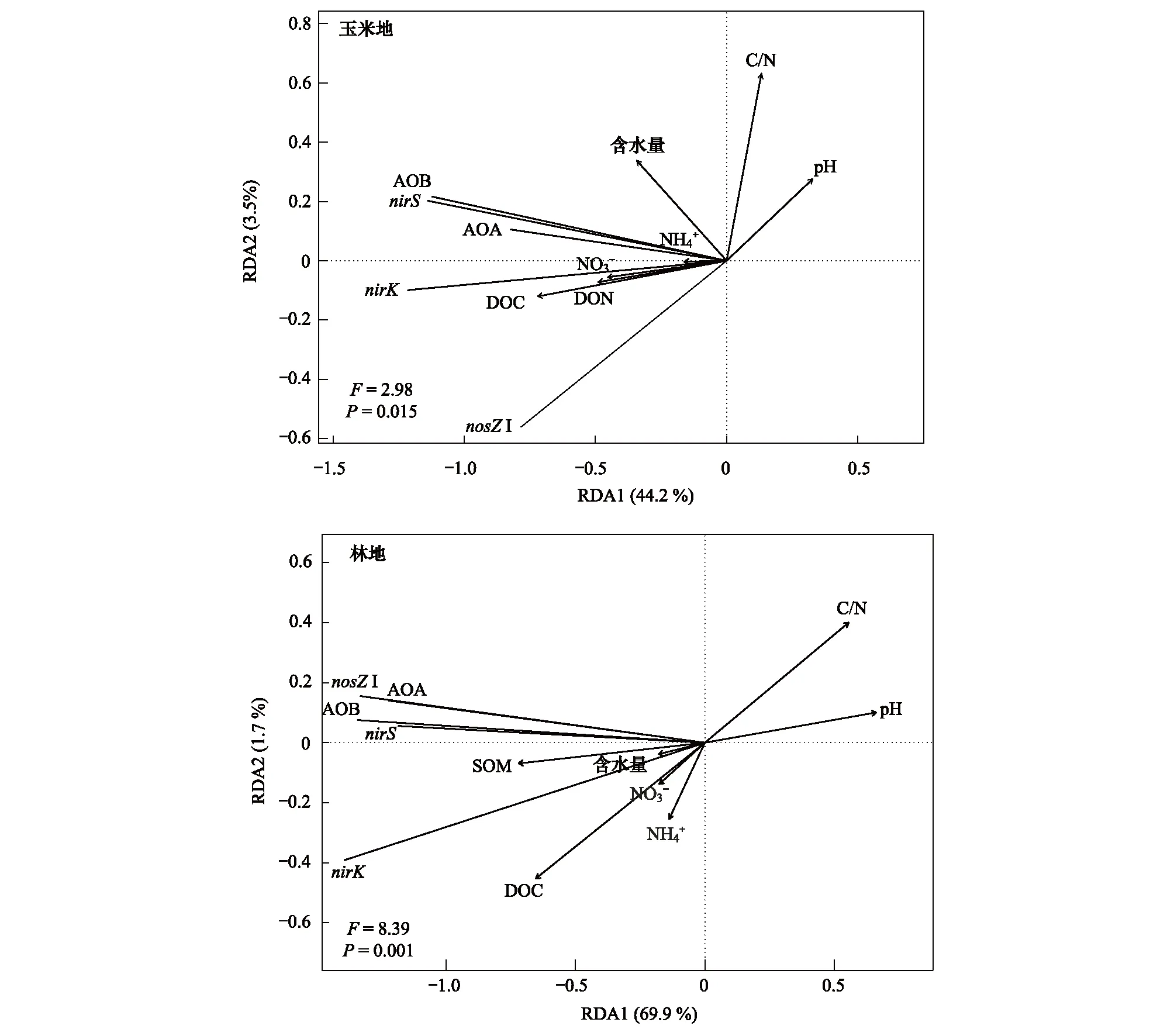
图3 玉米地和林地氨氧化、反硝化基因丰度和土壤理化性质的冗余分析(RDA)
Pearson相关性分析(图4)表明土壤有机质、总氮、可溶性有机碳和可溶性有机氮含量与氨氧化、反硝化基因丰度、氨氧化潜势和亚硝酸盐氧化潜势显著正相关,而土壤C/N比、pH与氨氧化、反硝化基因丰度、氨氧化潜势和亚硝酸盐氧化潜势显著负相关。玉米地春季样品土壤铵态氮、硝态氮含量与氨氧化潜势显著正相关,硝态氮含量还与AOBamoA、nirS和nirK基因丰度呈显著正相关关系。林地土壤春季样品铵态氮、硝态氮含量与氨氧化、反硝化基因丰度相关性却不显著。另外,玉米地秋季样品铵态氮含量与AOBamoA、nirS基因拷贝数呈显著正相关,而硝态氮含量与各功能基因丰度相关性均不显著;而林地土壤硝态氮含量与氮循环功能基因丰度呈显著正相关,相关系数大于0.5。

图4 春、秋两季玉米地和林地土壤理化性质与微生物丰度相关性分析
3 讨论
土地利用方式的改变会引起土壤理化性质的变化,从而影响土壤微生物资源可利用性,最终改变土壤微生物包括病毒、细菌、古菌和真菌群落多样性和结构[4,15—16]。本研究结果表明玉米地土壤养分含量、细菌和氮循环相关微生物的丰度显著高于次生林土壤,这与前人的研究结果不一致。有研究发现天然林地转变为人工林、耕地等方式后,土壤剖面有机碳储量降低[17],而退耕还林,由于减少人为干扰后土壤有机碳含量增加[18]。Templer等[3]发现原始林地、农田及再生林土壤的氮转化过程存在差异,即使再生林土壤的理化性质已与原始森林土壤接近,但其土壤微生物量氮、土壤呼吸速率、矿化和硝化速率均低于原始林地。引起结果差异的主要原因是农田土壤受外源物质输入的影响。本文中玉米地的养分含量显著高于次生林地主要长期受牛粪的影响,而林地土壤除大气氮沉降外,无外源物质的输入。牛粪等畜禽粪便作为一种良好的有机肥,含有丰富的易降解的有机碳、氮、磷和其它微量元素,如果管理和使用得当,极利于改善土壤肥力和健康,提高土壤微生物生物量[19]。以往长期施肥试验研究结果表明,有机粪肥的施用可改善土壤养分条件,促进土壤微生物和氮循环微生物的生长,进而影响土壤氮转化过程[20—21]。
通常认为氨氧化过程是硝化过程的限速步骤,然而在土壤环境受到干扰时亚硝酸盐氧化过程也可成为硝化过程的限速步骤[31]。本研究测定了两种土地利用方式下土壤氨氧化潜势和亚硝酸盐氧化潜势,发现玉米地土壤氨氧化潜势和亚硝酸盐氧化潜势显著高于林地土壤,这与土壤氨氧化微生物丰度变化趋势一致。相关性分析结果表明AOAamoA、AOBamoA基因丰度与氨氧化、亚硝酸盐氧化潜势之间存在显著相关性(图4)。蔡祖聪和赵维[32]观测了不同土地利用方式对湿润亚热带地区土壤硝化作用的影响,发现农用地的硝化潜势高于自然土壤(森林和灌丛)。也有研究结果表明有机肥施用可显著提高土壤氨氧化潜势和亚硝酸盐氧化潜势[22, 33—34]。因此,不同施肥管理方式可引起氨氧化微生物发生变化,进而影响土壤氨氧化、亚硝酸盐氧化过程。
硝化过程为反硝化过程提供底物,因此,硝化和反硝化过程通常耦合发生,共同作用并构成土壤氮素损失的主要途径[3]。本文发现两种土地利用模式下土壤硝化与反硝化微生物数量间均存在显著的正相关关系。前人用Meta分析结果也表明硝化、反硝化微生物紧密相关,且其丰度分别与硝化、反硝化速率之间存在显著的相关性[42],说明硝化、反硝化微生物的数量可间接反映硝化、反硝化速率。本研究中,玉米地土壤反硝化nirS、nirK和nosZI基因的拷贝数均显著高于林地土壤。玉米地春季土壤各剖面层的nirS、nirK和nosZI基因丰度比对应的林地剖面土壤高1—2个数量级。在土壤剖面上,nirS、nirK和nosZI基因丰度均随土壤深度增加而显著下降。除林地秋季样品外,各土层的nirS基因丰度均高于nirK基因,表明nirS型亚硝酸盐还原微生物在亚硝酸盐还原过程中起主导作用。此结果与Yuan等[43]报道的水稻土中nirS型反硝化微生物较nirK型具有更高的反应活性的结果较一致。一般认为潮土的砂粒含量较高,土壤有机碳含量较其它土壤类型低,通气性好,不利于反硝化过程的发生。但是在深层土壤中,由于硝酸盐的淋溶,底物充足,土壤氧含量低等因素,有利于反硝化过程的进行[42, 44]。RDA分析结果也表明土壤理化性质和反硝化基因存在明显的关系,土壤有机质含量、溶解性有机碳、氮含量、铵态氮和硝态氮含量可解释多数的变化丰富的反硝化功能基因。由于反硝化过程是一个耗能过程,微生物需要消耗碳源,因此土壤高有机碳含量可促进反硝化微生物的生长[45]。反硝化功能基因(nirS+nirK)/nosZ的丰度比值可反映反硝化过程N2O的排放情况。本研究中,玉米地土壤的(nirS+nirK)/nosZI比值在80—100 cm达到最大值(5.5),N2O排放潜势高,但由于N2O向上迁移的过程中,可被表层反硝化微生物截留、转化为N2。因此,推测深层土壤具有较高的反硝化潜力,可用于考虑筛选反硝化微生物的富集培养土壤,而如何平衡深层土壤反硝化微生物活性及硝酸盐淋溶则是今后地下水污染的重要课题之一[46—47]。
4 结论
本文结果表明:不同土地利用方式下,土壤硝化和反硝化微生物在季节更替和土壤垂直方向上存在显著差异,而不同氮转化微生物对季节变化的相应则不同。玉米地春季农田土壤氨氧化、反硝化微生物丰度均显著高于林地,氨氧化潜势也均高于林地。土壤碳、氮含量,硝化、反硝化微生物丰度及亚硝酸盐氧化潜势随土壤深度增加而显著下降。可见,如何提高土壤保氮能力、防控硝态氮淋溶和N2O排放仍是今后农田氮肥管理和生态环境保护的研究重点。
