贵州老鹰茶基于叶绿体DNA的遗传多样性分析
2022-08-15万诚李蕾嘉陈胜王济红王莹
万诚 李蕾嘉 陈胜 王济红 王莹
(贵州省生物研究所,贵州 贵阳 550009)
老鹰茶具消暑解渴、消食去胀、降血糖、降血脂、抑菌、消炎、抗氧化等独特的功效,在西南地区,人们把老鹰茶作为盛夏的民间饮品及民间草药[1]。国内学者在生物活性成分和营养成分上的研究越来越深入,近30a来一直是开发利用研究的热点[2]。老鹰茶基源植物的分类归属不明确,10a前报道的3属2种2变种:木姜子属朝鲜木姜子(Litsea coreana)的变种豹皮樟(L.coreana var.sinensis)和毛豹皮樟(L.coreana var.lanuginose);润楠属川黔润楠(Machilus chuanchienensis)和黄肉楠属红果黄肉楠(Actinodaphne cupularis)[3,4];近年来被报道为4属5种1变种:豹皮樟、川黔润楠、红果黄肉楠、木姜子(Litsea pungens)、狭叶润楠(Machilus rehderi)和山胡椒属香叶树(Lindera communis)[5]。关于老鹰茶植物的叶绿体DNA遗传多样性分析鲜有报道,Feng等[6]对采自中国7省区的23份老鹰茶植物进行了trnH-psbA、trnL-trnF和trnT-trnL 3个叶绿体标记的测序,鉴定出8个单倍型,并将23份样品聚为4个类群,认为老鹰茶植物基源为豹皮樟,但未将表观性状与DNA测序结果进行分析,不能为品种选育提供依据。Pan等[7]在Illumina MiSeq平台上,对采自正安的豹皮樟变种的叶绿体DNA全基因组进行测序,并与29种樟科植物叶绿体基因组构建了系统发育树。本研究拟通过对老鹰茶原料植物DNA叶绿体分子标记分析其遗传多样性,研究结果将为老鹰茶植物的分类归属提供科学依据,为进一步开展良种选育与利用奠定理论基础。
1 材料与方法
1.1 试验材料
老鹰茶植物分化为嫩芽被白色、被黄色和被粉红色绒毛3种芽表现型的现象,目前根据开花、结实情况判断,被白色绒毛芽表现型应该为雄株表征,被红色绒毛芽表现型应该为雌株表征,被黄色绒毛芽表现型应该为子代杂合体表征。
老鹰茶古树样品收集于贵州省遵义市湄潭县境内,针对红芽雌株、黄芽雌株、黄芽雄株和白芽雄株4种性别表型共10株进行采样,材料收集好后放入-80℃冰箱备用,样本详细信息见表1。
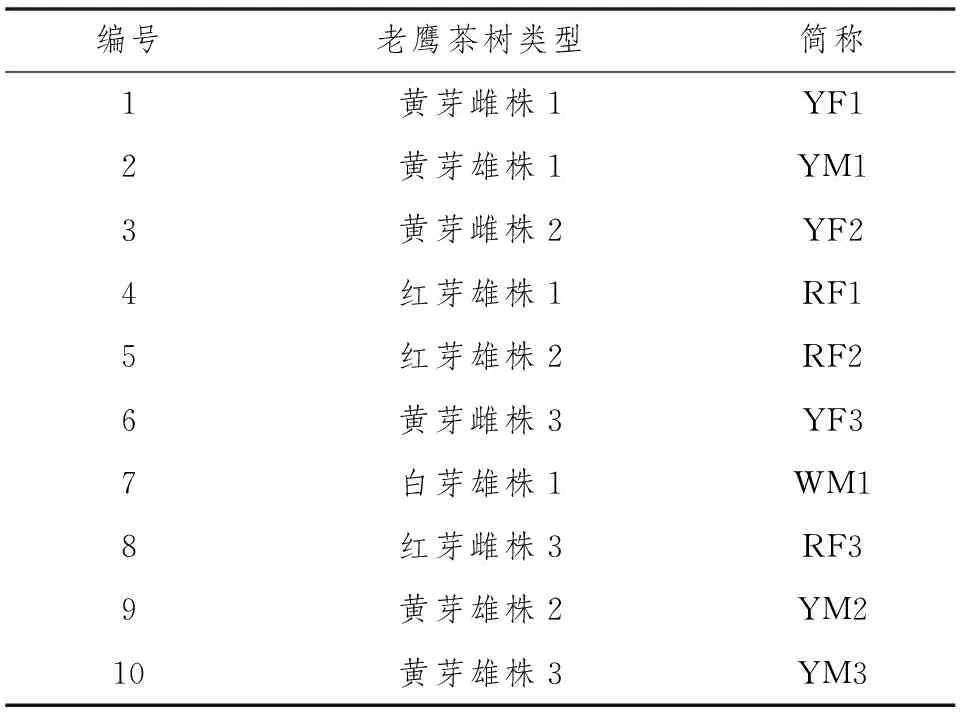
表1 供试老鹰茶树信息
1.2 DNA提取
本试验使用试剂盒法,选用上海生物工程股份有限公司的植物基因组DNA快速抽提试剂盒。
1.3 叶绿体DNA引物筛选
参考中华人民共和国林业行业标准LY/T 3191-2020 林木DNA条形码构建技术规程[8],选出标记叶绿体基因的H-psbA与L-F间区进行PCR扩增,扩增序列见表2。

表2 所选叶绿体间区引物序列
1.4 PCR扩增体系及电泳
PCR扩增体系选用20μL体系:2×SanTaq PCR Mix 10μL、无菌水7μL、引物2μL、DNA1μL。PCR程序为94℃预变性4min,94℃变性30s,以低于引物溶解温度5℃复性45s,72℃延伸2min,共35个循环;72℃延伸7min,4℃保存。
采用1%浓度琼脂糖凝胶检测已扩增好的目标片段,若目标条带清晰,不拖尾,无杂带干扰,说明扩增得到的是所需要的目标产物。
1.5 测序
将扩增得到PCR产物送至上海生物工程股份有限公司进行测序。
1.6 数据分析
将上海生物工程股份有限公司反馈的叶绿体DNA序列进行处理。同源性对比在美国国家生物信息网站中NCBI的BLAST中进行;使用MEGA-X[9]软件对2个叶绿体DNA间区正反向序列的拼接、比对与合并,构建系统发育树;将处理好的序列在美国国家生物信息网站NCBI中的BLAST中进行同源性对比;使用DnaSPver 6.0软件[10]计算10份老鹰茶植物叶绿体DNA的遗传多样性参数,包括多态性变异位点(Vs)、单一突变位点(Ss)、插入-缺失位点(Is)、简约信息位点(Ps)、核苷酸多样性(Pi)、平均核苷酸差异(k)、单倍型数(h)、单倍型多样性(Hd)、单倍型多样性方差(Vh)、单倍型多样性标准差(Sh)与Tajima’s D值,Tajima’s D是检验供试材料叶绿体DNA区域是否遵循中性进化模型的重要参数。
2 结果
2.1 PCR产物电泳检测
PCR扩增后检测结果见图1,2对叶绿体DNA扩增引物均扩增出清晰、单一明亮的目标条带,均为试验目标片段。
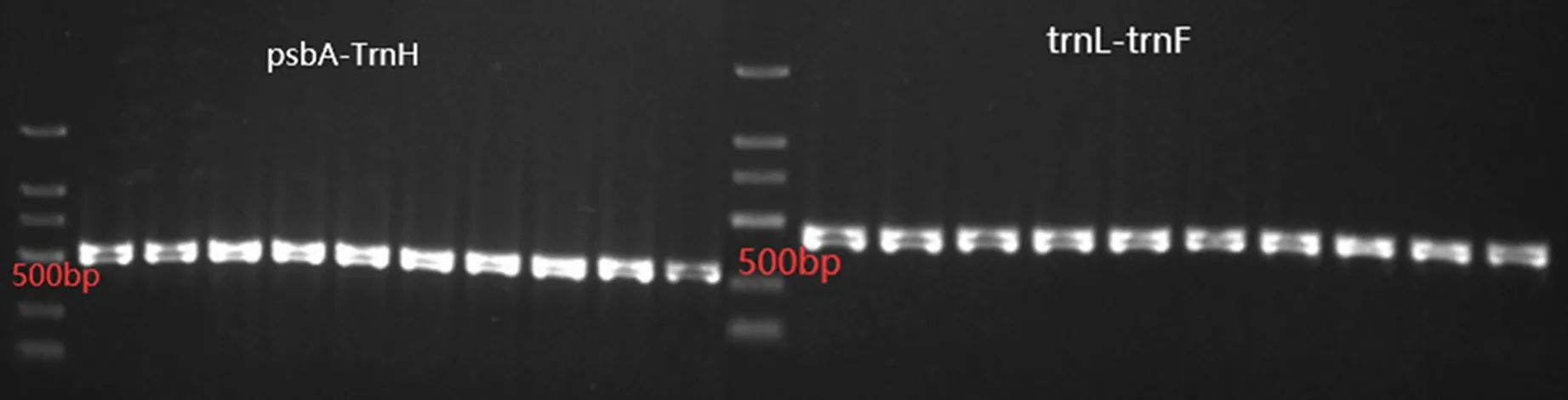
图1 2对叶绿体DNA引物扩增检测结果
2.2 老鹰茶植物同源性比对
将拼接好的序列导入美国国家生物信息网站NCBI中的BLAST进行比对,比对后发现与GenBank中的Actinodaphne lancifolia(登录号为NC_045251.1)的朝鲜木姜子种的相似值达100%。
2.3 老鹰茶植物叶绿体DNA多态性分析
通过2对叶绿体DNA通用引物对10份贵州省遵义市湄潭县采集的老鹰茶植物的2个叶绿体DNA间区进行扩增,结果见表3、表4,2个区域H-psbA与L-F在所有样本中的序列长度分别为530bp和446bp,其中,核苷酸多态性较高的区域为H-psbA,包含28个单一突变位点、11个简约信息位点和5个插入-缺失位点;核苷酸多态性比较低的区域为L-F,仅含2个简约信息位点和6个插入-缺失位点,没有单一突变位点。2个叶绿体DNA间区序列组合之后序列长度为976bp,含有28个单一突变位点、13个简约信息位点和11个插入-缺失位点,共计41个多态性变异位点。
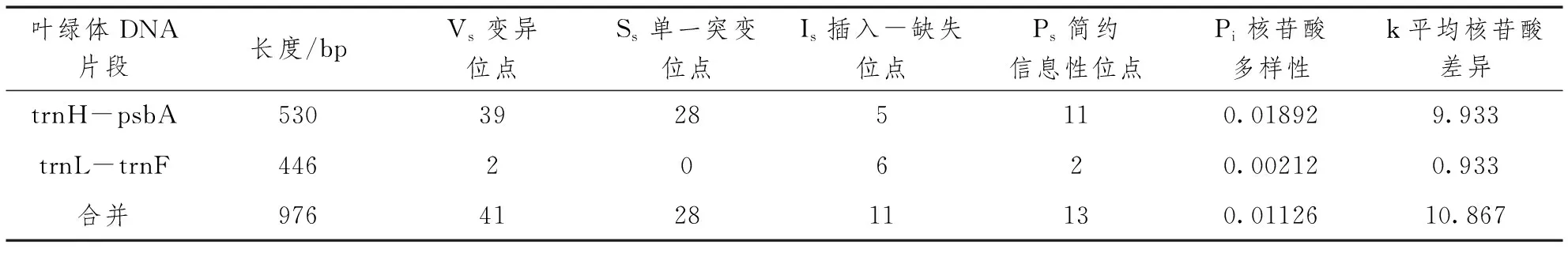
表3 供试老鹰茶植物的2个叶绿体DNA区域的多态性信息
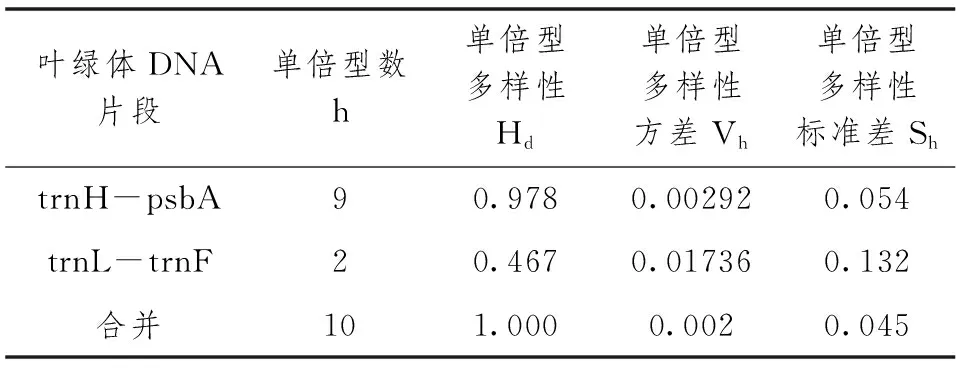
表4 供试老鹰茶植物叶绿体DNA单倍型多样性信息
10份老鹰茶植物的4个叶绿体间区共有多态性变异位点41个,2个区域H-psbA与L-F的变异位点分别为39个和2个,区域H-psbA含有其中95.1%的变异位点,是老鹰茶植物叶绿体DNA间区中多态性较高的区域。H-psbA区域的核苷酸多样性(Pi)为0.01892,平均核苷酸差异(k)为9.933;L-F区域的核苷酸多样性(Pi)为0.00212,平均核苷酸差异(k)为0.933。H-psbA区域的单倍型数目(h)与单倍型多样性(Hd)较高,分别为9个和0.978,但其单倍型多样性方差(Vh)和单倍型多样性标准差(Sh)较低,分别为0.00292和0.054;L-F区域的单倍型数目(h)与单倍型多样性(Hd)较低,分别为2个和0.467,但其单倍型多样性方差(Vh)和单倍型多样性标准差(Sh)较高,分别为0.01736和0.132。
10份老鹰茶植物的2个叶绿体DNA间区组合之后的片段共有10个单倍型,其核苷酸多样性(Pi)、平均核苷酸差异(k)、单倍型多样性(Hd)、单倍型多样性方差(Vh)和标准差(Sh)分别为0.01126、10.867、1.000、0.002、0.045。
2.4 Tajima’s检测
对2个区域进行Tajima’s测验,结果见表5,trnH-psbA区域中的Tajima’s D值是负值,为-1.35554,trnL-trnF区域中的Tajima’s D值是正值,为1.03299,2个区域均在P>0.10水平上不显著。2个叶绿体DNA间区序列组合之后的Tajima’s D值为-1.21568,在P>0.10水平上不显著,说明合并后的片段遵循中性进化模型。

表5 10份老鹰茶植物叶绿体DNA片段的Tajima’s值
2.5 老鹰茶不同类型间遗传关系
基于叶绿体DNA引物trnH-psbA与trnL-trnF扩增出来的序列通过合并后进行聚类分析,采用Neighbour-Joining(NJ)[11]的方法构建系统发育树,结果见图2,黄芽株之间亲缘关系较近,红芽株之间亲缘关系较近,白芽株与红芽株、黄芽株之间亲缘关系较远,但该方法并不能有效区别黄芽株之间的雌雄差异,聚类结果与传统解剖学分类结果大致相同,说明此聚类结果可信。
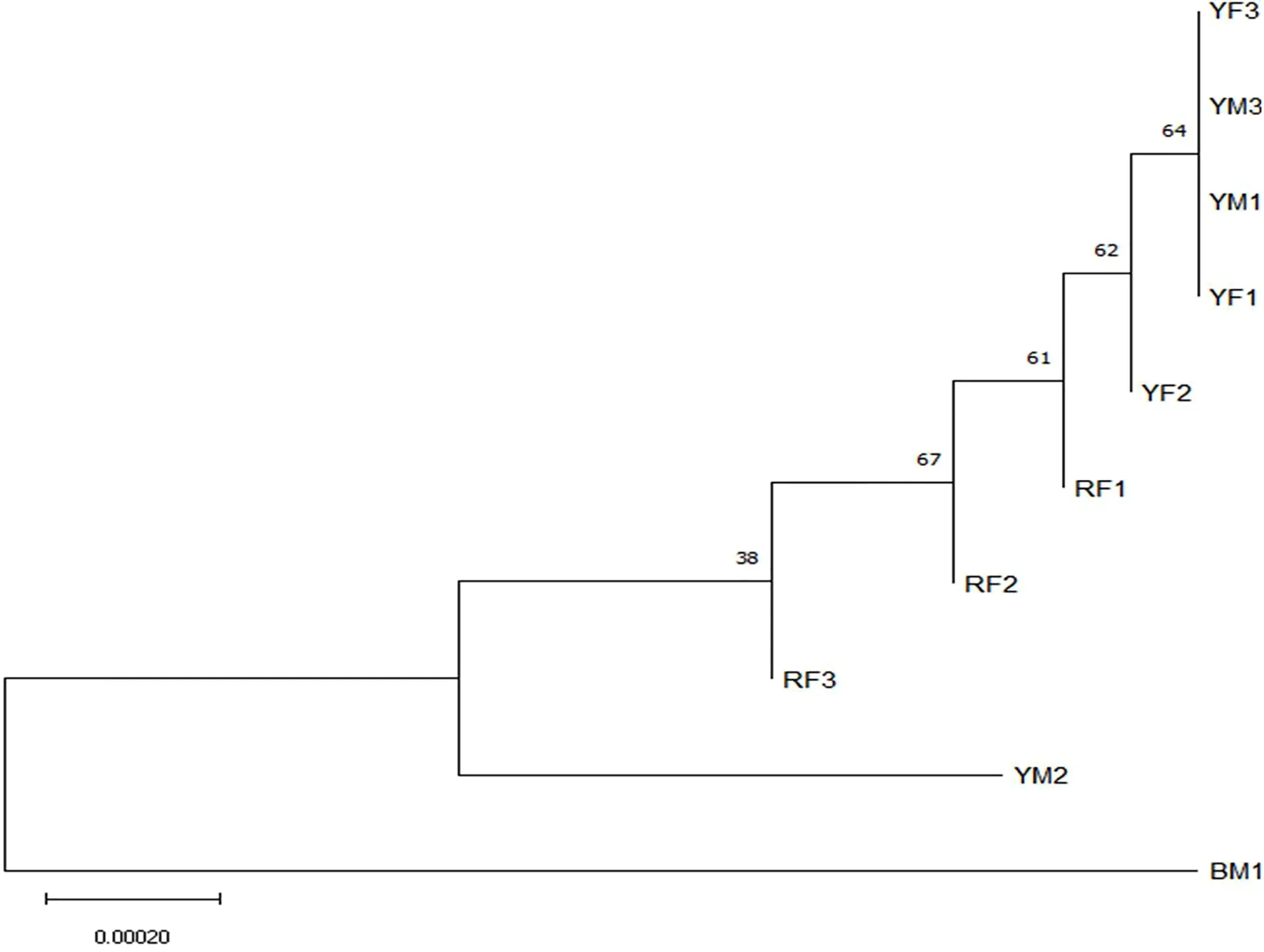
图2 10份老鹰茶植物基于NJ法构建的系统发育树
3 结论与讨论
学者Feng等[6]通过对中国7个省区的23份豹皮樟进行了trnH-psbA、trnL-trnF和trnT-trnL 3个叶绿体标记进行单倍体类型研究,本研究参考其方法,针对贵州省老鹰茶植物进行引物验证,发现trnT-trnL区域测序结果出现重叠峰,因此本研究涉及到的叶绿体DNA区域为trnH-psbA与trnL-trnF。
在贵州省遵义市湄潭县所收集的老鹰茶古树样本,测序后将trnH-psbA与trnL-trnF这2个叶绿体DNA区域合并后发现,有28个单一突变位点、13个简约信息位点和11个插入-缺失位点,共计41个多态性变异位点,2个区域H-psbA与L-F的变异位点分别为39个和2个。区域H-psbA是老鹰茶植物叶绿体DNA间区中多态性较高的区域,变异位点占95.1%,包含28个单一突变位点、11个简约信息位点和5个插入-缺失位点,可见区域H-psbA的突变类型主要表现为单一位点突变;区域L-F仅含2个简约信息位点和6个插入-缺失位点,没有单一突变位点,可见该区域突变类型主要表现为插入-缺失位点。通过计算得到贵州老鹰茶植物的叶绿体DNA核苷酸多样性(Pi)为0.01126,单倍型多样性(Hd)为1.000。在Tajima’s测验中发现,区域H-psbA与L-F在各检验水平上均不显著,说明这2个叶绿体DNA区域在进化上遵循中性模型,在分子水平导致进化的原因不是自然选择,而是突变压力和遗传漂变。
贵州省遵义市湄潭县老鹰茶叶绿体DNA序列与GenBank中的Actinodaphne lancifolia(登录号为NC_045251.1)的朝鲜木姜子种的相似值达100%。将10份老鹰茶植物基于叶绿体DNA引物trnH-psbA与trnL-trnF扩增出来的序列通过合并进行聚类分析,采用Neighbour-Joining(NJ)[11]的方法构建系统发育树,能够有效区分白芽株、红芽株、黄芽株,但不能有效区分黄芽株之间的雌雄差异。在10份老鹰茶植物中,黄芽雄株2号(YM2)的聚类结果较为特殊,距离其他黄芽株都较远,距离白芽雄株更近,由于黄芽株是白芽株与红芽株的后代,属于杂合株,可能黄芽雄株2号(YM2)的父本优势较强而表现的,该部分猜想还需下一步试验来验证。
