白菜NAC基因家族全基因组鉴定及其应答春化反应的表达分析
2022-08-11陈国户庞小可李广王浩吴思文温宏伟尹倩袁凌云侯金锋唐小燕汪承刚
陈国户,庞小可,李广,王浩,3,吴思文,温宏伟,尹倩,袁凌云,侯金锋,唐小燕,汪承刚*
(1.安徽农业大学园艺学院/安徽省园艺作物育种工程实验室,安徽 合肥 230036;2.安徽省皖江蔬菜产业技术研究院,安徽 马鞍山 238200;3.山东省济宁市鱼台县农业技术推广中心,山东 济宁 272300)
转录因子(transcription factor)是一类特异性蛋白质分子,可以与顺式作用元件(cis-element)发生特异性结合,或与其他蛋白相互作用调控目的基因的表达强度,在生命活动中起着重要的作用[1]。转录因子根据不同DNA结合域,可以将其分为NAC(NAM-ATAF1/2-CUC2)、AP2/EREBP(APETALA2/ethylene-responsive element binding proteins)、HB(heparin-binding)、MYB(v-myb avian myeloblastosis viral)、MADS(MCML,AGAMOUS,DEFICIENS,SRF)和SBP(squamosa promoter binding protein)等多个家族[2]。其中,NAC基因家族广泛存在于各植物中,是植物中最大、最重要的转录因子之一,参与各种生物过程的调控[3]。自矮牵牛属中发现第1个NAC转录因子编码基因NAM后,NAC基因家族已在多种植物中被全基因组鉴定,例如拟南芥有117成员[4],烟草有152个[5],水稻有151个[6],棉花有145个[7],玉米有87个[8]。
NAC基因家族名称由NAM(矮牵牛无顶端分生组织)、ATAF1/2(拟南芥转录激活因子1/2)和CUC2(拟南芥杯状子叶)首字母组合而成[9]。NAC转录因子蛋白N端有1个保守结构域,该结构域由5个基序(A、B、C、D和E)组成,长度约160个氨基酸,侧翼含有α-螺旋和β-折叠结构;其中A、C和D基序高度保守,A可能与NAC蛋白形成二聚体有关,C和D基序存在核定位信号并参与DNA的结合;B与E亚结构变异较大,可能与NAC基因的功能多样性密切相关[10]。NAC转录因子蛋白C端结构域差异很大,具有转录激活或抑制活性,参与多种网络的调控[11]。
NAC转录因子是一种多功能蛋白质,在调节植物生长发育、次生代谢及胁迫反应中具有重要的生物学功能,如茎尖分生组织、侧根、胚胎、种子发育[12-14],叶片衰老[15],细胞周期调控[16],以及各种应激反应中激素信号途径[14]等。NAC转录因子还在调控花器官发育方面发挥作用。例如,拟南芥AIF(antherindehiscencefactor)在花期发育过程中异位表达,导致花药不裂,出现不育表型[17];NST1(NACsecondarywallthickeningpromotingfactor1)和NST2也调控花药开裂[18];棉花NAC转录因子GhFSN5在拟南芥中异源表达,导致拟南芥转基因植株产生较小的荚果以及严重的不育性[19];甘蔗ScNAC23基因在拟南芥中异位表达加速了转基因植株抽薹、开花和叶片衰老[20];麻黄CcNAC1基因的过表达加速了植物的生长,促进了早期开花,提高了植物的耐旱性[21]。
白菜(Brassicarapa)是十字花科芸薹属中最重要的蔬菜之一,在全球范围内广泛种植和消费[22]。近年来,研究人员对白菜转录因子家族进行了大量的研究[23-25],其中对白菜NAC基因家族在不同发育阶段、环境胁迫和激素处理下的表达也进行了研究[26]。但NAC转录因子在应答春化反应中的表达分析尚未有报道。本研究通过生物信息学分析方法对白菜NAC转录因子进行全基因组鉴定,并对其理化性质、染色体分布、顺式作用元件、系统进化树及共线性等进行分析,同时基于转录组数据及RT-qPCR分析其在白菜不同组织及应答春化反应的表达模式,旨在为深入探索NAC基因应答春化反应的分子机制奠定基础。
1 材料与方法
1.1 数据来源
拟南芥基因组数据(TAIR10)来源于TAIR数据库;白菜基因组数据(V1.5)来源于BRAD数据库;NAC结构域序列文件(PF02365)从Pfam网站下载。白菜不同组织(根、茎、叶、花、荚等)[27]及春化反应(JWW春化0、25、30、35、45、50 d;XBJ春化0、10、15、25、40、50 d;4004春化0、35 d)[28-29]RNA-Seq原始数据从NCBI GEO与SRA数据库下载。RNA-Seq原始数据经过Trimmomatics及FastQC质控后,利用Hisat2比对至白菜基因组(V1.5)序列,再利用Feature Counts进行有参照转录组定量;基因表达水平采用FPKM值进行计算,差异表达基因采用DESeq2软件包进行计算[25,30]。
1.2 白菜NAC基因家族成员的鉴定
利用HMMER 3.0软件,以PF02365为模型,对白菜基因组(V1.5)进行全基因组搜索;同时利用拟南芥NAC基因家族蛋白序列[4],采用BLASTp技术对白菜基因组进行全基因组同源搜索(top≤5,E<1e-5)。将共同获得的白菜NAC候选序列,在InterProScan(https://www.ebi.ac.uk/interpro/about/interproscan/)、Pfam(http://pfam.xfam.org/)、SMART(http://smart.embl-heidelberg.de/)、SwissProt(https://www.expasy.org/resources/uniprotkb-swiss-prot)以及NR(https://www.ncbi.nlm.nih.gov/)等数据库中逐一鉴定;鉴定后的NAC基因家族再与白菜BRAD数据库中NAC基因家族成员进行比较,对有疑问的基因采用ClustalW及MEGA 7.0软件构建系统发育树进行鉴定,最终确定白菜NAC基因家族成员。
1.3 白菜NAC基因染色体定位及理化性质分析
利用TBtools软件[31]提取白菜NAC基因家族各基因、蛋白序列以及注释等信息,并绘制染色体定位图谱。使用EXPASY在线工具(https://web.expasy.org/compute_pi/),对NAC家族蛋白的相对分子质量与等电点等信息进行分析。
1.4 白菜NAC基因的保守域和系统进化树分析
通过MEME在线工具(http://meme-suite.org/tools/meme),对白菜NAC转录因子蛋白序列的保守基序进行分析,再利用TBtools软件对白菜NAC转录因子基因结构及结构域进行可视化分析。参考宋蒙飞等[32]的方法,使用ClustalW工具将拟南芥与白菜NAC蛋白序列进行比对,并利用MEGA 7.0软件,采用邻接法以及泊松校正与成对删除法(Bootstrap=1 000)构建系统进化树。
1.5 白菜NAC基因的共线性分析
利用TBtools软件对白菜及拟南芥NAC基因家族的蛋白序列进行all-BLASTp-all分析(top≤5,E<1e-5),采用TBtools软件中MCScanX工具提取共线性基因对,再利用Circos工具进行可视化。
1.6 白菜NAC基因家族组织表达及其应答春化反应分析
利用白菜不同组织转录组数据[27],分析白菜NAC基因家族的组织表达特征;利用白菜春化反应材料转录组数据[28-29],分析白菜NAC基因家族应答春化反应关键基因。对NAC基因家族春化反应关键基因,利用荧光定量PCR技术分析其在JWW和XBJ不同春化时间(JWW春化0、25、30、35、45、50 d;XBJ春化0、10、15、25、40、50 d)的相对表达量[28]。
2 结果与分析
2.1 白菜NAC基因家族的鉴定与理化性质分析
通过HMMER与BLASTp搜索,对白菜NAC基因家族进行全基因组鉴定,经过严格筛选,共获得188个NAC基因家族成员,对其序列进行了理化性质分析。白菜NAC基因家族碱基与氨基酸长度、相对分子质量与等电点等变异较大,NAC基因开放阅读框(ORF)长度最长为3 522 bp(Bra012470),最短为375 bp(Bra010895),氨基酸长度为124~1 173,相对分子质量为(14.43~128.20)×103,等电点(pI)为4.43~9.88(图1),表明它们可能在不同的微细胞环境中发挥着不同的作用。对其等电点与相对分子质量之间关系进行分析,发现白菜NAC基因家族蛋白相对分子质量主要集中在(20~60)×103,且酸性蛋白数量远大于碱性蛋白数量(图1)。

图1 白菜NAC基因家族蛋白等电点与相对分子质量分析Fig.1 Putative isoelectric points(pI)and relative molecular weights of NAC gene family proteins in Brassica rapa
2.2 白菜NAC基因家族染色体分布
如图2所示:白菜NAC基因家族188个成员不均等分布在10条染色体上。其中,A04号染色体上分布的NAC基因数量最少,只有5个成员(2.66%);A03号染色体上分布的NAC基因数量最多,有29个成员(15.4%),说明各染色体上的NAC基因数量与染色体大小无关。此外,白菜NAC基因家族中未发现基因串联重复现象。
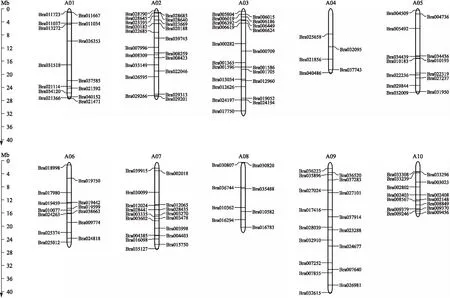
图2 白菜NAC基因家族的染色体定位Fig.2 Chromosomal mapping of NAC family genes in B.rapa
2.3 白菜NAC基因结构与保守域分析
基于白菜NAC基因家族的系统发育关系,对白菜NAC基因家族成员的内含子/外显子分布及保守基序进行分析,发现白菜NAC基因家族成员的基因结构及保守基序差异较小,并且基因结构和保守基序决定了其亲缘关系的远近(图3)。基因结构分析表明,白菜NAC基因家族成员含有0~16个数量不等的内含子;其中,4个成员(Bra004575、Bra004579、Bra033405、Bra036034)不含内含子,1个成员(Bra012470)含有16个内含子,其余大部分成员仅含有2~6个内含子,并且聚集在一起的NAC基因具有相似的结构。保守基序分析表明,白菜NAC基因家族成员基序结构与分布高度相似,N端结构域非常保守,绝大部分成员N端均含有motif 2、motif 3、motif 4、motif 5(NAC基因家族的基本特征),说明这些基序是NAC蛋白功能所必需的。
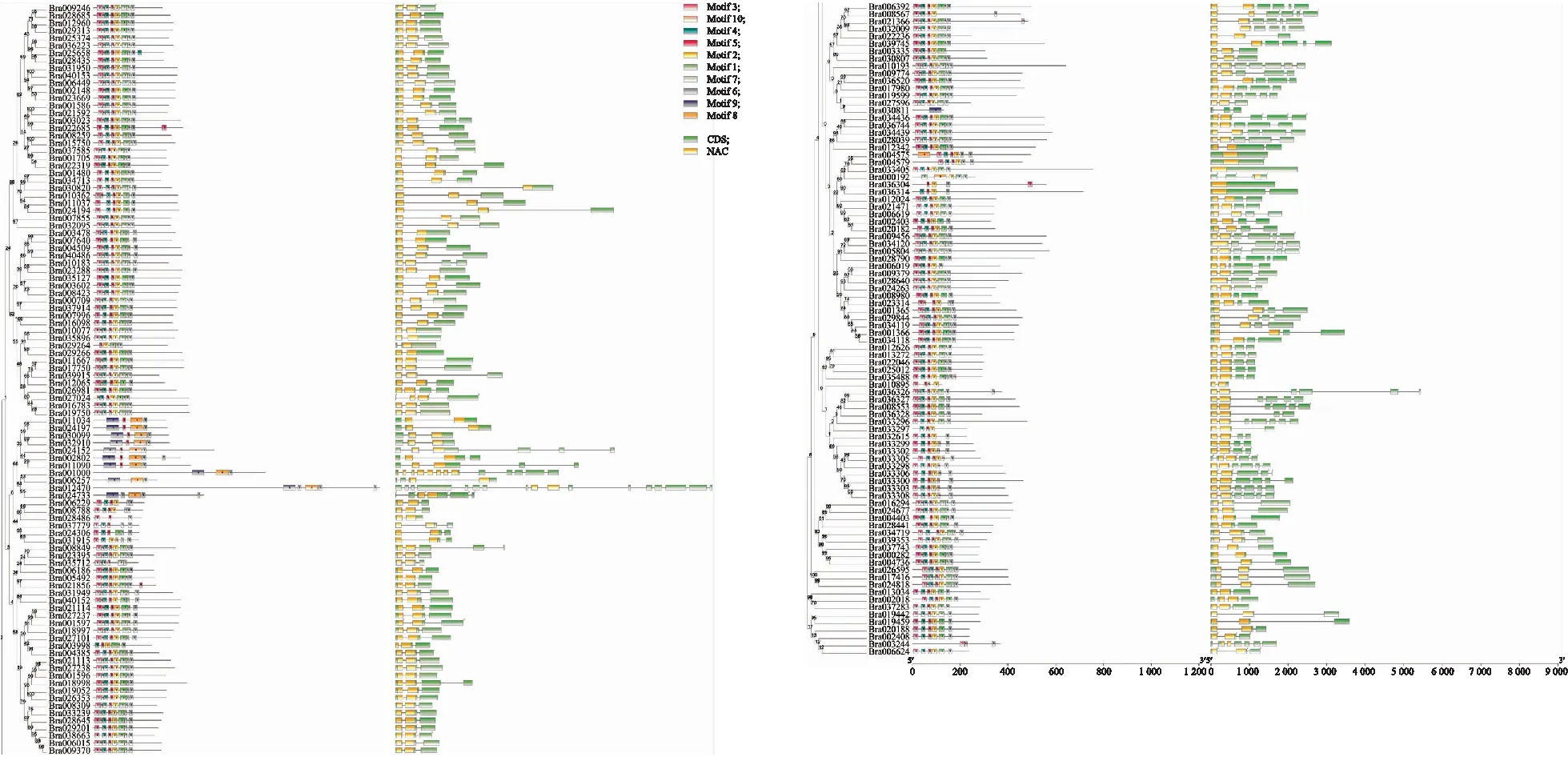
图3 白菜NAC基因家族保守基序及基因结构分析Fig.3 Conserved motif and gene structure analysis of NAC gene family in B.rapa
2.4 白菜NAC基因的系统发育分析
对白菜和拟南芥NAC家族成员蛋白的氨基酸序列进行系统进化树分析,结果(图4)表明,188个BrNAC与AtNAC成员共分成7个亚簇,其中A亚簇成员数量最多,含有66个BrNAC与35个AtNAC成员;B亚簇含有41个BrNAC与18个AtNAC成员;C亚簇含有7个BrNAC与8个AtNAC成员;D亚簇含有 47个BrNAC与20个AtNAC成员;E亚簇含有20个BrNAC与8个AtNAC成员;F亚簇含有5个BrNAC与3个AtNAC成员;G亚簇成员数量最少,仅含有2个BrNAC与2个AtNAC成员。各簇中BrNAC与AtNAC成员数量比值为1∶1~2.5∶1,表明白菜和拟南芥具有较近的亲缘关系,且两者之间存在着一些关系密切的同源NAC基因。
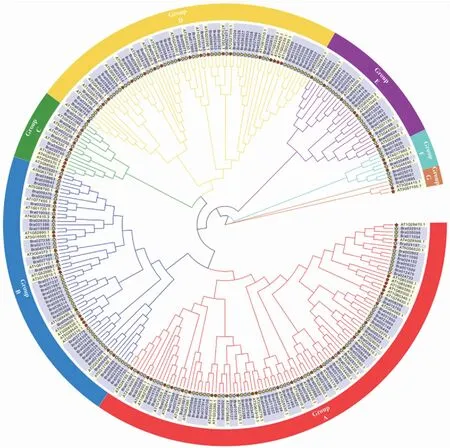
图4 白菜与拟南芥NAC基因家族系统进化树分析Fig.4 Phylogenetic tree of NAC gene family from B.rapa and Arabidopsis thaliana
2.5 白菜NAC家族基因共线性分析
由图5可见:白菜NAC基因家族188个成员中,共有155对旁系同源基因对,其中89对存在着单个基因对应多个基因的情况(1∶1~1∶4),表明白菜NAC家族在发育进化过程中发生了大量的基因复制现象。白菜与拟南芥NAC基因家族之间,同样存在着大量的直系同源基因对,160个BrNAC基因与88个AtNAC基因组成228个共线性基因对(图6),表明这些基因对可能来自同一祖先。
2.6 白菜NAC家族基因表达模式及响应春化反应分析
如图7所示:白菜NAC基因家族188个成员中,大部分成员在白菜不同组织中有转录表达(图7-A);对各组织FPKM>10的BrNAC基因进行分析,发现仅有13个BrNAC成员在各组织中有较高的表达丰度(图7-B),表明这些基因可能在白菜整个生长发育阶段发挥着重要的作用。此外,在高转录丰度成员中,有21个成员在花与荚中特异表达,有6个成员在根中特异表达,表明这些特异表达BrNAC基因具有较强的组织表达特异性,可能对促进特定组织的发育起着重要的作用。

图5 白菜NAC基因对复制事件分析Fig.5 Analysis of duplicated NAC gene pairs in B.rapa
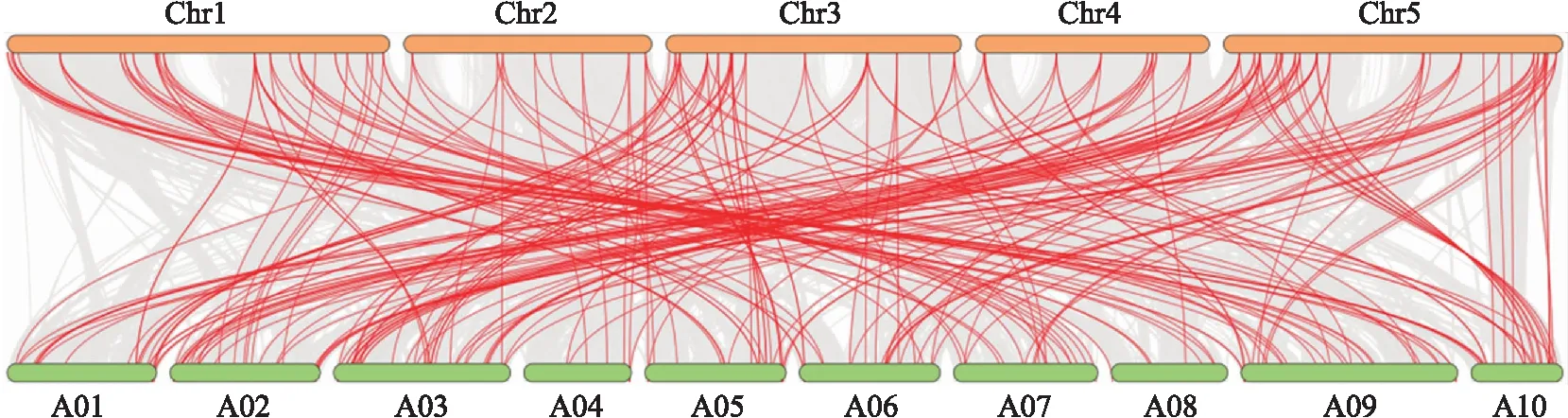
图6 拟南芥(Chr1—Chr5)与白菜(A01—A10)NAC基因家族的共线性分析Fig.6 Synteny analysis of NAC gene family between A.thaliana(Chr1-Chr5)and B.rapa(A01-A10)

图7 白菜NAC家族基因的组织表达模式分析[log2(FPKM)](A)及各组织中 高表达基因(FPKM>10)韦恩图分析(B)Fig.7 Expression analysis of NAC genes in different tissues based on RNA-Seq[log2(FPKM)](A) and Venn analysis of high-expression genes(FPKM>10)in different tissues(B)
利用白菜春化反应转录组数据分析BrNAC差异表达基因,结果(图8)表明,在白菜JWW、XBJ与4004材料春化转录组数据中,差异表达BrNAC基因共有64个,其中共有28个基因至少在2个材料中差异表达,说明这些BrNAC基因可能与春化反应相关。在28个应答春化反应差异表达基因中,6个BrNAC基因下调表达,22个BrNAC基因上调表达。

图8 白菜NAC基因家族应答春化反应分析Fig.8 Analysis of NAC gene family responsed to vernalization in B.rapa A. 白菜NAC基因应答春化反应DEG表达分析;B. 白菜材料 JWW、XBJ和4004中NAC基因应答春化反应DEG韦恩图分析。A. Expression analysis of BrNAC genes responsed to vernalization;B. Venn analysis of BrNAC genes responsed to vernalization among Chinese cabbage materials JWW,XBJ and 4004.
对JWW与XBJ材料中差异表达基因,分别进行表达趋势分析,结果如图9所示。JWW与XBJ这2个材料有相同的显著富集的趋势(P<0.05)。白菜NAC基因家族成员表达丰度,随春化时间的延长,2个材料共有16个基因逐渐上调表达(趋势7),分别有14个基因在春化25 d(JWW)及22个基因在春化10 d(XBJ)时丰度达到最大(趋势6)。此外,在显著富集趋势中,JWW与XBJ两个材料共同有BrNAC基因 12个,其中5个BrNAC基因与春化密切相关,表明这些基因为白菜NAC家族应答春化关键基因。

图9 白菜JWW和XBJ NAC基因应答春化反应DEG表达趋势(A、B)及韦恩图(C)分析Fig.9 DEG expression profiles(A,B)and Venn analysis(C)in Chinese cabbage JWW and XBJ
利用荧光定量PCR技术,进一步分析白菜NAC基因家族应答春化反应关键基因及春化途径基因BrFLC1的表达水平,结果如图10所示。Bra004385、Bra004736、Bra031950、Bra033296、Bra036327基因经过低温春化后,其在JWW和XBJ中的相对表达量均显著上升,其中4个BrNAC基因(Bra004385、Bra004736、Bra031950、Bra036327)的表达量在春化过程中维持较高水平;而Bra033296的表达量,JWW在春化30 d时达到最高,随后逐渐降低,但仍高于春化初始时期的表达量,XBJ在春化10 d时达到最高,随后随着春化时间表达量先降低后升高,再降低,最后再升高;而春化途径BrFLC1基因随春化时间延迟,其表达水平急剧下降后,缓慢上升然后再下降。
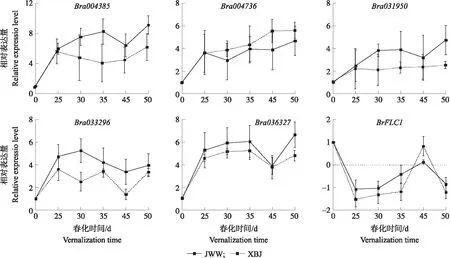
图10 白菜NAC基因应答春化反应关键基因及FLC基因的表达分析Fig.10 Expression analysis of the key NAC and FLC genes related to vernalization in B.rapa
3 讨论
植物特异性NAC转录因子(TF)是植物中最大的转录调节因子家族之一,在植物的生长发育调控和逆境反应中发挥着重要作用[3]。目前,多个物种的NAC基因家族已被鉴定出来[4-8],但对白菜NAC基因家族应答春化反应知之甚少。本研究中,利用同源比对及HMMER搜索,结合多重数据库严格筛选,共鉴定出188个BrNAC转录因子成员,是拟南芥NAC基因家族数量的1.5倍,但显著少于Liu等[26]的报道,其最主要原因为他们对白菜全基因组进行HMMER筛选时采用了较低的筛选参数(E<1e-3),并且未进行多个数据库的筛选鉴定。
在多基因家族进化过程中,基因结构普遍存在多样性,有利于基因进化选择新功能,以适应环境变化[33]。本研究对白菜NAC基因家族成员基本特征进行分析,其家族全部成员蛋白序列N端均含有NAM结构域、基因结构多样,但同一聚类亚组中的NAC成员具有相似的功能基序、外显子/内含子结构和位置,表明这些成员关系密切,可能具有相似的功能,这与Liu等[26]的研究一致。此外,同一亚组中的成员之间也存在序列大小、外显子的相对位置和数量等差异,这可能是白菜NAC基因家族在进化过程中发生了基因片段的剪接和插入所致,从而获得了新的基因功能[34]。
白菜在进化过程中经历过2次全基因组复制及1次全基因组三倍化事件[35]。本研究对白菜与拟南芥NAC成员进行系统进化树分析,7个亚簇中BrNAC与AtNAC成员数量比值为1∶1~2.5∶1,表明白菜与拟南芥之间在进化中存在着密切相关的同源NAC基因。白菜与拟南芥NAC基因家族成员之间存在大量的直系同源基因对,表明白菜NAC基因家族大部分成员与拟南芥NAC基因成员可能来自同一祖先,且白菜NAC基因家族的扩张主要来源于基因复制事件[33]。
大部分BrNAC基因具有组织表达特异性,仅少数BrNAC转录因子在白菜各组织中有较高的表达丰度,表明它们可能在白菜的生长发育中起重要作用。本研究中进一步利用春化转录组数据[28-29]分析白菜NAC基因家族应答春化反应关键基因,结果共有6个BrNAC成员在JWW、XBJ及4004材料春化前、后均显著差异表达,表明这些基因与春化反应密切相关。对JWW与XBJ材料中差异显著的BrNAC基因进行表达趋势分析,显示显著表达趋势中共有40个BrNAC基因;其中5个BrNAC成员为上述显著差异表达基因一致,进一步表明这些BrNAC成员与春化反应密切相关。对这5个BrNAC成员在不同春化时期的表达水平进行分析,显示其相对表达量均显著上升,并且在春化50 d时仍维持着较高的表达量;此外,上述 5个白菜NAC基因的表达趋势与春化关键基因FLC的表达趋势相反,特别是与Bra036327基因完全相反,表明该NAC基因为白菜春化应答基因,在白菜春化过程中可能具有重要的作用。
